Abstract
The symbiotic relationship between nitrogen-fixing cyanobacteria and plants offers a promising avenue for sustainable agricultural practices and environmental remediation. This review paper explores the molecular interactions between nitrogen-fixing cyanobacteria and nanoparticles, shedding light on their potential synergies in agricultural nanotechnology. Delving into the evolutionary history and specialized adaptations of cyanobacteria, this paper highlights their pivotal role in fixing atmospheric nitrogen, which is crucial for ecosystem productivity. The review discusses the unique characteristics of metal nanoparticles and their emerging applications in agriculture, including improved nutrient delivery, stress tolerance, and disease resistance. It delves into the complex mechanisms of nanoparticle entry into plant cells, intracellular transport, and localization, uncovering the impact on root-shoot translocation and systemic distribution. Furthermore, the paper elucidates cellular responses to nanoparticle exposure, emphasizing oxidative stress, signaling pathways, and enhanced nutrient uptake. The potential of metal nanoparticles as carriers of essential nutrients and their implications for nutrient-use efficiency and crop yield are also explored. Insights into the modulation of plant stress responses, disease resistance, and phytoremediation strategies demonstrate the multifaceted benefits of nanoparticles in agriculture. Current trends, prospects, and challenges in agricultural nanotechnology are discussed, underscoring the need for responsible and safe nanoparticle utilization. By harnessing the power of nitrogen-fixing cyanobacteria and leveraging the unique attributes of nanoparticles, this review paves the way for innovative, sustainable, and efficient agricultural practices.
1. Introduction
Molecular nitrogen (N2) makes up most of the atmosphere on earth and accounts for around 78% of the total gas volume. Despite its abundance, most living things cannot use gaseous nitrogen directly [1]. To overcome this limitation, certain microorganisms have evolved the ability to perform biological nitrogen fixation (BNF), converting atmospheric nitrogen into bioavailable forms such as ammonia and other nitrogen compounds [2]. To maintain the finite supply of biologically accessible nitrogen, BNF is a crucial ecological mechanism [3,4]. Primary productivity in a variety of ecosystems, including terrestrial and aquatic environments, as well as symbiotic relationships with plants, depend on this process [5]. Among the numerous BNF-capable species, nitrogen-fixing cyanobacteria play a crucial role in delivering fixed nitrogen into the biosphere [6].
The phylum of prokaryotic microbes known as cyanobacteria, or blue-green algae, has a spectacular evolutionary history that dates back around 2.7 billion years [7,8,9]. Earth’s atmosphere was shaped by these ancient bacteria because they were among the first to use photosynthesis, in which oxygen is released as a byproduct of metabolic activity [10,11,12]. Cyanobacteria occur in many different habitats and exhibit a wide range of morphologies, including unicellular, filamentous, and even colonial forms [13,14,15]. Nitrogen-fixing cyanobacteria carry out solar-powered BNF independently as free-living organisms, in contrast to higher plants that depend on symbiotic relationships with rhizobia for nitrogen fixation [16]. They have specialized cell structures termed heterocysts, which are oxygen-limited environments where nitrogen fixation occurs [17,18]. The ability of cyanobacteria to sequester oxygen-sensitive nitrogenase enzymes within heterocysts allows for effective nitrogen fixation while guarding against deactivation of the enzymes by oxygen [6,19,20]. Due to their exceptional traits and ecological importance, nitrogen-fixing cyanobacteria have drawn considerable attention for their potential applications in sustainable agriculture and environmental remediation [21]. Harnessing the nitrogen-fixing abilities of cyanobacteria could lead to reduced dependence on synthetic fertilizers, mitigating environmental pollution from excessive fertilizer usage [22]. Additionally, their potential to establish beneficial symbiotic associations with crop plants holds promise for enhancing crop yield and overall agricultural productivity [23]. Chemical fertilizers have traditionally been widely used to supplement nitrogen requirements in agriculture, resulting in increasing agricultural yields and sustaining the expanding human population [24]. Eutrophication, greenhouse gas emissions, and water pollution are some of the environmental issues that have been brought up by the overuse of synthetic fertilizers. In this situation, it is crucial to adopt sustainable methods that preserve agricultural production while reducing dependency on chemical fertilizers [25].
Agriculture is one of the many scientific disciplines particularly interested in the emerging science of nanotechnology. A particle is a tiny entity that functions as a unit in nanoscience or nanotechnology in terms of its properties and transportation. Therefore, one of the factors utilized to categorize such particles is the particle diameter. According to a convention, coarse particles are defined between 10,000 and 2500 nm in size, fine particles as those between 2500 and 100 nm in size, and nanoparticles (NPs) between 1 and 100 nm in size, whether they are dispersed in gaseous, liquid, or solid media [26] differ from their bulk counterparts in terms of specific physicochemical characteristics. These properties have allowed them to be used in a wide variety of farming practices, bringing potential benefits but also difficulties that require careful consideration.
Particularly in recent years, metal nanoparticles have shown promise for modernizing farming methods and advancing sustainable crop production. Silver (Ag), zinc oxide (ZnO), copper oxide (CuO), and iron oxide (Fe2O3) are metal nanoparticles with exceptional antibacterial, antioxidant, and catalytic characteristics. They can also be synthesized into precise shapes and sizes, which increases their adaptability and potential agricultural uses [27]. Scaridia galli, a common intestinal parasite in poultry, could be fought off using secondary metabolites from medicinal plants. The study examined the utilization of several secondary metabolites, such as alkaloids, flavonoids, tannins, saponins, and terpenes, found in diverse medicinal plants, as natural substitutes for synthetic medications for treating parasitic diseases in chickens [28]. To assess the potential of nanoplastics for environmental harm, particularly the toxicity of polystyrene nanoparticles to microalgae. The studies were carried out to determine the impact of various polystyrene nanoparticle concentrations on the development of a specific microalgae species. To analyze the toxicity of the nanoparticles, they evaluated variables like growth rate, biomass, chlorophyll content, and cell viability. The study’s findings showed that the growth and viability of microalgae were negatively impacted by exposure to polystyrene nanoparticles. Higher concentrations of nanoparticles resulted in slower growth rates, lower biomass production, less chlorophyll, and more cell death. The results imply that aquatic ecosystems, particularly microalgae populations, may be threatened by nanoplastics like polystyrene nanoparticles [29,30]. Various types of carbon nanoparticles are discussed, including fullerenes, graphene, carbon nanotubes, carbon nanofibers, and black carbon. It divulges details regarding their compositions, dimensions, and characteristics. The chapter also discusses many approaches, including chemical and physical ones, for synthesizing nanoparticles. Evaporation-condensation, spark discharge, and thermal breakdown are examples of physical processes; metallic precursors, reducing agents, and stabilizing agents are examples of chemical approaches. The section shows the benefits and difficulties of each synthesis technique [31,32,33].
The primary goals of using metal nanoparticles in agriculture are to increase agricultural yield as well as plant growth and nutrient uptake. These nanoparticles can have molecular interactions with plant systems, altering cellular functions and favorably affecting plant metabolism [34,35,36,37,38,39]. The targeted distribution of nutrients, hormones, and agrochemicals to plants is made possible by the role of metal nanoparticles as nanocarriers, which optimizes resource use and minimizes environmental effects [40,41]. Metal nanoparticles have enormous potential for use in agriculture; however, their possible detrimental effects on the environment and human health also raise questions. It is possible that the distinctive physicochemical characteristics of nanoparticles, which make them useful for a variety of applications, may also be to blame for their heightened reactivity and potential toxicity. To ensure the safe and sustainable application of metal nanoparticles, it is crucial to understand their fate and behaviors in agricultural ecosystems [42]. Numerous industries have been transformed by nanotechnology, which has provided ground-breaking solutions in fields like electronics, health, and environmental remediation [43]. Nanoparticles (NPs), one of the most promising developments, have drawn significant interest because of their distinctive physicochemical characteristics and possible uses [44]. These characteristics make NPs exceedingly desirable for a variety of applications, including improved imaging [45], improved catalytic activity, and targeted drug administration [46]. Although there is no denying the potential advantages of NPs, widespread acceptance of them has been significantly hampered by concerns about their safety and environmental impact [47]. Potential dangers may also be exacerbated by the distinct physicochemical characteristics of NPs that make them desirable for applications [48]. For example, due to their small size, NPs can easily overcome biological barriers and pose a risk to human health when they come into contact with living beings [47]. Additionally, the high surface area-to-volume ratio of NPs may increase reactivity and increase the risk of toxicity [49].
A detailed comprehension of NPs’ characteristics and behaviors in various situations is necessary due to the complex interaction between their advantages and hazards [50]. Effective risk assessment requires a thorough analysis of the toxicity profiles, fate, and transit of NPs in multiple environmental compartments [51]. To develop strategies to minimize potential hazards while maximizing the benefits of NPs, a multidisciplinary approach involving regulatory authorities, toxicologists, environmental scientists, and nanotechnology specialists is required [52]. Due to the growing global population and increasing demand for food, there is a need to increase agricultural production while ensuring sustainable agricultural practices [53]. In this context, metal nanoparticles (MNPs), which are at the cutting edge of research in the field of improving plant growth, have proven to be a promising way to solve these problems [54]. Due to their distinct physicochemical characteristics and high surface area-to-volume ratio, metal nanoparticles have a wide range of potential applications in agriculture [55,56]. The size, shape, and surface functionalization of MNPs can be changed through the controlled synthesis process, which affects how they interact with plants and their environment [57]. Additionally, MNPs have natural antibacterial characteristics that can aid in reducing plant diseases caused by various pathogens [58]. MNPs have therefore demonstrated potential as a greener substitute for traditional agrochemicals, reducing environmental impact and encouraging sustainable agriculture [1,59].
MNPs have great potential for use in agriculture, but it is important to assess their possible effects on the environment and their long-term consequences for soil health. To understand the benefits and potential hazards of MNP use in agriculture, a thorough evaluation of the available studies on their effects on plant growth enhancement and sustainability is required [60].
2. Nanoparticle Uptake and Distribution in Plants
The use of nanoparticles (NPs) in agricultural practices has opened up new opportunities to improve crop productivity and sustainability. Due to their unique physicochemical characteristics and high surface area-to-volume ratio, NPs are endowed with novel capabilities beyond conventional agricultural methods [61]. For this reason, NPs have been designed and manufactured to tackle certain agricultural issues such as nutrient deficit, pest control, and soil degradation. However, a thorough comprehension of NPs’ interactions in plants at the molecular, cellular, and physiological levels is necessary for their use in agriculture. The study of NP absorption, transport, and distribution within different plant tissues is crucial to this understanding [62].
A variety of factors influence the complicated process by which plants absorb NPs. The size, shape, surface charge, and composition of NPs, as well as how they interact with plant surfaces and cellular membranes, all depend on their physicochemical properties. These factors together affect how much and how quickly NPs are taken up by cells, with smaller NPs and those with positive charges frequently exhibiting enhanced cellular internalization [63].
The variance in NP uptake efficiency is further influenced by changes in plant species and genotypes, highlighting the complex interplay between plant-specific features and NP properties [64]. By affecting root architecture and physiology, environmental factors such as soil pH, nutrient availability, and organic matter content can significantly regulate NP uptake. NPs in soil can produce toxic effects at the levels of genes, cells, individual organisms, and communities [65]. As shown in Figure 1, NPs can enter cells through active transport and phagocytosis to induce oxidative stress, affect enzyme activity, damage DNA/RNA, and, in other ways, produce cell toxicity. NPs that do not enter cells can cause membrane damage through physical contact, and metal ions released by metal NPs enter cells mainly through passive diffusion to produce toxic effects [66]. It is believed that ROS production is the main mechanism leading to cell damage. Nanoparticles can stimulate the ground-state oxygen in organisms to produce a large number of ROS, such as hydroxyl radical (·OH), singlet oxygen (1O2), (O2−1), hydrogen peroxide (H2O2), etc. [67]. ·OH has a strong oxidation capacity with an oxidation potential of 2.8 V. It can irreversibly destroy many biological macromolecules, including sugars, fats, and proteins [68]. 1O2 can oxidize and damage a variety of biological components and destroy organisms. O2−1 is unstable and can be converted into •OH and 1O2 in biological systems [69]. It can cause an imbalance of the REDOX state in the cell, resulting in oxidative damage to the organism, such as cell inflammation and death [70]. A comprehensive understanding of NP absorption mechanisms that goes beyond passive diffusion and includes endocytic pathways is required due to the complicated interactions between these components [71].
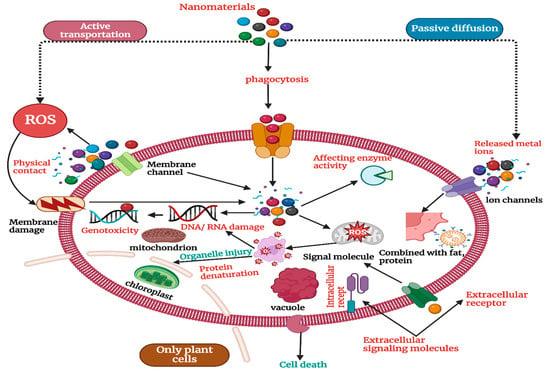
Figure 1.
Mechanism of the toxic effects of nanomaterials on soil organisms.
After internalization, NPs move through the complex network of plant tissues in simplified and apoplectic pathways. Movement through plasmodesmata, which are nanoscale channels connecting adjacent plant cells, is a component of symplastic transport [72]. As an alternative, NPs can move through the extracellular matrix, which consists of cell walls and intercellular voids, to move via the apoplastic space. The cell wall features, the specific developmental stage of the plant, and the physicochemical properties of NPs all influence the selection of the transport route [73]. Rapid NP movement inside the plant is facilitated by symplastic transport mediated by endocytic vesicles and actin-based movement; however, apoplastic transport may be hampered by physical barriers like the Casparian strip in the root endodermis [74].
The effectiveness of NP uptake is significantly influenced by the root architecture of plants. In particular, root tips, lateral roots, and hairs play a key role in mediating NP interactions with the rhizosphere [75].
The complex variety of organic chemicals found in root exudates affects NP behavior at the soil-root interface and can either increase or decrease NP uptake [76]. By developing symbiotic interactions that improve nutrient and water intake, mycorrhizal linkages further modify NP uptake. According to the available reports, the bryophyte-cyanobacteria symbionts can be divided into three classes according to the bryophyte taxa. Namely, tomentosum-cyanobacteria symbiosis, liverwort-cyanobacteria symbiosis, and bryophyte-cyanobacteria symbiosis. So far, it is known that 41 moss species from 35 genera are capable of symbiotic nitrogen fixation with cyanobacteria (Table 1), which represents only a small part of the moss species known worldwide. Mycorrhizal networks can make it easier for NPs to migrate from hyphal networks to plant roots, potentially affecting how they are later transported through the plant [77].
2.1. Mechanisms of Nanoparticle Entry into Plant Cells
In recent years, a lot of research has been done on the relationship between nanoparticles and plant cells [78]. Nanoparticles are attractive candidates for use in agriculture and environmental remediation because they have distinctive physicochemical features that allow them to interact with biological systems in unexpected ways [79]. Their capacity to infiltrate plant cells is one essential factor that controls the effective application of NPs in plant systems. Optimizing the transport and targeting of nanoparticles in plants requires an understanding of the mechanisms underlying their entrance [80].
To assimilate nanoparticles, plant cells have developed different pathways, each involving unique molecular and cellular processes. One of the possible ways in which plant cells can internalize NPs is through endocytosis, a well-studied process in mammalian cells [81]. One or more endocytic mechanisms, including clathrin-mediated endocytosis, caveolin-dependent endocytosis, and clathrin-independent endocytosis, have been proposed to be involved in nanoparticle uptake [82]. Other routes, such as direct passage of nanoparticles through the plasma membrane, have also been proposed [83]. The physical and chemical characteristics of nanoparticles are crucial in defining how they interact with plant cells [84]. The methods of nanoparticle entrance are greatly influenced by surface charge, size, shape, and surface functionalization [85]. For instance, it has been demonstrated that positively charged nanoparticles exhibit greater cellular internalization in comparison to their negatively or neutrally charged counterparts [86]. The diffusion of nanoparticles through cell walls and membranes is influenced by their size, with smaller nanoparticles potentially penetrating more deeply [87].
When nanoparticles enter plant cells, some signaling pathways may be activated [88]. The initial detection of nanoparticles that trigger subsequent signaling processes has been linked to recognition receptors on the cell surface, such as receptor-like kinases [89]. The complex web of interactions between nanoparticles and plant cells is highlighted by the activation of intracellular kinases and secondary messengers in response to nanoparticle uptake [90].
2.2. Cyanobacterial Intercellular Transportation
Cyanobacteria play a crucial role in the world’s biogeochemical cycles due to their capacity to harness light energy and transform carbon dioxide into organic molecules [91]. The complex mechanisms governing the physiology and relationships of cyanobacteria have been revealed thanks to the development of molecular and genomic technologies. However, the biological processes that control the intercellular communication and movement of cyanobacteria are still poorly understood [92].
Cyanobacteria have developed complex signal transduction and resource-sharing mechanisms to ensure coordinated responses to environmental signals. Small molecules, including nutrients and signaling chemicals, diffuse through channels known as septal junctions, one prominent method of intercellular transit. These septal connections, which are comparable to gap junctions in eukaryotic cells, enable direct cytoplasmic connections between neighboring cyanobacteria cells [93]. The function of outer membrane vesicles (OMVs) in facilitating the long-distance transit of bioactive chemicals and genetic material has also been clarified by recent investigations. OMVs, membrane-bound structures released from the cell surface, act as transporters for intracellular communication and may be crucial to the dynamics of the cyanobacterial community [94].
Cyanobacterial populations’ fitness and behaviors are significantly impacted by the interaction between intercellular transportation and cellular coordination [95]. Optimizing resource use and growth requires coordinated responses to environmental changes, such as those in nutrient availability and light intensity [96]. Intercellular movement makes it possible to share necessary metabolites such as fixed carbon and nitrogenous substances, which encourages population division [97]. Additionally, the transfer of genetic material via OMVs may promote horizontal gene transfer and genetic diversity, which may affect how well cyanobacteria can adapt to their environment. To fully understand the intricate dynamics of cyanobacterial communities in natural and manmade systems, it is imperative to comprehend these implications [98].

Table 1.
Types and symbiotic forms of cyanobacteria symbionts.
Table 1.
Types and symbiotic forms of cyanobacteria symbionts.
| Type | Moss Species and Relationship with Cyanobacteria | Cyanobacteria | Reference |
|---|---|---|---|
| Hornwort-cyanobacteria symbiont | Anthoceros sp. Dendroceros sp. Notothylas sp. Phaeoceros sp. | Nostoc sp. | [99,100,101,102,103] |
| Liverwort-cyanobacteria symbiont | Anastrophyllum involutifolium Blasia sp. Cavicularia sp Chiloscyphusleptanthus Marchantia sp. Porella sp. | Nostoc sp. Calothrix sp. Stigonema sp. Chlorogloeopsis sp. | [99,100,101,102,104] |
| Moss-cyanobacteria symbiont | Acroschisma wilsonii Andreaea alpine Andreaea laxifolia Aulacomnium palustre Blepharidophyllum densifolium Bryum sp. Calliergon richardsonii Ceratodon purpureus Clasmatocolea humilis Cryptochila grandiflora Dendroligotrichum squamosum Dicranoloma chilense Ditrichum cylindricarpum Drepanocladus sp Drepanocladus exannulatus Dupontia sp. Grimmia sp. Heteroscyphus magellanicus Hylocomium splendens Paludella squarrosa Pleurozium schreberi Ptilium sp. Racomitrium Subcrispipilum Racomitriumlaevigatum Racomitrium lanuginosum Racomitrium didymium Sanionia uncinate Sphagnum lindebergii Sphagnum riparium Tomentypnum nitens Weisia controversa | Nostoc sp. Nostoc muscorum Calothrix sp Stigonema sp Scytonema sp. Anobena sp. Oscillatoria sp. Lyngbya sp. | [99,100,101,102,105,106,107,108,109] |
There are numerous biotechnological and environmental applications that could benefit from the knowledge learned from investigating cyanobacterial intercellular transfer. The effectiveness of bio-production processes such as biofuel synthesis and bioremediation could be improved by cyanobacterial consortia designed to exploit intercellular communication routes [110]. Additionally, using cyanobacteria’s inherent intercellular transportation systems should make it easier to transfer bioactive substances under controlled conditions for use in environmental engineering applications. Researchers can advance the development of sustainable technologies with broad-reaching benefits by taking advantage of the special characteristics of cyanobacterial communication networks. To fully understand the intricate dynamics of cyanobacterial communities in natural and manmade systems, it is imperative to comprehend these implications [111].
Metal nanoparticles (MNPs) are being used more often in a variety of industrial applications, which has resulted in a significant amount of them being released into the environment, raising worries about potential negative impacts on ecosystems and human health [112]. Due to their ability to absorb and detoxify heavy metals, plant-cyanobacteria systems for bioremediation have become more popular among the numerous methods to reduce the environmental effects of MNPs [113]. The kinetics of absorption, transport, and localization of nanoparticles can be studied using the interactions between MNPs and plant-cyanobacteria systems [113].
2.3. Root-Shoot Translocation
Water, minerals, hormones, and other vital substances travel in both directions through the root-shoot axis. Numerous physiological factors, including the availability of nutrients, hormone signaling, and climatic conditions, influence this transport, which takes place via complex systems involving vascular tissues such as xylem and phloem. According to recent studies, the selective transfer of chemicals between roots and shoots is governed by transport proteins, ion channels, and molecular gradients [114].
2.4. Systemic Distribution and Long-Distance Signaling
Plants communicate in a systemic manner that extends beyond the root shoot axis and involves the transmission of chemicals and information over long distances. The coordination of physiological responses, including defense systems, photosynthesis, and growth regulation, is fundamentally influenced by this process [115]. With its ability to support mass flow, the phloem is a crucial channel for systemic distribution, enabling the spread of pathogens, proteins, RNAs, and nutrients throughout plants. Auxin, cytokine, and abscise acid-based hormonal signaling, in particular, play a role in the coordination of systemic reactions [116].
2.5. Cyanobacterial Symbiosis
Because of their famed photosynthetic powers, cyanobacteria frequently create symbiotic partnerships with plants, aiding in nutrient uptake and stress tolerance. Complex molecular interactions are required for the development of cyanobacterial colonies in plant tissues, and specialized structures such as heterocysts and symbiotic interfaces facilitate nutrient exchange. Abiotic stress resistance, nitrogen fixation, and plant growth are all impacted by this mutually beneficial connection [117].
3. Molecular Insights into Nanoparticle Toxicity
Beginning with cellular uptake, which is controlled by NP size, shape, surface charge, and composition, interactions between NPs and biological systems begin. These physicochemical characteristics have an important role in determining cellular internalization mechanisms, including endocytosis and direct penetration, according to recent studies. Electrostatic interactions may be important because they show that positively charged silver nanoparticles have increased cellular absorption compared with their negatively charged counterparts [118]. Numerous investigations have documented the genome toxicity caused by NP, which includes mutations, chromosomal abnormalities, and DNA strand breaks. These consequences may result from ROS-related secondary effects or from physical damage caused by direct interactions between NPs and DNA. It was revealed that the production of ROS and subsequent activation of DNA repair pathways by gold nanoparticles could cause DNA damage [119]. The key methods for toxicological appraisal of nanoparticles, as described in the search results, include a combination of in vitro and in vivo approaches. In vitro studies using cell lines are crucial for assessing the acute damaging effects of nanoparticles in specific cellular environments [120,121]. Genotoxicity testing is also used to evaluate the potential of nanoparticles to cause genetic damage such as mutations or chromosomal aberrations. Calculating the No Observed Adverse Effect Level (NOAEL) is an important step in nanoparticle toxicity testing, which helps determine the safe dose levels for further studies [122,123]. However, extrapolation of cell-based data into in vivo scenarios is problematic due to the extreme conditions in vitro, so in vivo testing remains crucial for assessing biodistribution, bioaccumulation, and systemic toxicity [124]. To fully investigate the toxicity and potential risk of a particular nanomaterial, a combination of multiple in vitro tests is required. Table 2 lists the toxicity of several typical nanoparticles that have been evaluated using different biological assessment models. The toxicity of nanoparticles to organisms mainly includes the following four aspects: (1) nanoparticles with nanoscale are easy to cross the cell membrane into the cell, easy to damage the cell membrane and interfere with the physiological activity in the cell; (2) Nanoparticles have high activity, and the generated reactive oxygen species (ROS) on the one hand is easy to damage the cell membrane, destroy the permeability of the cell, hinder the exchange of materials between the cell and the outside world, and easily cause protein denaturation; On the other hand, ROS produced by nanoparticles can stimulate the oxidative stress pathway in cells, resulting in cell damage [2]; (3) Nano-oxygen is applied to some nanoparticles that can dissolve metal ions (such as programmed copper, cadmium selenide, etc.), and the dissolved metal ions also enhance the toxicity of nanoparticles in terms of nanoscale. Taking the study on the toxicity of particles to particles in Figure 2 as an example, the toxicity of Escherichia coli nanoparticles can be divided into the following aspects: First, the nanoparticles have a large specific surface area, which can fully interact with the cell membrane of E.coli and destroy the permeability of cells, thus causing cytotoxicity; Secondly, ROS produced by nanoparticles themselves and possibly dissolved metal ions can cause toxicity in E. coli. Moreover, nanoparticles entering cells can stimulate oxidative pathways in cells, resulting in oxidative stress behavior, interfering with protein expression in cells, or post-transcriptional translation [113]. Finally, nanoparticles entering the cell can interact with proteins and their DNA in the cell, causing them to lose certain functions [124]. In previous toxicity studies, a single biological model was typically used to assess or study the toxicity of nanoparticles. Commonly used model organisms have included bacteria [122], algae [125], fish [126], zooplankton [127], mammalian cell lines [128], and so on. Studies have shown that nanoscale particles can seriously impair the physiological activities of cells, subcells, and proteins and even lead to cell death. The main hazards include entry and deposition in the relevant cells (nerve cells, hepatocytes, etc.) [128], protein degeneration and reduced enzyme activity [129], and genotoxicity [130]. DNA mutation and cell membrane damage [131] Oxidative stress [124] and mitochondrial damage [127] affect the expression of functional proteins [132] and immunity [133].
Table 2.
Toxicity studies of typical nanoparticles based on different biological evaluation models.
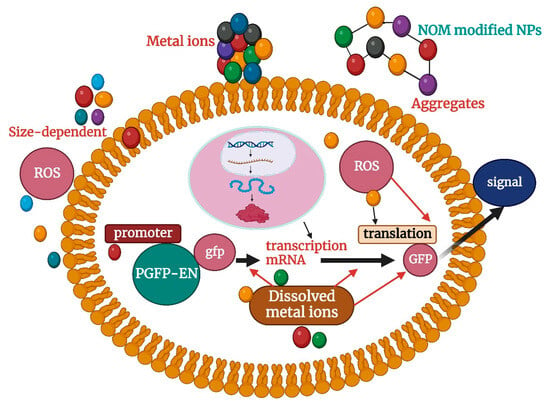
Figure 2.
Toxicological effects of nanoparticles on Escherichia coli.
ROS represents engineering (such as hydroxyl radical •OH, active oxygen singlet oxygen O2−, etc.); NOM stands for natural organic matter; NPs stands for nanoparticle.
Surface changes, antioxidant co-administration, and tailored medication delivery have all been investigated as methods to reduce NP toxicity. Surface customization of NPs with biocompatible ligands or polymers can alter their interaction with cells and lessen toxicity. It has been demonstrated that NP-induced oxidative stress can be reduced by the co-administration of antioxidants such as vitamin C or N-acetyl cysteine. Additionally, the development of NPs for targeted drug delivery offers hope for reducing side effects while enhancing therapeutic efficacy [164].
3.1. Plants and Cyanobacteria Cellular Reactions to Exposure to Metal Nanoparticles
Electronics, healthcare, and agriculture have undergone radical change due to the rapid development and commercialization of metal nanoparticles (NPs). However, given the frequency with which NPs are used, there are worries regarding possible biological and environmental effects. Due to their crucial ecological roles, plants and cyanobacteria in particular have drawn a lot of interest to the interactions between metal NPs and living organisms. Plants are the principal producers that support terrestrial ecosystems by affecting carbon sequestration, soil structure, and nutrient cycling [165].
As the most common photoautotrophic microorganisms in aquatic environments, cyanobacteria make a considerable contribution to the world’s nitrogen and carbon cycles. Understanding the effects of metal NPs on plants and cyanobacteria is essential for predicting and minimizing possible disruptions to ecosystem function, as metal NPs can easily enter terrestrial and aquatic systems [166].
Silver (Ag), gold (Au), zinc oxide (ZnO), and iron oxide (Fe3O4) are only a few examples of the wide variety of materials that make up metal NPs, each with unique physicochemical characteristics. Plants and cyanobacteria use various methods, including NP size, surface charge, and environmental factors, to absorb metal NPs. NPs have the ability to cross cell membranes and walls in plants to access intracellular compartments. Because of their straightforward cell structure, cyanobacteria can directly absorb NPs by passive diffusion or active transport [167].
3.2. Oxidative Stress and Its Implications for Plant-Cyanobacteria Health
In the fields of plant biology and microbial ecology, oxidative stress, defined as the excessive accumulation of ROS beyond the cellular antioxidant capacity, has attracted a lot of attention [168]. Because they are sessile organisms, plants are regularly exposed to a variety of environmental stresses, from biotic variables like herbivores, diseases, and symbiotic bacteria to abiotic factors such as light intensity, temperature changes, and nutrition availability. Cyanobacteria, which can perform photosynthesis and nitrogen fixation, are among these symbiotic partners and create complex relationships with numerous plant species throughout varied habitats [169].
Superoxide anion (O2), hydrogen peroxide (H2O2), and hydroxyl radicals (•OH) are three examples of ROS that are produced naturally during aerobic metabolism and photosynthesis [170]. ROS are crucial signaling molecules in low quantities, but when they build up excessively, they can cause cellular damage and oxidative stress. To maintain redox equilibrium and fight oxidative stress, plants and cyanobacteria have developed complex defense systems that include enzymatic antioxidants (such as superoxide dismutase, catalase, and peroxidases) and non-enzymatic antioxidants (such as ascorbate, glutathione). Nevertheless, a variety of external stressors can upset the equilibrium between ROS generation and scavenging, leading to oxidative stress-related cellular damage [171].
3.3. Signaling Pathways Triggered by Nanoparticle-Induced Stress in Mutualistic Systems
In species from numerous taxonomic groups, exposure to nanoparticles can impair redox balance, disturb cellular homeostasis, and induce a variety of stress reactions [172]. These stressors then trigger a complicated network of signaling channels that control defense and adaptation systems. Mutualistic partners can communicate and control their physiological reactions in the face of stress caused by NP thanks to the complex crosstalk between these pathways [173]. The essential elements of the complex signaling networks triggered in mutualistic organisms under NP stress include reactive oxygen species (ROS) signaling, hormone-mediated pathways, and phytochemical cascades. These signaling pathways help to shape the overall outcome of reciprocol interactions in the presence of NPs, in addition to mediating partner-specific responses [174].
4. Enhancement of Nutrient Uptake and Availability
Growth, development, and production are crucially influenced by how effectively cyanobacteria and plants acquire and use vital nutrients. However, the availability of nutrients, particularly micronutrients, in soil and aquatic habitats is often limited, leading to suboptimal physiological conditions and lower yields [175]. Metal nanoparticles (MNPs) have drawn a lot of interest due to their distinct physicochemical characteristics and prospective uses in biotechnology and agriculture. In order to overcome the problems caused by nutrient deficiency and improve the nutritional status of plants and cyanobacteria, exploiting the ability of MNPs to improve nutrient uptake and availability has great potential [113].
4.1. MNPs as Carriers of Essential Nutrients: Root Interactions and Cyanobacterial Associations
Due to their high surface-to-volume ratio and nanoscale size, MNPs have improved reactivity and solubility, making them efficient transporters of vital nutrients [176]. Root interactions are important for nutrient uptake, and MNPs can alter rhizosphere conditions to change the solubility and availability of nutrients. In addition, cyanobacteria can create complex relationships with plants because they are skilled nitrogen fixers and extracellular polymer makers. The addition of MNPs to these connections has the potential to increase nutrient transfer and build cooperative networks of nutrient exchange, improving nutrient flow and bioavailability [177].
4.2. Nutrient-Use Efficiency and Crop Yield Enhancement through Plant-Cyanobacteria Synergy
To ensure sustainable crop production and reduce environmental consequences, modern agriculture must prioritize optimizing nutrient use efficiency [178]. A promising method for reaching this goal is the symbiotic relationship between plants and cyanobacteria. MNPs can increase nutrient delivery and uptake, which can intensify the advantages of plant-cyanobacteria synergy. MNPs are carriers of both critical nutrients and bioactive chemicals. This interaction not only promotes efficient nutrient utilization but also increases crop production and quality, addressing the issue of global food security [179].
4.3. Ecological and Agricultural Implications
MNP inclusion in nutrient management plans has significant ecological and agricultural effects. MNPs can be used as precise instruments in agroecosystems to address certain nutrient deficiencies and reduce the overuse of traditional fertilizers [180]. In addition, the use of MNPs in ecologically delicate locations can encourage nutrient cycling that is sustainable, lowering the risk of eutrophication and nutrient runoff. However, the ecological effects of MNPs need to be carefully considered, and their long-term effects on soil health, microbial populations, and ecosystem dynamics must be evaluated holistically [181].
5. Stress Tolerance and Disease Resistance
Beyond symbiotic interactions, the complicated dance between plants and cyanobacteria includes mechanisms that allow these organisms to endure stress and fend off pathogenic threats. Partnerships between plants and cyanobacteria have drawn interest because of their ability to ameliorate adverse environmental conditions and increase crop productivity [182]. Agricultural systems face severe challenges from stressors, including salt and drought, demanding creative strategies to increase stress tolerance. Furthermore, the necessity of strong defense mechanisms is highlighted by the sensitivity of plants and cyanobacteria to pathogenic agents [183].
5.1. Nanoparticle-Mediated Stress Responses
Recent studies have shown how nanoparticles can be used to control stress responses in plant-cyanobacteria systems. Due to their distinctive physicochemical characteristics, nanoparticles have been demonstrated to have an impact on a number of biological functions, including stress-related signaling pathways [184]. By boosting antioxidant defense mechanisms and osmoprotectant accumulation, nanoparticle therapies have been suggested as viable ways to improve stress tolerance. Notably, nanoparticles have been linked to the control of gene expression networks associated with stress responses, thereby strengthening stress tolerance mechanisms [185].
5.2. Reinforcement of Pathogen Defense Mechanisms
Pathogens present considerable obstacles to the interactions between plants and cyanobacteria, in addition to abiotic stresses. Different diseases have the potential to upset the delicate balance of these alliances, which would limit plant development and damage cyanobacterial activity [186]. According to recent research, nanoparticle treatments may cause both partners to produce antimicrobial chemicals and upregulate defense-related genes. By priming defense mechanisms through the use of nanoparticles, disease resistance is increased, preserving the stability of plant-cyanobacterial systems [187].
5.3. Mitigation of Abiotic Stress Factors
The adaptability and durability of plant-cyanobacteria systems are demonstrated by their versatility in reducing abiotic stress effects. In many different agricultural environments, salt and drought are common stressors that can seriously hinder development and productivity. By improving water retention and nutrient uptake, nanoparticle therapies have been demonstrated to lessen the negative impacts of abiotic stresses and maintain cellular homeostasis in both plants and cyanobacteria. The potential of nanoparticle interventions to ensure the stability and productivity of plant-cyanobacteria interactions is highlighted by a multimodal approach to stress mitigation [188].
6. Impact on Key Plant and Cyanobacterial Processes
By facilitating energy flow and nutrient cycling, plants and cyanobacteria, as primary producers, play critical roles in ecosystem functioning [189]. However, because their performance is closely linked to current environmental circumstances, they are vulnerable to several stresses. The development, metabolism, and reproduction of plants and cyanobacteria are significantly impacted by environmental stressors such as high temperatures, pollution, and changed nutrient availability [190].
6.1. Impact on Photosynthesis and Carbon Fixation
In both plants and cyanobacteria, photosynthesis is essential for obtaining energy and assimilating carbon. However, stress-related modifications to the photosynthetic apparatus, such as changes in pigment composition and electron transport, can seriously impair the efficiency of energy conversion [191]. Additionally, the complex regulatory networks that control carbon fixation are prone to disruption under stress, leading to imbalances in the distribution of fixed carbon to crucial biomolecules [192].
The development and metabolic processes of plants and cyanobacteria are significantly influenced by the availability of critical nutrients. Elements can be out of balance due to stress-induced changes in nutrition absorption systems and assimilation pathways, endangering cellular homeostasis and overall fitness [193].
6.2. Effects of Nanoparticles on Photosynthesis, Energy Conversion, and Nitrogen Fixation by Cyanobacteria
The process of photosynthesis, which transforms light energy into chemical energy, is a crucial biological phenomenon that powers the ecosystems on Earth. In order to increase the efficiency of photosynthetic production in cyanobacteria, nanoparticles have been studied as potential modulators of light absorption and energy transfer. It is possible to improve light-harvesting complexes and fine-tune photosynthetic pigments through the controlled transport of nanoparticles to cyanobacterial cells, which will ultimately increase energy absorption and conversion [194].
Cyanobacteria have a surprising variety of metabolic pathways, including the storage and conversion of energy. Through better electron transport chains and improved synthesis of energy-rich compounds, it was investigated whether nanoparticles can support bioenergy production. Furthermore, nanoparticle-induced changes in cellular redox balance and electron flow have the ability to redirect metabolic flows toward desired end products [91].
6.3. Effects on Root Architecture, Mycorrhizal Associations, and Cyanobacterial Aggregates
Key elements of terrestrial ecosystems include root architecture, mycorrhizal connections, and cyanobacterial aggregates, which affect nutrient intake, soil structure, and plant health. Understanding these variables’ complex relationships is crucial for ensuring the resilience and sustainability of ecosystems. The spatial arrangement of roots in the soil is influenced by root architecture, which also affects nutrient and water uptake [195]. Root growth patterns are influenced by environmental factors such as gravity, moisture, and nutrient availability. A growing body of evidence points to the crucial role that microorganisms, such as mycorrhizal fungi and cyanobacteria, play in directing root branching, elongation, and tip growth [196].
The process of domestication has significantly altered crop root configuration, including, but not limited to, root branching, length, biomass, volume, anatomical structure, and surface area [197], which influence the recruitment and colonization of root microbial communities in wild and modern crop varieties [198] (Figure 3). Under modern intensive farmland management with high density and high nutrient input, crop domestication usually leads to fewer fine roots, shallower roots, increased lateral roots, and a lower root-to-stem ratio of crops [199,200] and this can have a profound impact on root microbes. For example, [201] showed that fine roots (or root hairs) of plants enrich more diverse microbial communities than coarse roots, and the abundance of root microorganisms in fine roots is generally higher than that in coarse roots. The authors of [202] also found that fine roots were more likely to promote microbial growth, while [203] found that the absence of root hair significantly reduced the complexity of rhizosphere microbial communities. The authors of [204] found that the root system of domesticated modern kidney beans was “coarser” than that of wild kidney beans, resulting in a significantly lower relative abundance of Bacteroides in the rhizosphere microorganisms of modern kidney beans. Therefore, the reduction of fine roots caused by domestication may be an important reason why the diversity of root bacteria and fungi of most modern crop varieties is lower than that of their wild ancestors.
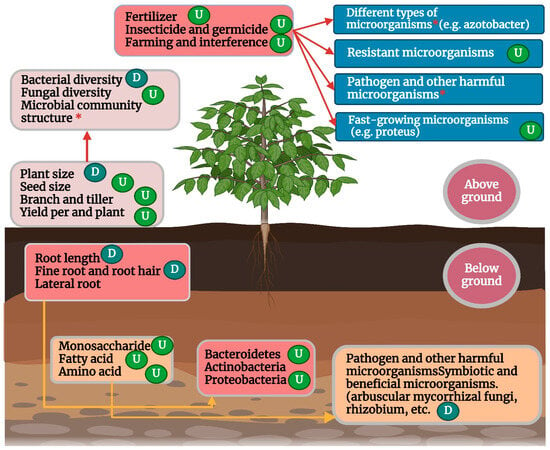
Figure 3.
Effects of domestication on microbiome community composition and diversity of crops (U) indicate significant increase and (D) decrease, respectively; (*) indicates a significant difference.
7. Phytoremediation Potential
Anthropogenic environmental contamination has become a global issue, calling for creative and long-lasting restoration techniques. An engineering method called phytoremediation uses the inherent capacities of plants and the microbes that live with them to mitigate various pollutants. This paper offers a thorough summary of the potential, methods, applications, difficulties, and recent developments in phytoremediation. Rhizofiltration, phytostabilization, phytoextraction, and phytovolatilization are only a few of the many mechanisms involved in phytoremediation. Through their root systems, plants can absorb, change, immobilize, or destroy contaminants, thereby cleansing the environment. Numerous studies have shown how these processes work in various plant species and polluted environments [205].
Plant-microbe interactions are essential to the efficacy of phytoremediation. The rhizosphere’s microorganisms support pollutant transformation and nutrient uptake, improving plant health and pollutant degradation. Recent research has emphasized the value of rhizobacterial populations and mycorrhizal connections in boosting phytoremediation effectiveness [206].
The efficacy of phytoremediation has improved thanks to developments in genetic engineering. Increased pollutant uptake, tolerance, and degradation rates may be due to a genetic change. The development of hyperaccumulator plants specifically suited to environmental remediation is possible thanks to advances in synthetic biology. Plants that overexpress metal transporters and enzymes involved in pollution metabolism have been successfully created by researchers [207].
7.1. Mechanisms of Phytoremediation Assisted by Metal Nanoparticles
Metal nanoparticles interact with plants and their rhizosphere through a variety of methods, including increased adsorption surface area, enhanced metal uptake through root penetration, and accelerated translocation in plant tissue. These interactions often result in increased plant development, reduced metal toxicity, and improved metal sequestration. One of the main factors affecting the effectiveness of nanoparticles is how they affect metal speciation and availability in soil [208].
7.2. Synergistic Effects on Plant-Microbe Interactions
In addition to plant physiology, metal nanoparticles also influence the composition and activity of rhizospheric microbial communities. These adaptations can lead to increased metal mobilization and transformation, increased plant stress tolerance, and increased nutrient availability. The potential for nanoparticle-assisted phytoremediation to increase the overall effectiveness of pollutant removal is highlighted by the complex interplay between nanoparticles, plants, and microorganisms [209].
Numerous metal-contaminated areas have been successfully remediated using metal nanoparticle-assisted phytoremediation. Numerous metal nanoparticles, including zero-valent iron, iron oxide, and titanium dioxide, have been the subject of case studies that have demonstrated their efficacy in accelerating the buildup and removal of metals from contaminated soils and streams. These studies give important information on how this strategy can be used in practice [210].
7.3. Ecological Considerations, Cyanobacterial Contributions, and Long-Term Effectiveness
Despite the positive results, there are still issues that need to be resolved, including the stability of nanoparticles, possible ecotoxicological effects, and regulatory issues. The fat and behavior of nanoparticles in the environment, as well as their long-term implications for ecosystems, are still being researched. The development of sustainable nanoparticle-assisted phytoremediation solutions must take into account the optimization of nanoparticle synthesis technologies and the assessment of their environmental effects [211].
8. Cutting-Edge Developments in Agricultural Nanotechnology
Population growth, climate change, and a decline in arable land pose growing problems for agriculture. By exploiting the special properties of nanoparticles to improve plant growth, nutrient supply, and pest control, agricultural nanotechnology offers promising answers. Nanoparticles are receiving attention because of their potential to improve soil structure, nutrient retention, and water availability. Examples include nanoclays, metal nanoparticles, and carbon-based nanomaterials [212]. The potential of nanomaterials to increase crop production and stress tolerance has been demonstrated. Man-made nanoparticles such as silicon and zinc oxide nanoparticles have been shown to improve plant growth, photosynthesis, and nutrient absorption [213]. Agrochemical nanoformulations such as insecticides and fertilizers enable controlled release and targeted delivery, reducing the amount of chemicals required and minimizing environmental impact [214]. By providing real-time information about soil moisture, nutrient levels, and pest infestation, nanotechnology-enabled sensors play a critical role in precision agriculture. These sensors, often combined with data analytics and wireless networks, help farmers make informed decisions, maximize resource utilization, and reduce input waste [215]. Furthermore, nanoscale biosensors are being developed to quickly detect plant diseases and ensure immediate intervention to reduce crop losses [216].
8.1. Integrating Cyanobacterial Solutions
Due to their ability to accumulate lipids and carbohydrates, cyanobacteria are potential candidates for biofuel production. The main objectives of the research were to improve pathways for lipid production and increase biomass productivity through genetic engineering and cultivation techniques. Due to improvements in metabolic engineering, strains of cyanobacteria capable of producing biofuels such as biodiesel and bioethanol have been created [217]. It is important to highlight the role that cyanobacteria play in reducing carbon dioxide levels and helping to adapt to climate change. Although cyanobacteria blooms are often considered destructive, they have the ability to store carbon. Cyanobacteria provide a natural solution for carbon capture and storage by absorbing atmospheric carbon and incorporating it into their biomass [218]. The use of cyanobacteria in environmentally friendly treatment methods is increasing in popularity due to their ability to absorb nutrients and toxins from wastewater. Photobioreactors and constructed wetlands are two examples of cyanobacteria-based wastewater treatment technologies that provide an environmentally friendly method of nutrient removal and water filtration. These systems produce biomass that, in addition to wastewater treatment, is also suitable for the production of bioenergy [219].
8.2. Cyanobacterial Biofertilizers and Soil Fertility
Cyanobacteria are essential for soil nutrient enrichment and the nitrogen cycle. Improved soil fertility and plant development are the result of their capacity to fix atmospheric nitrogen and secrete chemicals that encourage growth. According to research, using cyanobacterial biofertilizers can increase agricultural yields, especially in soils with low nitrogen levels. Additionally, the interactions between cyanobacteria and soil microbes can create a favorable rhizosphere environment, enhancing nutrient availability and plant health [220]. The overuse of artificial fertilizers has sparked worries about resource depletion and environmental degradation. A sustainable solution is provided by cyanobacterial biofertilizers, which increase the effectiveness of nutrient use and decrease the need for synthetic fertilizers. Studies have shown that cyanobacterial biofertilizers can improve nutrient uptake, maximize fertilizer use, and reduce nutrient leaching, thereby reducing the adverse impacts of nutrient runoff on the environment [221]. Precision farming methods allow for site-specific management of agricultural inputs thanks to developments in remote sensing, geographic information systems (GIS), and data analytics. By combining cyanobacterial biofertilizers with precision farming, it is possible to apply them in a way that is specifically matched to the nutrient needs of various crop zones. This strategy maximizes the advantages of using biofertilizers while optimizing nutrient distribution, cutting waste, and reducing waste [222]. It has been demonstrated that cyanobacterial biofertilizers increase crop tolerance to a variety of abiotic conditions, including salt, drought, and heavy metal toxicity. Along with greater nutritional availability, the growth-promoting compounds released by cyanobacteria help plants develop stronger stress resistance systems. Combining precision farming methods with the use of biofertilizers can help identify areas that are more vulnerable to stress and perform tailored interventions to lessen its effects [223].
8.3. Role of Cyanobacteria in Nanoparticle Applications
Cyanobacteria have the innate ability to synthesize nanoparticles by reducing metal ions. A specific method for regulating the size, shape, and composition of nanoparticles is enabled by their capacity to metabolize a variety of substrates and modulate biological functions. Recent studies have shown that cyanobacteria are capable of biosynthesizing metallic, metal oxide, and semiconductor nanoparticles, which have uses in photonics, catalysis, and other fields [224].
Nanoparticles must have their surfaces functionalized in order to improve their stability, biocompatibility, and specific uses. Through the use of genetic engineering or environmental modification, cyanobacteria provide a biogenic substrate for the synthesis of nanoparticles with customized surface properties. This provides opportunities for the development of bio-inspired, biocompatible nanoparticles with improved capabilities for imaging in medicine, drug delivery, and environmental sensing [225].
8.4. Ethical Considerations and Regulatory Implications
The use of cyanobacteria for the synthesis of nanoparticles raises ethical questions about the release of contaminants into the environment, unforeseen consequences, and potential hazards to ecosystems and human health. Rigorous risk assessment, transparency, and adherence to regulatory frameworks are necessary to strike a balance between technological growth and responsible innovation. Consideration must also be given to the moral ramifications of patenting biogenic nanoparticles produced by natural processes. Collaborations between scientists, politicians, ethicists, and stakeholders across other disciplines are necessary to strike a balance between the advantages of cyanobacteria-mediated nanoparticle applications and ethical issues. The development of open guidelines, encouraging public participation, and advancing sustainable innovation through well-informed decision-making should be the main goals of responsible research and innovation [226].
9. Current Trends in Agricultural Nanotechnology
The need to enhance food production to meet the demands of an expanding global population while reducing environmental effects is one of the many difficulties facing agriculture. By offering methods to improve agricultural processes at the nanoscale, nanotechnology offers creative responses to these problems. Crop yields, resource use, and overall agricultural sustainability can be improved by nanoscale manipulation of materials and structures [227].
The development and use of nanoparticles for crop improvement are two major trends in agricultural nanotechnology. The potential for improving nutrient uptake, enhancing plant growth, and reducing stress factors has been demonstrated by nanoparticles such as nanopesticides and nanofertilizers. By directly delivering nutrients to plant cells, these nanoparticles can improve nutrient uptake efficiency while lowering environmental contamination [228].
By giving soil managers access to real-time information on soil moisture, nutrient levels, and microbial activity, nanoscale sensors play a crucial role in soil management. These nanosensors provide farmers with exact information to optimize irrigation and fertilization practices, thereby improving resource management and increasing crop output. Additionally, nanoparticles can be employed to enhance soil structure and water retention, promoting long-term sustainability and soil health [229]. Several recent studies have demonstrated the significant potential of nanotechnology in enhancing crop productivity, resource utilization, and environmental impacts in the context of precision and sustainable agriculture. These studies have explored various nanotechnological interventions, such as foliar and soil bio-augmentation, to address challenges in agriculture, including climate change, soil contamination, and increasing food demands. The application of nanotechnology in agriculture is increasingly recognized as a critical aspect of precision farming, offering new solutions to combat existing problems and improve modern farming practices. Figure 4 provides a diagrammatic representation of the application of nanotechnology in precise and sustainable agriculture and current trends in the field. The interaction between plants and nanomaterials is the focus of these studies and highlights the potential for developing novel and desirable materials for crop improvement. While the use of nanotechnology in agriculture holds promise for sustainable green agriculture, it also presents challenges such as nanotoxicity and environmental impact, which require careful regulation and long-term evaluation.
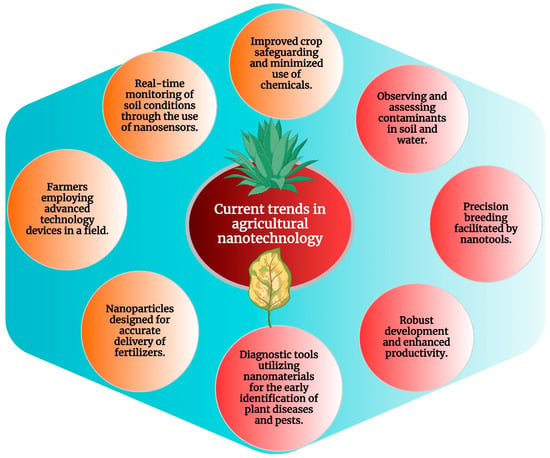
Figure 4.
Current trends in agricultural nanotechnology.
9.1. Integrating Metal Nanoparticles into Modern Agricultural Practices
It has been demonstrated that adding metal nanoparticles, such as copper, gold, and silver, can improve plant growth, disease resistance, and nutrient uptake. The antibacterial capabilities of metal nanoparticles can reduce plant diseases and enhance crop health [230]. They have shown improved nutrient efficiency and decreased chemical discharge in nanopesticides and nanofertilizers, which support sustainable agriculture practices [231]. Additionally, the sustained effects of the controlled release of metal nanoparticles from nanomaterials eliminate the need for repeated applications. Modern agricultural techniques that incorporate metal nanoparticles have a significant promise for increasing yields while reducing their negative environmental effects [232].
9.2. Integrating Cyanobacterial Solutions
Photosynthetic microorganisms, called cyanobacteria, provide a sustainable solution to agricultural problems. Opportunities for biofertilization and promoting plant growth are provided by their capacity to fix atmospheric nitrogen and produce bioactive chemicals [233]. It has been shown that cyanobacterial inoculants increase soil fertility, nutrient availability, and water retention [220]. Additionally, cyanobacteria support soil health and microbial diversity by enhancing soil structure through the production of exopolymeric compounds. A natural and environmentally benign method of increasing agricultural output while lowering chemical inputs is the incorporation of cyanobacterial solutions [234].
9.3. Synergies with Precision Farming, Sustainable Agriculture, and Cyanobacterial Biofertilizers
The combination of cyanobacterial biofertilizers, precision farming, and agricultural nanotechnology has enormous promise for maximizing resource usage and reducing environmental impact. Utilizing real-time data, precision farming methods such as remote sensing and GPS-guided machinery allow for the targeted application of nanomaterials and cyanobacterial solutions [235]. This interaction improves crop quality, lowers waste, and improves fertilizer management. Additionally, the combination of cyanobacterial biofertilizers and precision farming promotes long-term agricultural sustainability by improving the health and fertility of the soil. The joint application of these technologies represents a major step towards a future where agriculture is more effective and environmentally sensitive [236].
9.4. Balancing Benefits, Ethical Concerns, and the Pivotal Role of Cyanobacteria in Nanoparticle Applications
It is quite possible to maximize resource use and reduce environmental effects by combining agricultural nanotechnology, precise farming, and cyanobacterial biofertilizers. The focused application of nanomaterials and cyanobacterial solutions is enabled by precision farming techniques, such as remote sensing and GPS-controlled devices, based on real-time data [237]. This interaction optimizes fertilizer management, lowers waste, and raises crop quality. Additionally, the use of biofertilizers derived from cyanobacteria in conjunction with precision farming promotes long-term soil fertility and health. A substantial step towards a more effective and environmentally responsible agricultural future has been made by the coordinated use of these technologies [238].
10. Future Prospects and Challenges
10.1. Unexplored Areas of Research in Plant-Cyanobacteria-Nanoparticle Interactions
Although tremendous progress has been made in understanding the interactions between cyanobacteria and plants, little is known about how nanoparticles may affect this complex relationship. Recent studies demonstrate that nanoparticles and cyanobacterial bioinoculants may work in concert to improve plant growth, stress tolerance, and nutrient uptake. However, further research is needed to determine the underlying mechanisms and precise effects of nanoparticles on cyanobacterial symbiosis and subsequent plant responses. Opportunities exist to decipher innovative mechanisms that may support sustainable agriculture and environmental health in this unexplored zone [239].
10.2. Regulatory Aspects, Ethical Considerations, and the Potential of Cyanobacterial Bioinoculants
The ability of cyanobacterial bioinoculants to fix atmospheric nitrogen, improve soil fertility, and stimulate plant growth highlights their potential to revolutionize sustainable agriculture. The use of bioinoculants, such as cyanobacteria, is subject to growing regulatory frameworks that seek to assure product efficacy, safety, and minimal environmental impact. Biosafety, intellectual property rights, and the equal sharing of rewards are all ethical issues. It is crucial to strike a balance between innovation and responsible use. By lowering the use of chemical inputs, preserving resources, and enhancing soil health, cyanobacterial bioinoculants have the potential to revolutionize agricultural practices; however, their integration must be achieved within strict regulatory and ethical frameworks [239,240].
10.3. Steps toward Responsible and Safe Utilization of Nanoparticles in a Cyanobacteria-Augmented Environment
A cautious approach is required when the agricultural world investigates the incorporation of nanoparticles into systems enhanced with cyanobacteria. To examine potential nontoxicity, effects on unintended organisms, and long-term repercussions on soil and aquatic ecosystems, thorough risk assessments must be carried out. Unintended consequences can be reduced by including life cycle analyses and creating standardized methods for nanoparticle formulation and use. To create policies that encourage the responsible use of nanoparticles in cyanobacteria-augmented environments while minimizing ecological and human health hazards, collaboration between scientists, policymakers, and stakeholders is crucial [147].
11. Conclusions
In the field of agricultural science, the combination of nitrogen-fixing cyanobacteria and nanoparticle-based methods holds great promise for improving crop yields. This study explores the detailed molecular processes that make this partnership effective and how it can be used in farming. Nitrogen-fixing cyanobacteria have evolved special traits that help them deal with low levels of nitrogen in the soil. Meanwhile, nanoparticles provide new ways to deliver nutrients to plants and make them stronger. By closely studying how these two work together, from the tiniest levels of molecules to larger ecosystems, we learn more about how they can boost crop growth and output. In the forthcoming stages, the amalgamation of cyanobacteria and nanoparticles demonstrates promise in fostering sustainable agricultural methodologies, reducing dependency on synthetic fertilizers, and mitigating environmental impacts. Nevertheless, comprehensive scrutiny of the ecological and ethical ramifications of these methodologies is indispensable to guarantee prudent implementation and stewardship of agricultural resources. To conclude, the amalgamation of cyanobacteria and nanoparticles unveils an enticing frontier for agricultural progression, providing avenues for heightened efficiency and ecological sustainability in food production. Sustained research efforts and thoughtful consideration are essential as we traverse the dynamic terrain of agricultural innovation.
Author Contributions
Conceptualization: T.N., S.F., R.Z. and L.G.; Writing original draft: R.Z. and S.S.; Writing—review and editing: T.N., S.F., and B.B. All authors have read and agreed to the published version of the manuscript.
Funding
The authors would like to extend their sincere appreciation to the USDA Hatch Project SD00H691-20 titled Engineering Solar-powered N2-fixing Cyanobacteria for Agricultural and Industrial Applications (to R.Z.).
Acknowledgments
This acknowledgement recognizes the contribution of Biorender in creating the figures for the paper and highlights the platform’s role in enhancing the visual presentation of scientific information. The authors would like to extend their sincere appreciation to the USDA Hatch Project SD00H691-20 titled Engineering Solar-powered N2-fixing Cyanobacteria for Agricultural and Industrial Applications (to R.Z.).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gu, J.; Yang, J. Nitrogen (N) transformation in paddy rice field: Its effect on N uptake and relation to improved N management. Crop Environ. 2022, 1, 7–14. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant associated rhizobacteria for biocontrol and plant growth enhancement. Front. Plant Sci. 2021, 12, 634796. [Google Scholar] [CrossRef] [PubMed]
- Bellenger, J.P.; Darnajoux, R.; Zhang, X.; Kraepiel, A.M.L. Biological nitrogen fixation by alternative nitrogenases in terrestrial ecosystems: A review. Biogeochemistry 2020, 149, 53–73. [Google Scholar] [CrossRef]
- Smercina, D.N.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. To fix or not to fix: Controls on free-living nitrogen fixation in the rhizosphere. Appl. Environ. Microbiol. 2019, 85, e02546-18. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.; Bosch, J.; Coclet, C.; Johnson, J.; Lebre, P.; Salawu-Rotimi, A.; Vikram, S.; Makhalanyane, T.; Cowan, D. Microbial nitrogen cycling in Antarctic soils. Microorganisms 2020, 8, 1442. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sangwan, S.; Kaur, H.; Patra, A.; Anamika; Mehta, S. Diversity and evolution of nitrogen fixing bacteria. In Sustainable Agriculture Reviews 60: Microbial Processes in Agriculture; Springer: Berlin/Heidelberg, Germany, 2023; Volume 60, pp. 95–120. [Google Scholar]
- Nawaz, T.; Gu, L.; Fahad, S.; Saud, S.; Harrison, M.T.; Zhou, R. Sustainable protein production through genetic engineering of cyanobacteria and use of atmospheric N2 gas. Food Energy Secur. 2024, 13, e536. [Google Scholar] [CrossRef]
- Hu, C.; Rzymski, P. Non-photosynthetic Melainabacteria (Cyanobacteria) in human gut: Characteristics and association with health. Life 2022, 12, 476. [Google Scholar] [CrossRef] [PubMed]
- Runge, E.A.; Mansor, M.; Kappler, A.; Duda, J.P. Microbial biosignatures in ancient deep-sea hydrothermal sulfides. Geobiology 2023, 21, 355–377. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, T.; Gu, L.; Fahad, S.; Saud, S.; Hassan, S.; Harrison, M.T.; Liu, K.; Zhou, R. Unveiling the antioxidant capacity of fermented foods and food microorganisms: A focus on cyanobacteria. J. Umm Al-Qura Univ. Appl. Sci. 2024, 10, 232–243. [Google Scholar] [CrossRef]
- Callieri, C.; Cabello-Yeves, P.J.; Bertoni, F. The “dark side” of picocyanobacteria: Life as we do not know it (yet). Microorganisms 2022, 10, 546. [Google Scholar] [CrossRef]
- Mallén-Ponce, M.J.; Huertas, M.J.; Florencio, F.J. Exploring the diversity of the thioredoxin systems in cyanobacteria. Antioxidants 2022, 11, 654. [Google Scholar] [CrossRef] [PubMed]
- Garlapati, D.; Chandrasekaran, M.; Devanesan, A.; Mathimani, T.; Pugazhendhi, A. Role of cyanobacteria in agricultural and industrial sectors: An outlook on economically important byproducts. Appl. Microbiol. Biotechnol. 2019, 103, 4709–4721. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.P.; Nguyen, L.N.; Zdarta, J.; Nga, T.T.; Nghiem, L.D. Blue-green algae in surface water: Problems and opportunities. Curr. Pollut. Rep. 2020, 6, 105–122. [Google Scholar] [CrossRef]
- Righini, H.; Francioso, O.; Martel Quintana, A.; Roberti, R. Cyanobacteria: A natural source for controlling agricultural plant diseases caused by fungi and oomycetes and improving plant growth. Horticulturae 2022, 8, 58. [Google Scholar] [CrossRef]
- Margesin, R.; Collins, T. Microbial ecology of the cryosphere (glacial and permafrost habitats): Current knowledge. Appl. Microbiol. Biotechnol. 2019, 103, 2537–2549. [Google Scholar] [CrossRef] [PubMed]
- Kollmen, J.; Strieth, D. The beneficial effects of cyanobacterial co-culture on plant growth. Life 2022, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Pernil, R.; Schleiff, E. Metalloproteins in the Biology of Heterocysts. Life 2019, 9, 32. [Google Scholar] [CrossRef]
- Alleman, A.B.; Peters, J.W. Mechanisms for Generating Low Potential Electrons across the Metabolic Diversity of Nitrogen-Fixing Bacteria. Appl. Environ. Microbiol. 2023, 89, e00378-23. [Google Scholar] [CrossRef]
- Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting biological nitrogen fixation: A route towards a sustainable agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef]
- Carvalho, S.; Kürten, B.; Krokos, G.; Hoteit, I.; Ellis, J. The red sea. In World Seas: An Environmental Evaluation; Academic Press: Cambridge, MA, USA, 2019; pp. 49–74. [Google Scholar]
- Williamson, C.E.; Neale, P.J.; Hylander, S.; Rose, K.C.; Figueroa, F.L.; Robinson, S.A.; Häder, D.P.; Wängberg, S.Å.; Worrest, R.C. The interactive effects of stratospheric ozone depletion, UV radiation, and climate change on aquatic ecosystems. Photochem. Photobiol. Sci. 2019, 18, 717–746. [Google Scholar] [CrossRef]
- McGregor, G.B.; Sendall, B.C. Iningainema pulvinus gen nov., sp nov. (Cyanobacteria, Scytonemataceae) a new nodularin producer from Edgbaston Reserve, north-eastern Australia. Harmful Algae 2017, 62, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Galloway, J.N.; Aber, J.D.; Erisman, J.W.; Seitzinger, S.P.; Howarth, R.W.; Cowling, E.B.; Cosby, B.J. The nitrogen cascade. Bioscience 2003, 53, 341–356. [Google Scholar] [CrossRef]
- Fadeel, B.; Garcia-Bennett, A.E. Better safe than sorry: Understanding the toxicological properties of inorganic nanoparticles manufactured for biomedical applications. Adv. Drug Deliv. Rev. 2010, 62, 362–374. [Google Scholar] [CrossRef]
- Rai, M.K.; Deshmukh, S.D.; Ingle, A.P.; Gade, A.K. Silver nanoparticles: The powerful nanoweapon against multidrug-resistant bacteria. J. Appl. Microbiol. 2012, 112, 841–852. [Google Scholar] [CrossRef]
- Zaman, M.A.; Abbas, R.Z.; Qamar, W.; Qamar, M.F.; Mehreen, U.; Shahid, Z.; Kamran, M. Role of secondary metabolites of medicinal plants against Ascaridia galli. Worlds Poult. Sci. J. 2020, 76, 639–655. [Google Scholar] [CrossRef]
- Huq, M.E.; Fahad, S.; Shao, Z.; Sarven, M.S.; Khan, I.A.; Alam, M.; Saeed, M.; Ullah, H.; Adnan, M.; Saud, S.; et al. Arsenic in a groundwater environment in Bangladesh: Occurrence and mobilization. J. Environ. Manag. 2020, 262, 110318. [Google Scholar] [CrossRef]
- Chen, D.; Mubeen, B.; Hasnain, A.; Rizwan, M.; Adrees, M.; Naqvi, S.A.H.; Iqbal, S.; Kamran, M.; El-Sabrout, A.M.; Elansary, H.O.; et al. Role of promising secondary metabolites to confer resistance against environmental stresses in crop plants: Current scenario and future perspectives. Front. Plant Sci. 2022, 13, 881032. [Google Scholar] [CrossRef]
- Zahoor, M.; Nazir, N.; Iftikhar, M.; Naz, S.; Zekker, I.; Burlakovs, J.; Uddin, F.; Kamran, A.W.; Kallistova, A.; Pimenov, N.; et al. A review on silver nanoparticles: Classification, various methods of synthesis, and their potential roles in biomedical applications and water treatment. Water 2021, 13, 2216. [Google Scholar] [CrossRef]
- Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of cadmium: Physiological, biochemical, and molecular mechanisms. Biology 2020, 9, 177. [Google Scholar] [CrossRef]
- Hussain, M.; Zahra, N.; Lang, T.; Zain, M.; Raza, M.; Shakoor, N.; Adeel, M.; Zhou, H. Integrating nanotechnology with plant microbiome for next-generation crop health. Plant Physiol. Biochem. 2023, 196, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Medina-Velo, I.A.; Cota-Ruiz, K.; Moreno-Olivas, F.; Gardea-Torresdey, J.L. Can abiotic stresses in plants be alleviated by manganese nanoparticles or compounds? Ecotoxicol. Environ. Saf. 2019, 184, 109671. [Google Scholar] [CrossRef] [PubMed]
- Zahra, Z.; Habib, Z.; Hyun, H.; Shahzad, H.M.A. Overview on recent developments in the design, application, and impacts of nanofertilizers in agriculture. Sustainability 2022, 14, 9397. [Google Scholar] [CrossRef]
- do Espirito Santo Pereira, A.; Caixeta Oliveira, H.; Fernandes Fraceto, L.; Santaella, C. Nanotechnology potential in seed priming for sustainable agriculture. Nanomaterials 2021, 11, 267. [Google Scholar] [CrossRef]
- Kumar, B.; Singhal, R.K.; Chand, S.; Chauhan, J.; Kumar, V.; Mishra, U.N.; Hidangmayum, A.; Singh, A.; Bose, B. Nanopriming in sustainable agriculture: Recent advances, emerging challenges and future prospective. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 339–365. [Google Scholar] [CrossRef]
- Garg, R.; Maheshwari, S. Seed Priming and Fortification of Seeds Using Nanotechnology: A review. EPH—Int. J. Environ. Agric. Res. 2023, 9, 11–17. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of metal and metal oxide nanoparticles on plant: A critical review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Mitra, D.; Adhikari, P.; Djebaili, R.; Thathola, P.; Joshi, K.; Pellegrini, M.; Adeyemi, N.O.; Khoshru, B.; Kaur, K.; Priyadarshini, A.; et al. Biosynthesis and characterization of nanoparticles, its advantages, various aspects and risk assessment to maintain the sustainable agriculture: Emerging technology in modern era science. Plant Physiol. Biochem. 2023, 196, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Elmer, W.; White, J.C. The future of nanotechnology in plant pathology. Annu. Rev. Phytopathol. 2018, 56, 111–133. [Google Scholar] [CrossRef]
- Guleria, A.; Sachdeva, H.; Saini, K.; Gupta, K.; Mathur, J. Recent trends and advancements in synthesis and applications of plant-based green metal nanoparticles: A critical review. Appl. Organomet. Chem. 2022, 36, e6778. [Google Scholar] [CrossRef]
- Jain, A.; Ranjan, S.; Dasgupta, N.; Ramalingam, C. Nanomaterials in food and agriculture: An overview on their safety concerns and regulatory issues. Crit. Rev. Food Sci. Nutr. 2018, 58, 297–317. [Google Scholar] [CrossRef]
- Mohamed, N.A.; Marei, I.; Crovella, S.; Abou-Saleh, H. Recent developments in nanomaterials-based drug delivery and upgrading treatment of cardiovascular diseases. Int. J. Mol. Sci. 2022, 23, 1404. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.A.; Syeda, J.T.; Wasan, K.M.; Wasan, E.K. An overview of chitosan nanoparticles and its application in non-parenteral drug delivery. Pharmaceutics 2017, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Jinnouchi, R.; Asahi, R. Predicting catalytic activity of nanoparticles by a DFT-aided machine-learning algorithm. J. Phys. Chem. Lett. 2017, 8, 4279–4283. [Google Scholar] [CrossRef]
- Martínez, G.; Merinero, M.; Pérez-Aranda, M.; Pérez-Soriano, E.M.; Ortiz, T.; Villamor, E.; Begines, B.; Alcudia, A. Environmental impact of nanoparticles’ application as an emerging technology: A review. Materials 2020, 14, 166. [Google Scholar] [CrossRef]
- Nel, A.E.; Mädler, L.; Velegol, D.; Xia, T.; Hoek, E.M.; Somasundaran, P.; Klaessig, F.; Castranova, V.; Thompson, M. Understanding biophysicochemical interactions at the nano–bio interface. Nat. Mater. 2009, 8, 543–557. [Google Scholar] [CrossRef]
- Monteiro-Riviere, N.A.; Tran, C.L. (Eds.) Nanotoxicology: Progress toward Nanomedicine; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar] [CrossRef]
- Schloemer, T.; Narayanan, P.; Zhou, Q.; Belliveau, E.; Seitz, M.; Congreve, D.N. Nanoengineering Triplet–Triplet Annihilation Upconversion: From Materials to Real-World Applications. ACS Nano 2023, 17, 3259–3288. [Google Scholar] [CrossRef]
- Heilgeist, S.; Sekine, R.; Sahin, O.; Stewart, R.A. Finding nano: Challenges involved in monitoring the presence and fate of engineered titanium dioxide nanoparticles in aquatic environments. Water 2021, 13, 734. [Google Scholar] [CrossRef]
- Renn, O.; Roco, M.C. Nanotechnology and the need for risk governance. In Emerging Technologies; Routledge: London, UK, 2020; pp. 321–359. [Google Scholar] [CrossRef]
- Rozaki, Z. Food security challenges and opportunities in Indonesia post COVID-19. Adv. Food Secur. Sustain. 2021, 6, 119–168. [Google Scholar] [CrossRef]
- Majumdar, S.; Keller, A.A. Omics to address the opportunities and challenges of nanotechnology in agriculture. Crit. Rev. Environ. Sci. Technol. 2021, 51, 2595–2636. [Google Scholar] [CrossRef]
- Krishnani, K.K.; Boddu, V.M.; Chadha, N.K.; Chakraborty, P.; Kumar, J.; Krishna, G.; Pathak, H. Metallic and non-metallic nanoparticles from plant, animal, and fisheries wastes: Potential and valorization for application in agriculture. Environ. Sci. Pollut. Res. 2022, 29, 81130–81165. [Google Scholar] [CrossRef]
- El Semary, N.A. Nitrogen-fixing Cyanothece sp. as a mixotroph and silver nanoparticle synthesizer: A multitasking exceptional cyanobacterium. Braz. J. Biol. 2022, 82, e265135. [Google Scholar] [CrossRef]
- Shnoudeh, A.J.; Hamad, I.; Abdo, R.W.; Qadumii, L.; Jaber, A.Y.; Surchi, H.S.; Alkelany, S.Z. Synthesis, characterization, and applications of metal nanoparticles. In Biomaterials and Bionanotechnology; Academic Press: Cambridge, MA, USA, 2019; pp. 527–612. [Google Scholar] [CrossRef]
- Brandelli, A.; Ritter, A.C.; Veras, F.F. Antimicrobial activities of metal nanoparticles. In Metal Nanoparticles in Pharma; Springer: Berlin/Heidelberg, Germany, 2017; pp. 337–363. [Google Scholar] [CrossRef]
- Nawaz, T.; Saud, S.; Gu, L.; Khan, I.; Fahad, S.; Zhou, R. Cyanobacteria: Harnessing the Power of Microorganisms for Plant Growth Promotion, Stress Alleviation, and Phytoremediation in the Era of Sustainable Agriculture. Plant Stress 2024, 11, 100399. [Google Scholar] [CrossRef]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; Rehman, H.; Ashraf, I.; Sanaullah, M. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2020, 721, 137778. [Google Scholar] [CrossRef] [PubMed]
- Burketová, L.; Martinec, J.; Siegel, J.; Macůrková, A.; Maryška, L.; Valentová, O. Noble metal nanoparticles in agriculture: Impacts on plants, associated microorganisms, and biotechnological practices. Biotechnol. Adv. 2022, 58, 107929. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Maldonado, A.; Ortega-Ortíz, H.; Morales-Díaz, A.B.; González-Morales, S.; Morelos-Moreno, Á.; Cabrera-De la Fuente, M.; Sandoval-Rangel, A.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Nanoparticles and nanomaterials as plant biostimulants. Int. J. Mol. Sci. 2019, 20, 162. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Rizvi, A.; Ali, K.; Lee, J.; Zaidi, A.; Khan, M.S.; Musarrat, J. Nanoparticles in the soil–plant system: A review. Environ. Chem. Lett. 2021, 19, 1545–1609. [Google Scholar] [CrossRef]
- Hussain, M.; Shakoor, N.; Adeel, M.; Ahmad, M.A.; Zhou, H.; Zhang, Z.; Xu, M.; Rui, Y.; White, J.C. Nano-enabled plant microbiome engineering for disease resistance. Nano Today 2023, 48, 101752. [Google Scholar] [CrossRef]
- Li, X.; He, F.; Wang, Z.; Xing, B. Roadmap of environmental health research on emerging contaminants: Inspiration from the studies on engineered nanomaterials. Eco-Environ. Health 2022, 1, 181–197. [Google Scholar] [CrossRef] [PubMed]
- De la Torre Roche, R.; Servin, A.; Hawthorne, J.; Xing, B.; Newman, L.A.; Ma, X.; Chen, G.; White, J.C. Terrestrial Trophic Transfer of Bulk and Nanoparticle La2O3 Does Not Depend on Particle Size. Environ. Sci. Technol. 2015, 49, 11866–11874. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Gu, Z.; Pan, Y.; Chen, J.; Xie, Q.; Xu, S.; Gao, M.; Cao, X.; Liu, S.; Wang, W.; et al. Biotransformation of rare earth oxide nanoparticles eliciting microbiota imbalance. Part. Fibre Toxicol. 2021, 18, 17. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, J.; Shang, E.; Xia, X.; Niu, J.; Crittenden, J. Effects of Chloride Ions on Dissolution, ROS Generation, and Toxicity of Silver Nanoparticles under UV Irradiation. Environ. Sci. Technol. 2018, 52, 4842–4849. [Google Scholar] [CrossRef] [PubMed]
- Shang, E.; Li, Y.; Niu, J.; Zhou, Y.; Wang, T.; Crittenden, J.C. Relative importance of humic and fulvic acid on ROS generation, dissolution, and toxicity of sulfide nanoparticles. Water Res. 2017, 124, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Mubeen, B.; Hasnain, A.; Wang, J.; Zheng, H.; Naqvi, S.A.H.; Prasad, R.; Rehman Au Sohail, M.A.; Hassan, M.Z.; Farhan, M.; Khan, M.A. Current Progress and Open Challenges for Combined Toxic Effects of Manufactured Nano-Sized Objects (MNO’s) on Soil Biota and Microbial Community. Coatings 2023, 13, 212. [Google Scholar] [CrossRef]
- Sabourian, P.; Yazdani, G.; Ashraf, S.S.; Frounchi, M.; Mashayekhan, S.; Kiani, S.; Kakkar, A. Effect of physico-chemical properties of nanoparticles on their intracellular uptake. Int. J. Mol. Sci. 2020, 21, 8019. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Jin, L.; Wang, X. Cadmium absorption and transportation pathways in plants. Int. J. Phytoremediat. 2017, 19, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Jin, M.; Yao, L.; He, B.; Ahmed, S.; Safdar, W.; Ahmad, I.; Cheng, D.B.; Lei, Z.; Sun, T. Physicochemical properties, pharmacokinetics, toxicology and application of nanocarriers. J. Mater. Chem. B 2023, 11, 716–733. [Google Scholar] [CrossRef] [PubMed]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of nanoparticle toxicity on their physical and chemical properties. Nanoscale Res. Lett. 2018, 13, 44. [Google Scholar] [CrossRef]
- Yusefi-Tanha, E.; Fallah, S.; Rostamnejadi, A.; Pokhrel, L.R. Root system architecture, copper uptake and tissue distribution in soybean (Glycine max L.) Merr.) grown in copper oxide nanoparticle (CuONP)-amended soil and implications for human nutrition. Plants 2020, 9, 1326. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kumar, A. Understanding the effect of the interaction of nanoparticles with roots on the uptake in plants. J. Environ. Nanotechnol. 2020, 3, 277–304. [Google Scholar] [CrossRef]
- Apodaca, S.A.; Cota-Ruiz, K.; Hernandez-Viezcas, J.A.; Gardea-Torresdey, J.L. Arbuscular mycorrhizal fungi alleviate phytotoxic effects of copper-based nanoparticles/compounds in spearmint (Mentha spicata). ACS Agric. Sci. Technol. 2022, 2, 661–670. [Google Scholar] [CrossRef]
- Arruda, S.C.C.; Silva, A.L.D.; Galazzi, R.M.; Azevedo, R.A.; Arruda, M.A.Z. Nanoparticles applied to plant science: A review. Talanta 2015, 131, 693–705. [Google Scholar] [CrossRef]
- Columbus, S.; Ramachandran, K.; Shameer, M.; Daoudi, K.; Gaidi, M. Antimicrobial nanosystems for environmental remediation applications. In Antimicrobial Nanosystems; Elsevier: Amsterdam, The Netherlands, 2023; pp. 417–435. [Google Scholar] [CrossRef]
- Ali, S.; Mehmood, A.; Khan, N. Uptake, translocation, and consequences of nanomaterials on plant growth and stress adaptation. J. Nanomater. 2021, 2021, 6677616. [Google Scholar] [CrossRef]
- Chou, L.Y.; Ming, K.; Chan, W.C. Strategies for the intracellular delivery of nanoparticles. Chem. Soc. Rev. 2011, 40, 233–245. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, M.S.; Susnik, E.; Drasler, B.; Taladriz-Blanco, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Understanding nanoparticle endocytosis to improve targeting strategies in nanomedicine. Chem. Soc. Rev. 2021, 50, 5397–5434. [Google Scholar] [CrossRef] [PubMed]
- Zangi, R.; Filella, M. Transport routes of metalloids into and out of the cell: A review of the current knowledge. Chem. Biol. Interact. 2012, 197, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.R.; Zia-ur-Rehman, M.; Sohail, M.I.; Haq, M.A.; Khalid, H.; Ayub, M.A.; Ishaq, G. Effects of rare earth oxide nanoparticles on plants. In Nanomaterials in Plants, Algae, and Microorganisms; Academic Press: Cambridge, MA, USA, 2018; pp. 239–275. [Google Scholar] [CrossRef]
- Singh, A.; Singh, N.B.; Hussain, I.; Singh, H.; Singh, S.C. Plant-nanoparticle interaction: An approach to improve agricultural practices and plant productivity. Int. J. Pharm. Sci. Invent. 2015, 4, 25–40. [Google Scholar]
- Park, J.H.; Gu, L.; Von Maltzahn, G.; Ruoslahti, E.; Bhatia, S.N.; Sailor, M.J. Biodegradable luminescent porous silicon nanoparticles for in vivo applications. Nat. Mater. 2009, 8, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liao, C.; Liu, S.; Xia, T.; Jiang, G. Nanotechnology: New opportunities for the development of patch-clamps. J. Nanobiotechnol. 2021, 19, 97. [Google Scholar] [CrossRef]
- Khodakovskaya, M.; Dervishi, E.; Mahmood, M.; Xu, Y.; Li, Z.; Watanabe, F.; Biris, A.S. Carbon nanotubes are able to penetrate plant seed coat and dramatically affect seed germination and plant growth. ACS Nano 2009, 3, 3221–3227. [Google Scholar] [CrossRef] [PubMed]
- Chandler, M.; Johnson, M.B.; Panigaj, M.; Afonin, K.A. Innate immune responses triggered by nucleic acids inspire the design of immunomodulatory nucleic acid nanoparticles (NANPs). In Therapeutic RNA Nanotechnology; Taylor and Francis: Abingdon, UK, 2021; pp. 1045–1060. [Google Scholar] [CrossRef]
- da Silva Santos, E.; Nogueira, K.A.B.; Fernandes, L.C.C.; Martins, J.R.P.; Reis, A.V.F.; Neto, J.D.B.V.; da Silva Júnior, I.J.; Pessoa, C.; Petrilli, R.; Eloy, J.O. EGFR targeting for cancer therapy: Pharmacology and immunoconjugates with drugs and nanoparticles. Int. J. Pharm. 2021, 592, 120082. [Google Scholar] [CrossRef]
- Gu, L.; Nawaz, T.; Qiu, Y.; Wu, Y.; Zhou, R. Photosynthetic conversion of CO2 and H2O to long-chain terpene alcohol by genetically engineered N2-fixing cyanobacteria. In Photosynthesis; Academic Press: Cambridge, MA, USA, 2023; pp. 451–461. [Google Scholar] [CrossRef]
- Flores, E.; Herrero, A. Compartmentalized function through cell differentiation in filamentous cyanobacteria. Nat. Rev. Microbiol. 2010, 8, 39–50. [Google Scholar] [CrossRef]
- Kieninger, A.K.; Maldener, I. Cell–cell communication through septal junctions in filamentous cyanobacteria. Curr. Opin. Microbiol. 2021, 61, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M.; Brown, A.C. Bacterial outer membrane vesicles as antibiotic delivery vehicles. Front. Immunol. 2021, 12, 733064. [Google Scholar] [CrossRef] [PubMed]
- Biller, S.J.; McDaniel, L.D.; Breitbart, M.; Rogers, E.; Paul, J.H.; Chisholm, S.W. Membrane vesicles in sea water: Heterogeneous DNA content and implications for viral abundance estimates. ISME J. 2017, 11, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Naz, T.; Iqbal, M.M.; Fahad, S.; Akhtar, J.; Saqib, M.; Alamri, S.; Siddiqui, M.H.; Saud, S.; Khattak, J.Z.K.; Ali, S.; et al. Bio-fortification of Two Wheat Cultivars with Iron and Zinc Through Their Soil and Foliar Application in Salt-Factored Soil: Growth, Ionic, Physiological, and Biochemical Modifications. J. Plant Growth Regul. 2023, 42, 5727–5745. [Google Scholar] [CrossRef]
- Lea-Smith, D.J.; Ross, N.; Zori, M.; Bendall, D.S.; Dennis, J.S.; Scott, S.A.; Smith, A.G.; Howe, C.J. Thylakoid terminal oxidases are essential for the cyanobacterium Synechocystis sp. PCC 6803 to survive rapidly changing light intensities. Plant Physiol. 2013, 162, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Patterson-Fortin, L.M. LexA-Mediated Repression of the Cyanobacterial RNA Helicase, crhR; University of Alberta: Edmonton, AL, Canada, 2008. [Google Scholar] [CrossRef]
- Rippka, R.; Waterbury, J.B.; Stanier, R.Y. Provisional generic assignments for cyanobacteria in pure culture. In The Prokaryotes: A Handbook on Habitats, Isolation, and Identification of Bacteria; Springer: Berlin/Heidelberg, Germany, 1981; pp. 247–256. [Google Scholar] [CrossRef]
- Houle, D.; Bilodeau Gauthier, S.; Paquet, S.; Planas, D.; Warren, A. Identification of two genera of N2-fixing cyanobacteria growing on three feather moss species in boreal forests of Quebec, Canada. Botany 2006, 84, 1025–1029. [Google Scholar] [CrossRef]
- Meeks, J.C. Physiological adaptations in nitrogen-fixing Nostoc–plant symbiotic associations. In Prokaryotic Symbionts in Plants; Springer: Berlin/Heidelberg, Germany, 2007; pp. 181–205. [Google Scholar] [CrossRef]
- Arróniz-Crespo, M.; Pérez-Ortega, S.; De Los Ríos, A.; Green, T.G.; Ochoa-Hueso, R.; Casermeiro, M.Á.; de la Cruz, M.T.; Pintado, A.; Palacios, D.; Rozzi, R.; et al. Bryophyte-cyanobacteria associations during primary succession in recently Deglaciated areas of Tierra del Fuego (Chile). PLoS ONE 2014, 9, e96081. [Google Scholar] [CrossRef]
- Warshan, D.; Liaimer, A.; Pederson, E.; Kim, S.Y.; Shapiro, N.; Woyke, T.; Altermark, B.; Pawlowski, K.; Weyman, P.D.; Dupont, C.L.; et al. Genomic changes associated with the evolutionary transitions of Nostoc to a plant symbiont. Mol. Biol. Evol. 2018, 35, 1160–1175. [Google Scholar] [CrossRef]
- West, N.J.; Adams, D.G. Phenotypic and genotypic comparison of symbiotic and free-living cyanobacteria from a single field site. Appl. Environ. Microbiol. 1997, 63, 4479–4484. [Google Scholar] [CrossRef]
- Gentili, F.; Nilsson, M.C.; Zackrisson, O.; DeLuca, T.H.; Sellstedt, A. Physiological and molecular diversity of feather moss associative N2-fixing cyanobacteria. J. Exp. Bot. 2005, 56, 3121–3127. [Google Scholar] [CrossRef]
- Shakoor, N.; Hussain, M.; Adeel, M.; Azeem, I.; Ahmad, M.A.; Zain, M.; Zhang, P.; Li, Y.; Quanlong, W.; Horton, R.; et al. Lithium-induced alterations in soybean nodulation and nitrogen fixation through multifunctional mechanisms. Sci. Total Environ. 2023, 904, 166438. [Google Scholar] [CrossRef] [PubMed]
- Rousk, K.; Jones, D.L.; Deluca, T.H. Moss-cyanobacteria associations as biogenic sources of nitrogen in boreal forest ecosystems. Front. Microbiol. 2013, 4, 150. [Google Scholar] [CrossRef] [PubMed]
- van den Elzen, E.; Bengtsson, F.; Fritz, C.; Rydin, H.; Lamers, L.P. Variation in symbiotic N2 fixation rates among Sphagnum mosses. PLoS ONE 2020, 15, e0228383. [Google Scholar] [CrossRef] [PubMed]
- Sorkin, M.L.; Nusinow, D.A. Time will tell: Intercellular communication in the plant clock. Trends Plant Sci. 2021, 26, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Regot, S.; Macia, J.; Conde, N.; Furukawa, K.; Kjellén, J.; Peeters, T.; Hohmann, S.; De Nadal, E.; Posas, F.; Solé, R. Distributed biological computation with multicellular engineered networks. Nature 2011, 469, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Habibullah, G.; Viktorova, J.; Ruml, T. Current strategies for noble metal nanoparticle synthesis. Nanoscale Res. Lett. 2021, 16, 47. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.; Prakash, R.; Rai, M.P. Latest advances and status analysis of nanomaterials for microalgae photosystem, lipids and biodiesel: A state of art. J. Environ. Chem. Eng. 2022, 11, 109111. [Google Scholar] [CrossRef]
- Scharwies, J.D.; Dinneny, J.R. Water transport, perception, and response in plants. J. Plant Res. 2019, 132, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin, self-organisation, and the colonial nature of plants. Curr. Biol. 2011, 21, R331–R337. [Google Scholar] [CrossRef]
- Singh, S.; Bhoi, T.K.; Vyas, V. Interceding Microbial Biofertilizers in Agroforestry System for Enhancing Productivity. In Plant Growth Promoting Microorganisms of Arid Region; Springer: Singapore, 2023; pp. 161–183. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.K.; Zhou, Y.; Wang, J.; Jiang, Y.; Shen, N.; Wang, Y.; Yang, L.; Jiang, M. Unlocking the strength of plant growth promoting Pseudomonas in improving crop productivity in normal and challenging environments: A review. J. Plant Interact. 2022, 17, 220–238. [Google Scholar] [CrossRef]
- Abdellatif, A.A.; Khan, R.A.; Alhowail, A.H.; Alqasoumi, A.; Sajid, S.M.; Mohammed, A.M.; Alsharidah, M.; Al Rugaie, O.; Mousa, A.M. Octreotide-conjugated silver nanoparticles for active targeting of somatostatin receptors and their application in a nebulized rat model. Nanotechnol. Rev. 2021, 11, 266–283. [Google Scholar] [CrossRef]
- Bouallegui, Y.; Ben Younes, R.; Oueslati, R.; Sheehan, D. Redox proteomic insights into involvement of clathrin-mediated endocytosis in silver nanoparticles toxicity to Mytilus galloprovincialis. PLoS ONE 2018, 13, e0205765. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.K.; Lyon, D.Y.; Alvarez, P.J. Comparative eco-toxicity of nanoscale TiO2, SiO2, and ZnO water suspensions. Water Res. 2006, 40, 3527–3532. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, A.; Costa, B.; Ferreira, M.V.; Miguéis, D.; Louros, J.M.S.; Durazzo, A.; Lucarini, M.; Eder, P.; Chaud, M.V.; Morsink, M.; et al. Nanotoxicology and Nanosafety: Safety-By-Design and Testing at a Glance. Int. J. Environ. Res. Public Health 2020, 17, 4657. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Savage, D.T.; Hilt, J.Z.; Dziubla, T.D. In Vitro Methods for Assessing Nanoparticle Toxicity. Methods Mol. Biol. 2019, 1894, 1–29. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Drasler, B.; Sayre, P.; Steinhäuser, K.G.; Petri-Fink, A.; Rothen-Rutishauser, B. In vitro approaches to assess the hazard of nanomaterials. NanoImpact 2017, 8, 99–116. [Google Scholar] [CrossRef]
- Li, J.; Hu, C.; Liu, B.; Liu, Z. Dual pathway reduction of Mo4+ and photogenerated electrons restore catalytic sites to enhance heterogeneous peroxymonosulfate activation system. Chem. Eng. J. 2023, 452, 139246, ISSN 1385-8947. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, H.; Liang, C.; Duan, L. Novel technology for synergistic SO2 reduction during the carbonation process via a CaO-Char mixed system. Chem. Eng. J. 2024, 488, 150678, ISSN 1385-8947. [Google Scholar] [CrossRef]
- Chen, X.; Lu, R.; Liu, P.; Li, X. Effects of nano-TiO2 on Chlamydomonas reinhardtii cell surface under UV, natural light conditions. J. Wuhan Univ. Technol. Mater. Sci. Ed. 2017, 32, 217–222. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Maiorano, G.; Sabella, S.; Sorce, B.; Brunetti, V.; Malvindi, M.A.; Cingolani, R.; Pompa, P.P. Effects of cell culture media on the dynamic formation of protein-nanoparticle complexes and influence on the cellular response. ACS Nano 2010, 47, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chang, Y.; Chen, Y. Toxicity and bioaccumulation of TiO2 nanoparticle aggregates in Daphnia magna. Chemosphere 2010, 78, 209–215. [Google Scholar] [CrossRef]
- Wang, H.; Wick, R.L.; Xing, B. Toxicity of nanoparticulate and bulk ZnO, Al2O3 and TiO2 to the nematode Caenorhabditis elegans. Environ. Pollut. 2009, 157, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Long, T.C.; Saleh, N.; Tilton, R.D.; Lowry, G.V.; Veronesi, B. Titanium dioxide (P25) produces reactive oxygen species in immortalized brain microglia (BV2): Implications for nanoparticle neurotoxicity. Environ. Sci. Technol. 2006, 40, 4346–4352. [Google Scholar] [CrossRef]
- Lynch, I.; Cedervall, T.; Lundqvist, M.; Cabaleiro-Lago, C.; Linse, S.; Dawson, K.A. The nanoparticle–protein complex as a biological entity; a complex fluids and surface science challenge for the 21st century. Adv. Colloid Interface Sci. 2007, 134, 167–174. [Google Scholar] [CrossRef]
- Hillegass, J.M.; Shukla, A.; Lathrop, S.A.; MacPherson, M.B.; Fukagawa, N.K.; Mossman, B.T. Assessing nanotoxicity in cells in vitro. Wiley Interdisciplinary Reviews: Nanomed. Nanobiotechnology 2010, 2, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Simon-Deckers, A.; Loo, S.; Mayne-L’hermite, M.; Herlin-Boime, N.; Menguy, N.; Reynaud, C.; Gouget, B.; Carrière, M. Size, composition and shape dependent toxicological impact of metal oxide nanoparticles and carbon nanotubes toward bacteria. Environ. Sci. Technol. 2009, 43, 8423–8429. [Google Scholar] [CrossRef] [PubMed]
- Servin, A.; Elmer, W.; Mukherjee, A.; De la Torre-Roche, R.; Hamdi, H.; White, J.C.; Bindraban, P.; Dimkpa, C. A review of the use of engineered nanomaterials to suppress plant disease and enhance crop yield. J. Nanopart. Res. 2015, 17, 92. [Google Scholar] [CrossRef]
- Azameti, M.K.; Imoro, A.W. Nanotechnology: A promising field in enhancing abiotic stress tolerance in plants. Crop Des. 2023, 28, 100037. [Google Scholar] [CrossRef]
- Allouzi, M.M.; Tang, D.Y.; Chew, K.W.; Rinklebe, J.; Bolan, N.; Allouzi, S.M.; Show, P.L. Micro (nano) plastic pollution: The ecological influence on soil-plant system and human health. Sci. Total Environ. 2021, 20, 147815. [Google Scholar] [CrossRef]
- Karlsson, H.L.; Cronholm, P.; Gustafsson, J.; Moller, L. Copper oxide nanoparticles are highly toxic: A comparison between metal oxide nanoparticles and carbon nanotubes. Chem. Res. Toxicol. 2008, 21, 1726–1732. [Google Scholar] [CrossRef]
- Jia, G.; Wang, H.; Yan, L.; Wang, X.; Pei, R.; Yan, T.; Zhao, Y.; Guo, X. Cytotoxicity of carbon nanomaterials: Single-wall nanotube, multi-wall nanotube, and fullerene. Environ. Sci. Technol. 2005, 39, 1378–1383. [Google Scholar] [CrossRef]
- Luanpitpong, S.; Wang, L.; Rojanasakul, Y. The effects of carbon nanotubes on lung and dermal cellular behaviors. Nanomedicine 2014, 9, 895–912. [Google Scholar] [CrossRef] [PubMed]
- Simon-Deckers, A.; Gouget, B.; Mayne-L’Hermite, M.; Herlin-Boime, N.; Reynaud, C.; Carriere, M. In vitro investigation of oxide nanoparticle and carbon nanotube toxicity and intracellular accumulation in A549 human pneumocytes. Toxicology 2008, 253, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Vinson, D.; Abt, D.; Hurt, R.H.; Rand, D.M. Differential toxicity of carbon nanomaterials in Drosophila: Larval dietary uptake is benign, but adult exposure causes locomotor impairment and mortality. Environ. Sci. Technol. 2009, 43, 6357–6363. [Google Scholar] [CrossRef]
- Petersen, E.J.; Henry, T.B.; Zhao, J.; MacCuspie, R.I.; Kirschling, T.L.; Dobrovolskaia, M.A.; Hackley, V.; Xing, B.; White, J.C. Identification and avoidance of potential artifacts and misinterpretations in nanomaterial ecotoxicity measurements. Environ. Sci. Technol. 2014, 48, 4226–4246. [Google Scholar] [CrossRef] [PubMed]
- Lyon, D.Y.; Brunet, L.; Hinkal, G.W.; Wiesner, M.R.; Alvarez, P.J. Antibacterial activity of fullerene water suspensions (nC60) is not due to ROS-mediated damage. Nano Lett. 2008, 8, 1539–1543. [Google Scholar] [CrossRef]
- Liu, J.; Aruguete, D.M.; Murayama, M.; Hochella, M.F., Jr. Influence of size and aggregation on the reactivity of an environmentally and industrially relevant nanomaterial (PbS). Environ. Sci. Technol. 2009, 43, 8178–8183. [Google Scholar] [CrossRef]
- Carvalho, L.Â.; Oya-Silva, L.F.; Perussolo, M.C.; Guaita, G.O.; Brito, J.C.; Evans, A.A.; Prodocimo, M.M.; Cestari, M.M.; Braga, T.T.; de Assis, H.C. Experimentally exposed toxic effects of long-term exposure to environmentally relevant concentrations of CIP in males and females of the silver catfish Rhamdia quelen. Chemosphere 2023, 336, 139216. [Google Scholar] [CrossRef]
- Kent, R.D.; Vikesland, P.J. Dissolution and persistence of copper-based nanomaterials in undersaturated solutions with respect to cupric solid phases. Environ. Sci. Technol. 2016, 50, 6772–6781. [Google Scholar] [CrossRef]
- Schmidt, S.N.; Mayer, P. Linking algal growth inhibition to chemical activity: Baseline toxicity required 1% of saturation. Chemosphere 2015, 120, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.M.; Feris, K.; Bell, J.; Wingett, D.G.; Hanley, C.; Punnoose, A. Selective toxicity of zinc oxide nanoparticles to prokaryotic and eukaryotic systems. Appl. Phys. Lett. 2007, 90, 213902. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.S.; Koh, D.C.; Kong, I.C. Toxicity evaluation of individual and mixtures of nanoparticles based on algal chlorophyll content and cell count. Materials 2018, 11, 121. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Kovochich, M.; Liong, M.; Madler, L.; Gilbert, B.; Shi, H.; Yeh, J.I.; Zink, J.I.; Nel, A.E. Comparison of the mechanism of toxicity of zinc oxide and cerium oxide nanoparticles based on dissolution and oxidative stress properties. ACS Nano 2008, 2, 2121–2134. [Google Scholar] [CrossRef] [PubMed]
- Blinova, I.; Ivask, A.; Heinlaan, M.; Mortimer, M.; Kahru, A. Ecotoxicity of nanoparticles of CuO and ZnO in natural water. Environ. Pollut. 2010, 158, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Thill, A.; Zeyons, O.; Spalla, O.; Chauvat, F.; Rose, J.; Auffan, M.; Flank, A.M. Cytotoxicity of CeO2 nanoparticles for Escherichia coli. Physico-chemical insight of the cytotoxicity mechanism. Sci. Total Environ. 2006, 40, 6151–6156. [Google Scholar] [CrossRef] [PubMed]
- Keenan, C.R.; Goth-Goldstein, R.; Lucas, D.; Sedlak, D.L. Oxidative stress induced by zero-valent iron nanoparticles and Fe (II) in human bronchial epithelial cells. Environ. Sci. Technol. 2009, 43, 4555–4560. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Mashayekhi, H.; Xing, B. Bacterial toxicity comparison between nano-and micro-scaled oxide particles. Environ. Pollut. 2009, 157, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.M.; Hess, K.L.; Gearhart, J.M.; Geiss, K.T.; Schlager, J.J. In vitro toxicity of nanoparticles in BRL 3A rat liver cells. Toxicol. In Vitro 2005, 19, 975–983. [Google Scholar] [CrossRef]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-dependent cytotoxicity of gold nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the gram-negative bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of silver nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef]
- Kawata, K.; Osawa, M.; Okabe, S. In vitro toxicity of silver nanoparticles at noncytotoxic doses to HepG2 human hepatoma cells. Environ. Sci. Technol. 2009, 43, 6046–6051. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Greden, K.; Alvarez, P.J.; Gregory, K.B.; Lowry, G.V. Adsorbed polymer and NOM limits adhesion and toxicity of nano scale zerovalent iron to E. coli. Environ. Sci. Technol. 2010, 44, 3462–3467. [Google Scholar] [CrossRef]
- Stebounova, L.V.; Adamcakova-Dodd, A.; Kim, J.S.; Park, H.; O’Shaughnessy, P.T.; Grassian, V.H.; Thorne, P.S. Nanosilver induces minimal lung toxicity or inflammation in a subacute murine inhalation model. Part. Fibre Toxicol. 2011, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.Y.; Byeon, J.H.; Park, J.H.; Hwang, J. Susceptibility constants of Escherichia coli and Bacillus subtilis to silver and copper nanoparticles. Sci. Total Environ. 2007, 373, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Fujioka, K.; Oku, T.; Suga, M.; Sasaki, Y.F.; Ohta, T.; Yasuhara, M.; Suzuki, K.; Yamamoto, K. Physicochemical properties and cellular toxicity of nanocrystal quantum dots depend on their surface modification. Nano Lett. 2004, 4, 2163–2169. [Google Scholar] [CrossRef]
- Hussain, S.M.; Javorina, A.K.; Schrand, A.M.; Duhart, H.M.; Ali, S.F.; Schlager, J.J. The interaction of manganese nanoparticles with PC-12 cells induces dopamine depletion. Toxicol. Sci. 2006, 92, 456–463. [Google Scholar] [CrossRef]
- Jiang, W.; Kim, B.Y.; Rutka, J.T.; Chan, W.C. Nanoparticle-mediated cellular response is size-dependent. Nat. Nanotechnol. 2008, 3, 145–150. [Google Scholar] [CrossRef]
- Braydich-Stolle, L.K.; Speshock, J.L.; Castle, A.; Smith, M.; Murdock, R.C.; Hussain, S.M. Nanosized aluminum altered immune function. ACS Nano 2010, 4, 3661–3670. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, L.; Liang, C.; Wang, C.; Peng, R.; Liu, Z. Photothermal therapy with immune-adjuvant nanoparticles together with checkpoint blockade for effective cancer immunotherapy. Nat. Commun. 2016, 7, 13193. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef]
- De La Torre-Roche, R.; Hawthorne, J.; Deng, Y.; Xing, B.; Cai, W.; Newman, L.A.; Wang, Q.; Ma, X.; Hamdi, H.; White, J.C. Multiwalled carbon nanotubes and C60 fullerenes differentially impact the accumulation of weathered pesticides in four agricultural plants. Environ. Sci. Technol. 2013, 47, 12539–12547. [Google Scholar] [CrossRef] [PubMed]
- Schwab, F.; Zhai, G.; Kern, M.; Turner, A.; Schnoor, J.L.; Wiesner, M.R. Barriers pathways and processes for uptake, translocation and accumulation of nanomaterials in plants–Critical review. Nanotoxicology 2016, 10, 257–278. [Google Scholar] [CrossRef]
- Sanati, M.; Afshari, A.R.; Kesharwani, P.; Sukhorukov, V.N.; Sahebkar, A. Recent trends in the application of nanoparticles in cancer therapy: The involvement of oxidative stress. J. Control. Release 2022, 348, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Adaptive response to oxidative stress: Bacteria, fungi, plants and animals. Comp. Biochem. Physiol. Part C Toxicol. Appl. Pharmacol. 2011, 153, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Krumova, K.; Cosa, G. Overview of Reactive Oxygen Species; The Royal Society of Chemistry: London, UK, 2016; pp. 1–21. [Google Scholar] [CrossRef]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Soares, C.; Sousa, B.; Martins, M.; Kumar, V.; Shahzad, B.; Sidhu, G.P.; Bali, A.S.; Asgher, M.; Bhardwaj, R.; et al. Nitric oxide-mediated regulation of oxidative stress in plants under metal stress: A review on molecular and biochemical aspects. Physiol. Plant. 2020, 168, 318–344. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Clague, L.A. Comparative Genomics and Transcriptomics of Steinernema carpocapsae and Caenorhabditis elegans; University of California: Irvine, CA, USA, 2020. [Google Scholar]
- Nawaz, T.; Gu, L.; Fahad, S.; Saud, S.; Jiang, Z.; Hassan, S.; Harrison, M.Y.; Liu, K.; Khan, M.A.; Liu, K.; et al. A comprehensive review of the therapeutic potential of cyanobacterial marine bioactives: Unveiling the hidden treasures of the sea. Food Energy Secur. 2023, 12, e495. [Google Scholar] [CrossRef]
- Vurro, M.; Miguel-Rojas, C.; Pérez-de-Luque, A. Safe nanotechnologies for increasing the effectiveness of environmentally friendly natural agrochemicals. Pest Manag. Sci. 2019, 75, 2403–2412. [Google Scholar] [CrossRef] [PubMed]
- Mani, P.K.; Mondal, S. Agri-nanotechniques for plant availability of nutrients. In Plant Nanotechnology: Principles and Practices; Springer: Berlin/Heidelberg, Germany, 2016; pp. 263–303. [Google Scholar] [CrossRef]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef] [PubMed]
- Shiohara, A.; Hoshino, A.; Hanaki, K.I.; Suzuki, K.; Yamamoto, K. On the cyto-toxicity caused by quantum dots. Microbiol. Immunol. 2004, 48, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Bao, M.; Wang, B.; Wu, S.; Luo, L.; Li, B.; Lin, J. Inhibitory effects of Cu2O/SiO2 on the growth of Microcystis aeruginosa and its mechanism. Nanomaterials 2019, 9, 1669. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, I.R.; Olsson, A.L.; Tufenkji, N. Deposition kinetics of quantum dots and polystyrene latex nanoparticles onto alumina: Role of water chemistry and particle coating. Environ. Sci. Technol. 2013, 47, 2212–2220. [Google Scholar] [CrossRef] [PubMed]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. 2011, 75, 583–609. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Basir, A.; Adnan, M.; Fahad, S.; Ali, J.; Mussart, M.; Mian, I.A.; Ahmad, M.; Saleem, M.H.; Naseem, W.; et al. Biochar to Improve Crops Yield and Quality Under a Changing Climate. In Sustainable Agriculture Reviews 61: Biochar to Improve Crop Production and Decrease Plant Stress under a Changing Climate; Springer International Publishing: Cham, Switzerland, 2023; pp. 57–73. [Google Scholar] [CrossRef]
- Bityutsky, V.S.; Tsekhmistrenko, S.I.; Tsekhmistrenko, O.S.; Tymoshok, N.O.; Spivak, M.Y. Regulation of redox processes in biological systems with the participation of the Keap1/Nrf2/ARE signaling pathway, biogenic selenium nanoparticles as Nrf2 activators. Regul. Mech. Biosyst. 2020, 11, 483–493. [Google Scholar] [CrossRef]
- Abideen, Z.; Hanif, M.; Munir, N.; Nielsen, B.L. Impact of nanomaterials on the regulation of gene expression and metabolomics of plants under salt stress. Plants 2022, 11, 691. [Google Scholar] [CrossRef]
- Wu, N.; Li, Z.; Wu, F.; Tang, M. Comparative photochemistry activity and antioxidant responses in male and female Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Sci. Rep. 2016, 6, 37663. [Google Scholar] [CrossRef]
- Atkinson, J.T. Controlling Bioenergetic Systems Using Protein Design and Synthetic Biology. Ph.D. Thesis, Rice University, Houston, TX, USA, 2019. [Google Scholar]
- Shah, S.; Chen, C.; Sun, Y.; Wang, D.; Nawaz, T.; El-Kahtany, K.; Fahad, S. Mechanisms of nitric oxide involvement in plant-microbe interaction and its enhancement of stress resistance. Plant Stress 2023, 2, 100191. [Google Scholar] [CrossRef]
- Romanowska-Duda, Z.; Szufa, S.; Grzesik, M.; Piotrowski, K.; Janas, R. The promotive effect of cyanobacteria and Chlorella sp. foliar biofertilization on growth and metabolic activities of willow (Salix viminalis L.) plants as feedstock production, solid biofuel and biochar as C carrier for fertilizers via torrefaction process. Energies 2021, 14, 5262. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Los, D.A.; Schmitt, F.J.; Zharmukhamedov, S.K.; Kuznetsov, V.V.; Allakhverdiev, S.I. The impact of the phytochromes on photosynthetic processes. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 400–408. [Google Scholar] [CrossRef]
- Lin, W.R.; Tan, S.I.; Hsiang, C.C.; Sung, P.K.; Ng, I.S. Challenges and opportunity of recent genome editing and multi-omics in cyanobacteria and microalgae for biorefinery. Bioresour. Technol. 2019, 291, 121932. [Google Scholar] [CrossRef]
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant biostimulants from cyanobacteria: An emerging strategy to improve yields and sustainability in agriculture. Plants 2021, 10, 643. [Google Scholar] [CrossRef]
- Wang, W.; Liu, X.; Lu, X. Engineering cyanobacteria to improve photosynthetic production of alka (e) nes. Biotechnol. Biofuels 2013, 6, 69. [Google Scholar] [CrossRef]
- Bombar, D.; Heller, P.; Sanchez-Baracaldo, P.; Carter, B.J.; Zehr, J.P. Comparative genomics reveals surprising divergence of two closely related strains of uncultivated UCYN-A cyanobacteria. ISME J. 2014, 8, 2530–2542. [Google Scholar] [CrossRef]
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Berta, G.; Doussan, C.; Merchan, F.; Crespi, M. Plant Root Growth, Architecture and Function; Springer: Berlin/Heidelberg, Germany, 2009; pp. 153–187. [Google Scholar] [CrossRef]
- Derakhshan Nejad, Z.; Jung, M.C.; Kim, K.H. Remediation of soils contaminated with heavy metals with an emphasis on immobilization technology. Environ. Geochem. Health 2018, 40, 927–953. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Chen, H.; Sarsaiya, S.; Qin, S.; Liu, H.; Awasthi, M.K.; Kumar, S.; Singh, L.; Zhang, Z.; Bolan, N.S.; et al. Current research trends on micro-and nano-plastics as an emerging threat to global environment: A review. Hazard. Mater. 2021, 409, 124967. [Google Scholar] [CrossRef]
- Chouhan, G.K.; Verma, J.P.; Jaiswal, D.K.; Mukherjee, A.; Singh, S.; de Araujo Pereira, A.P.; Liu, H.; Abd_Allah, E.F.; Singh, B.K. Phytomicrobiome for promoting sustainable agriculture and food security: Opportunities, challenges, and solutions. Microbiol. Res. 2021, 248, 126763. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. New roots for agriculture: Exploiting the root phenome. Philos. Trans. R. Soc. B 2012, 367, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Shenton, M.; Iwamoto, C.; Kurata, N.; Ikeo, K. Effect of wild and cultivated rice genotypes on rhizosphere bacterial community composition. Rice 2016, 9, 42. [Google Scholar] [CrossRef]
- Szoboszlay, M.; Lambers, J.; Chappell, J.; Kupper, J.V.; Moe, L.A.; McNear, D.H., Jr. Comparison of root system architecture and rhizosphere microbial communities of Balsas teosinte and domesticated corn cultivars. Soil Biol. Biochem. 2015, 80, 34–44. [Google Scholar] [CrossRef]
- Schmidt, J.E.; Mazza Rodrigues, J.L.; Brisson, V.L.; Kent, A.; Gaudin, A.C.M. Impacts of directed evolution and soil management legacy on the maize rhizobiome. Soil Biol. Biochem. 2020, 145, 107794. [Google Scholar] [CrossRef]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Corneo, P.E.; Suenaga, H.; Kertesz, M.A.; Dijkstra, F.A. Effect of twenty-four wheat genotypes on soil biochemical and microbial properties. Plant Soil 2016, 404, 141–155. [Google Scholar] [CrossRef]
- Robertson-Albertyn, S.; Alegria Terrazas, R.; Balbirnie, K.; Blank, M.; Janiak, A.; Szarejko, I.; Chmielewska, B.; Karcz, J.; Morris, J.; Hedley, P.E.; et al. Root hair mutations displace the barley rhizosphere microbiota. Front. Plant Sci. 2017, 8, 1094. [Google Scholar] [CrossRef]
- Pérez-Jaramillo, J.E.; Carrión, V.J.; Bosse, M.; Ferrão, L.F.V.; de Hollander, M.; Garcia, A.A.F.; Ramírez, C.A.; Mendes, R.; Raaijmakers, J.M. Linking rhizosphere microbiome composition of wild and domesticated Phaseolus vulgaris to genotypic and root phenotypic traits. ISME J. 2017, 11, 2244–2257. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Minkina, T.; Kumari, A.; Shende, S.S.; Ranjan, A.; Faizan, M.; Barakvov, A.; Gromovik, A.; Gorbunova, N.; Rajput, P.; et al. A review on nanobioremediation approaches for restoration of contaminated soil. Eurasian J. Soil Sci. 2022, 11, 43–60. [Google Scholar] [CrossRef]
- Sharma, N.; Kumari, R.M.; Gupta, N.; Syed, A.; Bahkali, A.H.; Nimesh, S. Poly-(lactic-co-glycolic) acid nanoparticles for synergistic delivery of epirubicin and paclitaxel to human lung cancer cells. Molecules 2020, 25, 4243. [Google Scholar] [CrossRef]
- Zheng, Y.; Sun, Z.; Liu, Y.; Cao, T.; Zhang, H.; Hao, M.; Chen, R.; Dzakpasu, M.; Wang, X.C. Phytoremediation mechanisms and plant eco-physiological response to microorganic contaminants in integrated vertical-flow constructed wetlands. J. Hazard. Mater. 2022, 424, 127611. [Google Scholar] [CrossRef] [PubMed]
- Rather, M.A.; Bhuyan, S.; Chowdhury, R.; Sarma, R.; Roy, S.; Neog, P.R. Nanoremediation strategies to address environmental problems. Sci. Total Environ. 2023, 10, 163998. [Google Scholar] [CrossRef] [PubMed]
- Salam, A.; Afridi, M.S.; Javed, M.A.; Saleem, A.; Hafeez, A.; Khan, A.R.; Zeeshan, M.; Ali, B.; Azhar, W.; Sumaira; et al. Nano-priming against abiotic stress: A way forward towards sustainable agriculture. Sustainability 2022, 14, 14880. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C. ZnO nanoparticle biosynthesis and its effect on phosphorous-mobilizing enzyme secretion and gum contents in Clusterbean (Cyamopsis tetragonoloba L.). Agric. Res. 2013, 2, 48–57. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A. Multifunctional hybrid nanomaterials for sustainable agri-food and ecosystems: A note from the editor. In Multifunctional Hybrid Nanomaterials for Sustainable Agri-Food and Ecosystems; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–19. [Google Scholar] [CrossRef]
- Li, M.; Sun, X.; Yin, M.; Shen, J.; Yan, S. Recent advances in nanoparticle-mediated co-delivery system: A promising strategy in medical and Agricultural Field. Int. J. Mol. Sci. 2023, 24, 5121. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Golińska, P. Myconanotechnology Emerging Trends and Applications; Routledge: London, UK, 2023. [Google Scholar]
- Agarwal, P.; Soni, R.; Kaur, P.; Madan, A.; Mishra, R.; Pandey, J.; Singh, S.; Singh, G. Cyanobacteria as a promising alternative for sustainable environment: Synthesis of biofuel and biodegradable plastics. Front. Microbiol. 2022, 13, 939347. [Google Scholar] [CrossRef] [PubMed]
- Kothari, R.; Ahmad, S.; Pathak, V.V.; Pandey, A.; Kumar, A.; Shankarayan, R.; Black, P.N.; Tyagi, V.V. Algal-based biofuel generation through flue gas and wastewater utilization: A sustainable prospective approach. Biomass Convers. Biorefin. 2021, 11, 1419–1442. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Singh, M. Cyanobacteria: A sustainable and commercial bio-resource in production of bio-fertilizer and bio-fuel from waste waters. Environ. Sustain. 2019, 3, 100008. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef]
- Cao, T.N.; Mukhtar, H.; Le, L.T.; Tran, D.P.; Ngo, M.T.; Nguyen, T.B.; Bui, X.T. Roles of microalgae-based biofertilizer in sustainability of green agriculture and food-water-energy security nexus. Sci. Total Environ. 2023, 870, 161927. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Rajakumar, G.; Chung, I.M. Nanotechnology: Current uses and future applications in the food industry. 3 Biotech 2018, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Litter, M.I.; Ahmad, A. (Eds.) Industrial Applications of Nanoparticles; CRC Press: Boca Raton, FL, USA, 2023. [Google Scholar]
- Hamida, R.S.; Ali, M.A.; Redhwan, A.; Bin-Meferij, M.M. Cyanobacteria—A promising platform in green nanotechnology: A review on nanoparticles fabrication and their prospective applications. Int. J. Nanomed. 2020, 13, 6033–6066. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Wang, Y.; Qin, Y.X. Promoting neuroregeneration by applying dynamic magnetic fields to a novel nanomedicine: Superparamagnetic iron oxide (SPIO)-gold nanoparticles bounded with nerve growth factor (NGF). Nanomed. Nanotechnol. Biol. 2018, 14, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, T.; Costantini, M.; Pal, S.; Kumar, A. Oxygenation therapies for improved wound healing: Current trends and technologies. J. Mater. Chem. B. 2022, 10, 7905–7923. [Google Scholar] [CrossRef] [PubMed]
- Smith, O.M.; Cohen, A.L.; Reganold, J.P.; Jones, M.S.; Orpet, R.J.; Taylor, J.M.; Thurman, J.H.; Cornell, K.A.; Olsson, R.L.; Ge, Y.; et al. Landscape context affects the sustainability of organic farming systems. Proc. Natl. Acad. Sci. USA 2020, 117, 2870–2878. [Google Scholar] [CrossRef]
- Rosenhauer, A.M.; Beach, L.Q.; Jeffress, E.C.; Thompson, B.M.; McCann, K.E.; Partrick, K.A.; Diaz, B.; Norvelle, A.; Choi, D.C.; Huhman, K.L. Brain-derived neurotrophic factor signaling mitigates the impact of acute social stress. Neuropharmacology 2019, 148, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Hassanpour Adeh, E.; Selker, J.S.; Higgins, C.W. Remarkable agrivoltaic influence on soil moisture, micrometeorology and water-use efficiency. PLoS ONE 2018, 13, e0203256. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Raliya, R.; Biswas, P.; Tarafdar, J.C. TiO2 nanoparticle biosynthesis and its physiological effect on mung bean (Vigna radiata L.). Biotechnol. Rep. 2015, 5, 22–26. [Google Scholar] [CrossRef]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.S.; Saxena, A.K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatal. Agric. Biotechnol. 2020, 23, 101487. [Google Scholar] [CrossRef]
- Raimi, A.; Adeleke, R.; Roopnarain, A. Soil fertility challenges and Biofertiliser as a viable alternative for increasing smallholder farmer crop productivity in sub-Saharan Africa. Cogent Food Agric. 2017, 3, 1400933. [Google Scholar] [CrossRef]
- Pahalvi, H.N.; Rafiya, L.; Rashid, S.; Nisar, B.; Kamili, A.N. Chemical fertilizers and their impact on soil health. Microbiota and Biofertilizers, Volume 2: Ecofriendly Tools for Reclamation of Degraded Soil Environs; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–20. [Google Scholar] [CrossRef]
- Rajput, V.D.; Kumari, A.; Upadhyay, S.K.; Minkina, T.; Mandzhieva, S.; Ranjan, A.; Sushkova, S.; Burachevskaya, M.; Rajput, P.; Konstantinova, E.; et al. Can nanomaterials improve the soil microbiome and crop productivity? Agriculture 2023, 13, 231. [Google Scholar] [CrossRef]
- Gebbers, R.; Adamchuk, V.I. Precision agriculture and food security. Science 2010, 327, 828–831. [Google Scholar] [CrossRef] [PubMed]
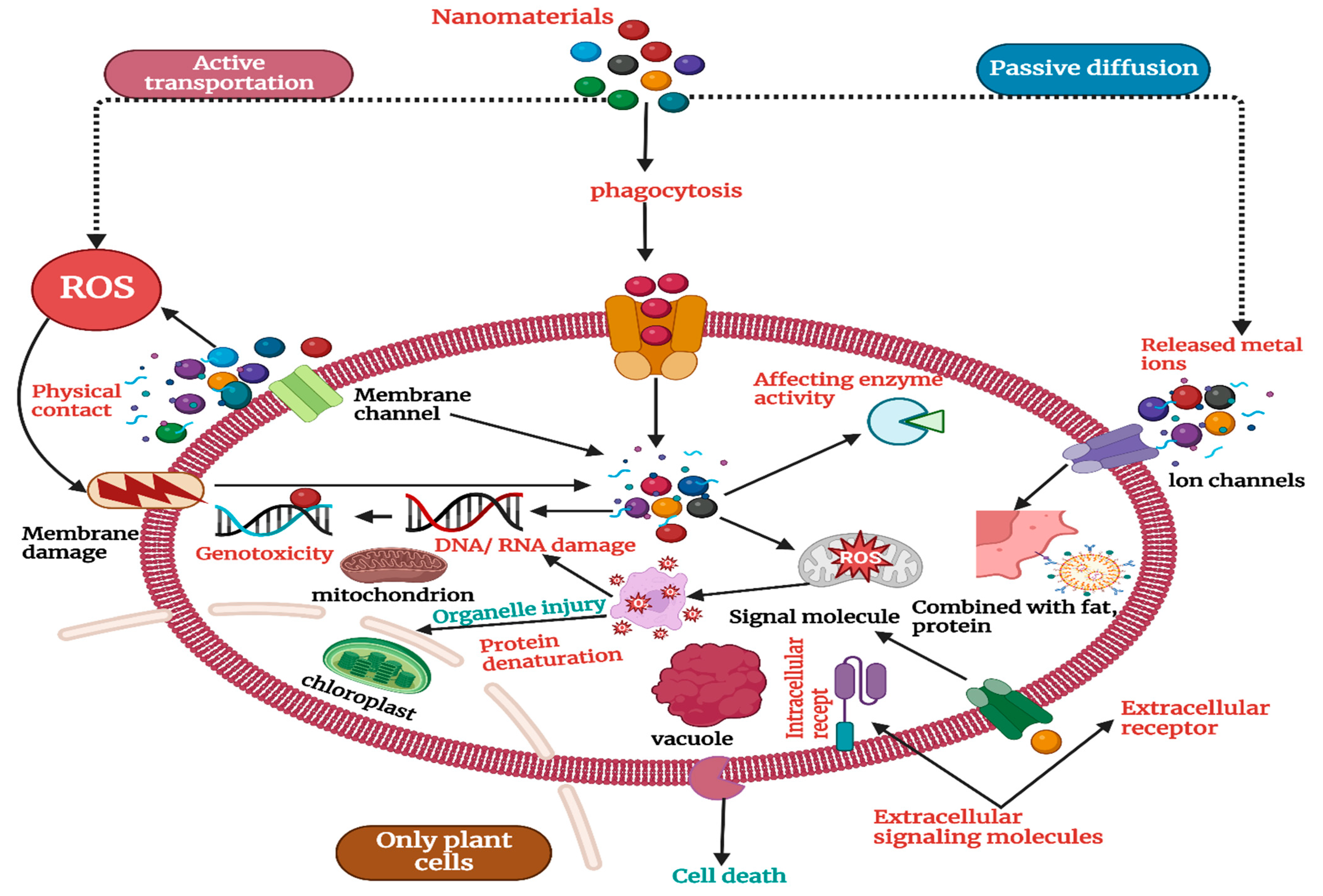
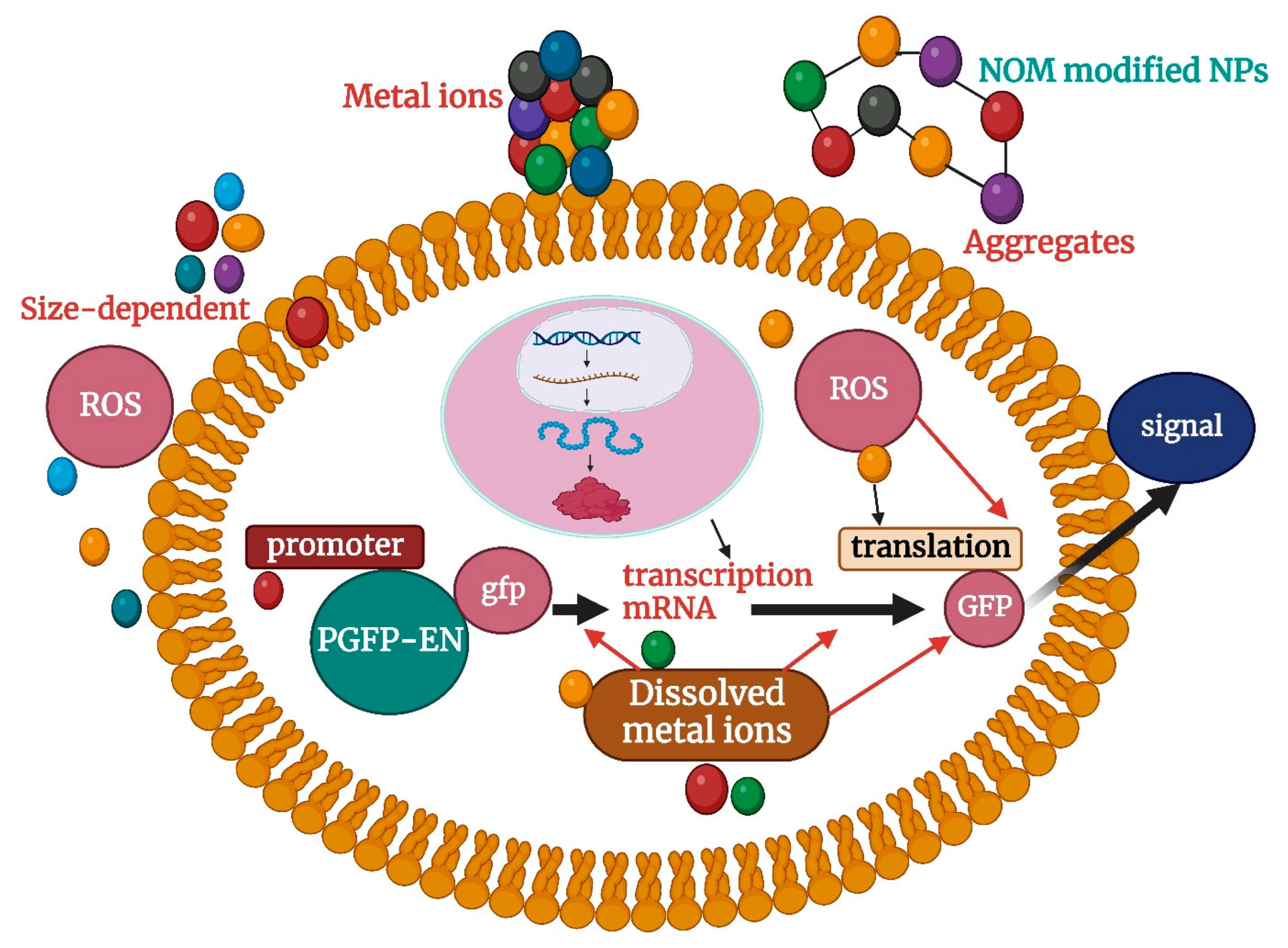
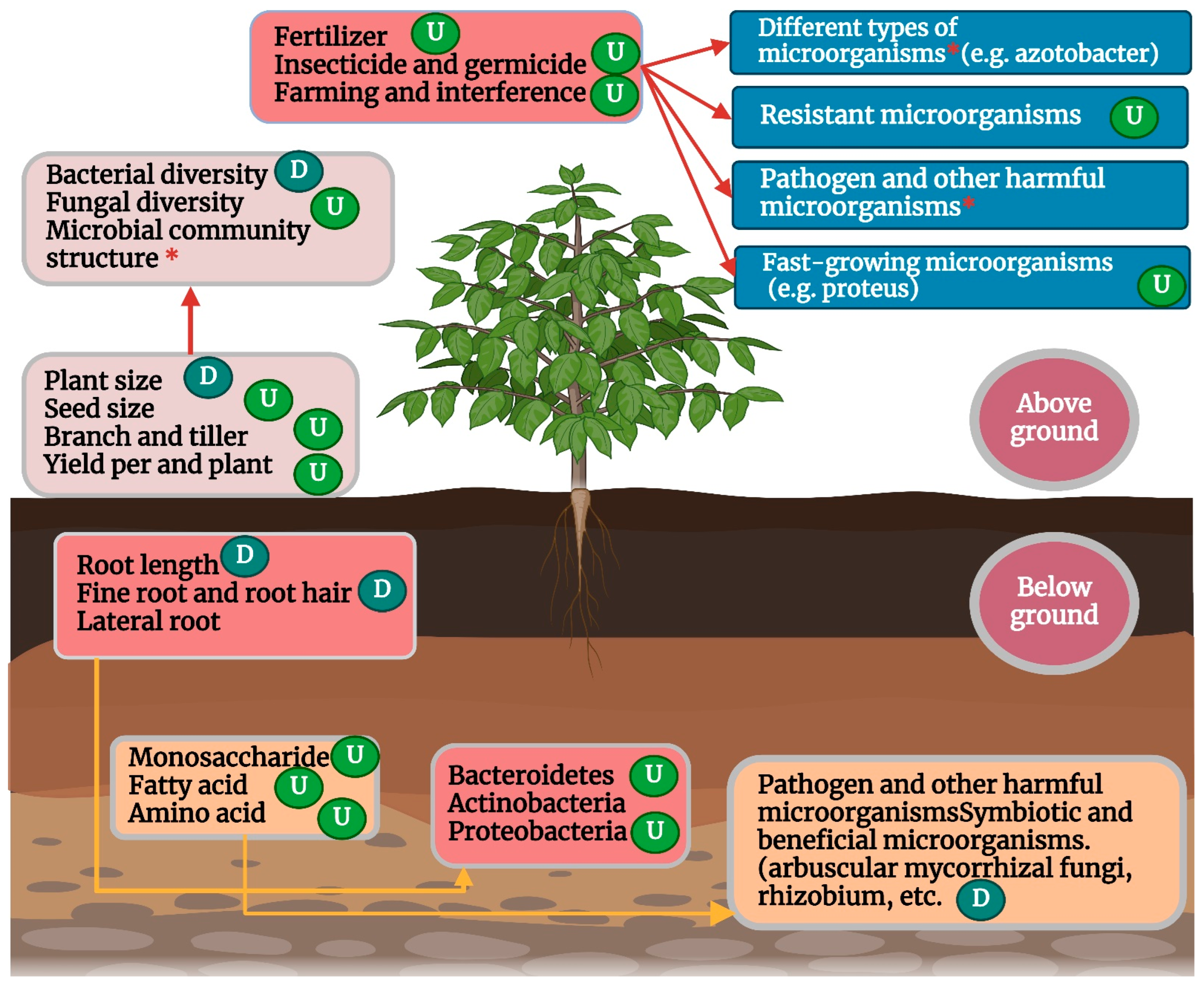
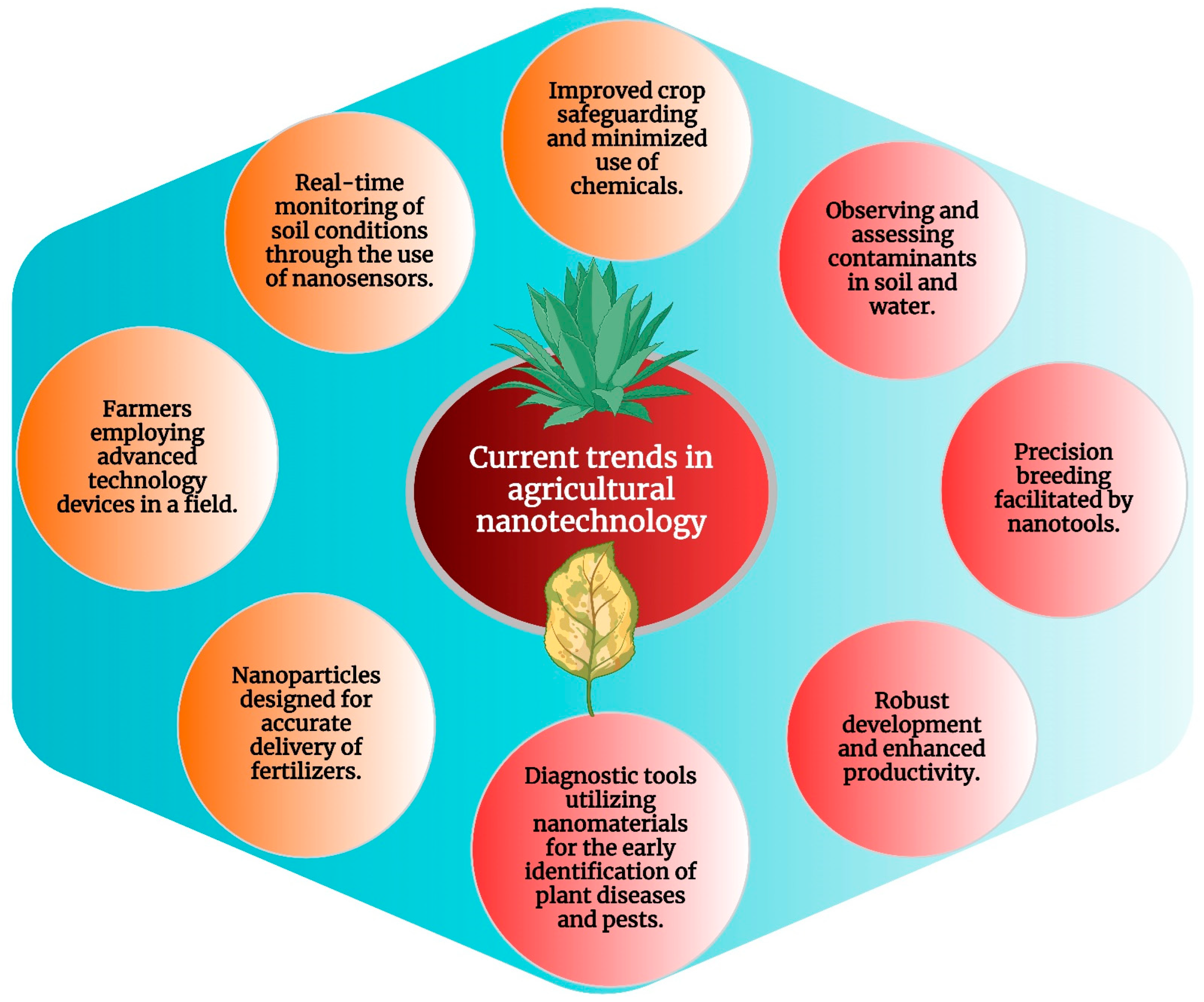
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).