
Role of Gut Microecology in the Pathogenesis of Drug-Induced Liver Injury and Emerging Therapeutic Strategies

Abstract
1. Introduction
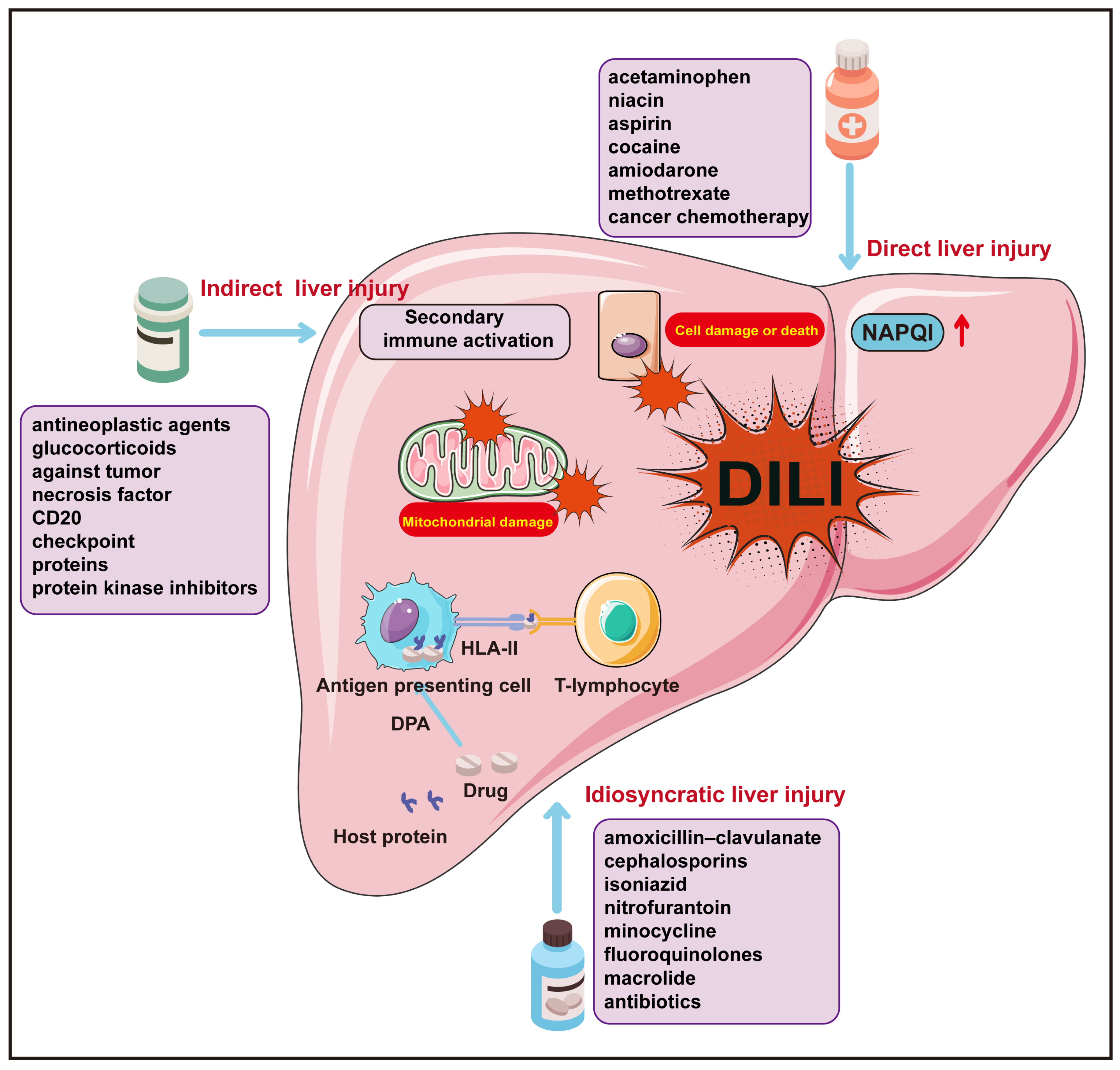
2. Drug-Induced Liver Injury (DILI)

{kind=link}
{kind=link}
| Drug Type | Common Drugs |
|---|---|
| Nonsteroidal Anti-inflammatory | APAP, Celecoxib, Diclofenac, Nimesulide |
| Herbal And Dietary Supplements | Polygonum Multiflorum Thunb, Cinobufacini, Xianlinggubao, Guizhi Fuling Capsule, Tripterygium Wilfordii |
| Anti-Tuberculosis | Isoniazid, Rifampin, Pyrazinamide, Thiacetazone, Protionamide |
| Anti-Fungal Drugs | Ketoconazole, Fluconazole, Itraconazole, Voriconazole |
| Antibiotic | Amoxycillin/Clavulanic Acid, Vancomycin, Pefloxacin, Enoxacin, Ofloxacin, Ciprofloxacin, Roxithromycin, Azithromycin, Clarithromycin, Vancomycin, Norvancomycin, Lincomycin, Clindamycin |
| Central Nervous System Drugs | Chlorpromazine, Trifluoperazine, Risperidone, Phenobarbital, Valproate, Carbamazepine |
| Iron | Polysaccharide Iron Complex, Ferrous Succinate, Ferrous Fumarate, Ferrous Gluconate |
| Direct Liver Injury | Idiosyncratic Liver Injury | Indirect Liver Injury | |
|---|---|---|---|
| Specific Mechanisms | The direct (inherent) liver injury of drugs and their active metabolites, along with the body’s intrinsic pathophysiological damage response. | Pharmacometabolic dysfunctions associated with human genetic polymorphism, or drug–host protein conjugate specificity and human leukocyte antigen (HLA)-restricted acquired immune responses. | The biological activity secondary to pharmaceuticals or their active metabolites often exert hepatotoxic effects indirectly through modulating the immune system. |
| Clinical Manifestations | Elevated liver enzymes, acute hepatic necrosis, hepatic sinusoidal obstruction syndrome, acute fatty liver, and nodular regeneration, among others. | Acute hepatocellular injury, cholestatic hepatitis, mixed hepatitis, simple cholestasis, and chronic hepatitis, among others. | Acute hepatitis, immune-mediated hepatitis, steatohepatitis, and chronic hepatitis, among others. |
| Common Drugs | Acetaminophen, aspirin, methotrexate, and other antineoplastic chemotherapy agents, highly effective antiretroviral drugs, synthetic anabolic steroids, statins, cyclosporine, heparin, valproic acid, niacin, butyrate, cocaine, amiodarone, tacrolimus, and so forth. | Amoxicillin/clavulanate, flucloxacillin, cephalosporins, macrolides, nitrofurantoin, minocycline, allopurinol, propylthiouracil, diclofenac, leflunomide, thalidomide, lapatinib, pazopanib, flutamide, and so forth. | Immunotherapy checkpoint inhibitors, antitumor necrosis factor monoclonal antibodies, anti-CD20 monoclonal antibodies, protein kinase inhibitors, corticosteroids, select anti-neoplastic agents, and drugs related to energy metabolism and interfering substances. |
| Study Subjects | Drug Involved | Microbial Composition Shifts Compared to Normal Controls | Source | |
|---|---|---|---|---|
| Increased | Decreased | |||
| Mice | Methotrexate (MTX) | Firmicutes; Actinobacteriota; Proteobacteria; Blautia; Ruminococcu_torques_group; Staphylococcus; Enterorhabdus; Enterococcus | Bacteroidota; Verrucomicrobiota; Lactobacillus; Allobaculum; norank_f_Muribaculaceae; Dubosiella; Ruminococcus | [27] |
| DILI patient; Healthy controls | - | Bacteroides; Alistipes | Acetobacteroides; Blautia; Caloramator; Coprococcus; Flavobacterium; Lachnospira; Natronincola; Oscillospira; Pseudobutyrivibrio; Shuttleworthia | [28] |
| DILI patient; Healthy controls | - | Bacteroidota; Firmicutes; Fusobacteriota; Acidobacteriota; Streptococcus; Faecalibacterium; Bacteroides; Klebsiella; Enterococcus; Veillonella | Proteobacteria; Verrucomicrobiota; Desulfobacterota; Streptococcus; Faecalibacterium; Bacteroides; Klebsiella; Blautia; Ralstonia; Dialister | [29] |
| Mice | Perfluoro-octanoic acid | Parabacteroides | Dehalobacterium; Bacteroides; Lactobacillus; Bifidobacterium | [30] |
| Mice | Acetaminophen (APAP) | Akkermansia muciniphila; Verrucomicrobiales; Verrucomicrobiaceae | Firmicutes; Clostridia; Clostridiales; Lachnospiraceae | [31] |
| Rats | Bisphenol A (BPA) | Prevotellaceae_NK3B31_group; Firmicutes | Prevotella_9; Bacteroidetes; Ruminococcaceae_UC-G014 | [32] |
| DILI patient; Healthy controls | Antithyroid drugs (ATDs) | Eubacteriumrectale; Romboutsia Dorea | Faecalibacterium; Clostridium_sensu_stricto_1; Bacteroides | [33] |
| Rats | Oral iron | Defluviitaleaceae UCG-011; RuminococcaceaeUCG-014 Coprococcus 1 | Lachnospiraceae FCS020; genus Allobaculum | [34] |
| Rats | Tacrine | Bacteroides | Lactobacillus | [35] |
3. Gut Microbiota
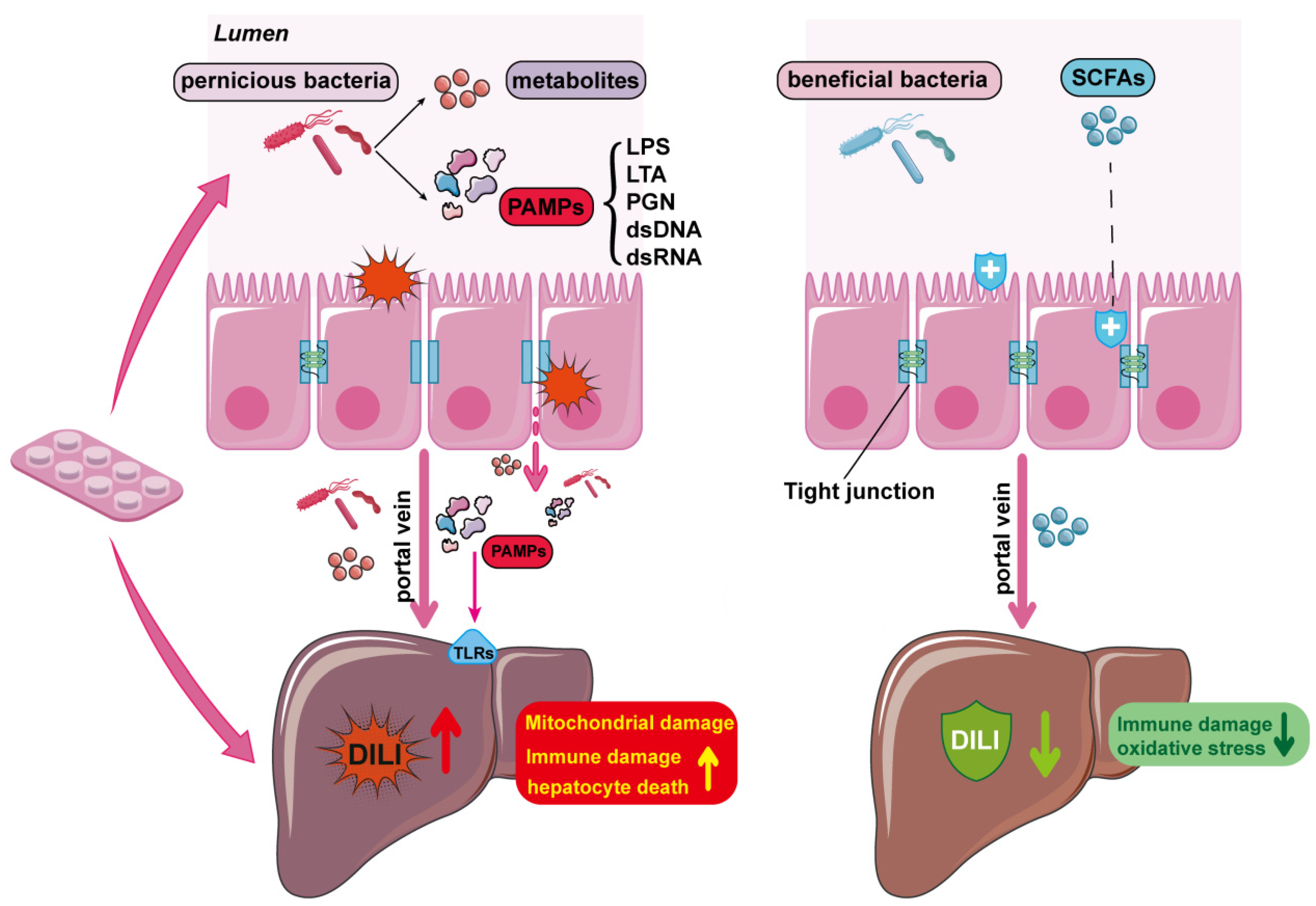
4. Effects of Gut Microbiota on DILI
4.1. Effects of Gut Microbiota on APAP-Induced Liver Injury
4.1.1. Effects of Gut Microbiota Abundance and Diversity on APAP-Induced Liver Injury
4.1.2. Effects of Gut Microbial Metabolites on APAP-Induced Liver Injury
4.1.3. Effects of Gut Barrier on APAP-Induced Liver Injury
4.2. Effect of Gut Microbiota on the Hepatotoxicity of Other Drugs
4.2.1. Effect of Gut Microbiota on the Anti-Tuberculosis Drug-Induced Liver Injury
4.2.2. Effect of Gut Microbiota on Iron-Induced Liver Injury
4.2.3. Effect of Gut Microbiota on the Herbal Medicine-Induced Liver Injury
5. Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Le Daré, B.; Ferron, P.J.; Gicquel, T. Once upon a time the hepatotoxicity. Med. Sci. M/S 2021, 37, 235–241. [Google Scholar]
- Garcia-Cortes, M.; Robles-Diaz, M.; Stephens, C.; Ortega-Alonso, A.; Lucena, M.I.; Andrade, R.J. Drug induced liver injury: An update. Arch. Toxicol. 2020, 94, 3381–3407. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-X.; Deng, X.-R.; Zhang, C.-H.; Yuan, H.-J. Gut microbiota and metabolic syndrome. Chin. Med. J. 2020, 133, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Gong, S.; Lan, T.; Zeng, L.; Luo, H.; Yang, X.; Li, N.; Chen, X.; Liu, Z.; Li, R.; Win, S.; et al. Gut microbiota mediates diurnal variation of acetaminophen induced acute liver injury in mice. J. Hepatol. 2018, 69, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef]
- Wiest, R.; Albillos, A.; Trauner, M.; Bajaj, J.S.; Jalan, R. Targeting the gut-liver axis in liver disease. J. Hepatol. 2017, 67, 1084–1103. [Google Scholar] [CrossRef]
- Tilg, H.; Zmora, N.; Adolph, T.E.; Elinav, E. The intestinal microbiota fuelling metabolic inflammation. Nat. Rev. Immunol. 2020, 20, 40–54. [Google Scholar] [CrossRef]
- Russo, M.W.; Galanko, J.A.; Shrestha, R.; Fried, M.W.; Watkins, P. Liver transplantation for acute liver failure from drug induced liver injury in the united states. Liver Transplant. 2004, 10, 1018–1023. [Google Scholar] [CrossRef]
- Björnsson, E.S.; Andrade, R.J. Long-term sequelae of drug-induced liver injury. J. Hepatol. 2022, 76, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Hoofnagle, J.H.; Björnsson, E.S. Drug-induced liver injury—Types and phenotypes. N. Engl. J. Med. 2019, 381, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Björnsson, E.; Jerlstad, P.; Bergqvist, A.; Olsson, R. Fulminant drug-induced hepatic failure leading to death or liver transplantation in sweden. Scand. J. Gastroenterol. 2005, 40, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Liu, Y.; Shang, J.; Xie, Q.; Li, J.; Yan, M.; Xu, J.; Niu, J.; Liu, J.; Watkins, P.B.; et al. Incidence and etiology of drug-induced liver injury in mainland china. Gastroenterology 2019, 156, 2230–2241. [Google Scholar] [CrossRef] [PubMed]
- Kaplowitz, N. Idiosyncratic drug hepatotoxicity. Nat. Rev. Drug Discov. 2005, 4, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Russmann, S.; Jetter, A.; Kullak-Ublick, G.A. Pharmacogenetics of drug-induced liver injury. Hepatology 2010, 52, 748–761. [Google Scholar] [CrossRef] [PubMed]
- Hui, C.L.; Lee, Z.J. Hepatic disorders associated with exogenous sex steroids: Mr imaging findings. Abdom. Radiol. 2019, 44, 2436–2447. [Google Scholar] [CrossRef]
- Lu, Z.; Luo, Q.; Zhao, L.; Shi, Y.; Wang, N.; Wang, L.; Han, Z. The mutational features of aristolochic acid-induced mouse and human liver cancers. Hepatology 2020, 71, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Lammert, C.; Einarsson, S.; Saha, C.; Niklasson, A.; Bjornsson, E.; Chalasani, N. Relationship between daily dose of oral medications and idiosyncratic drug-induced liver injury: Search for signals. Hepatology 2008, 47, 2003–2009. [Google Scholar] [CrossRef]
- Chen, M.; Borlak, J.; Tong, W. Predicting idiosyncratic drug-induced liver injury: Some recent advances. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 721–723. [Google Scholar] [CrossRef]
- Carrascosa, M.F.; Salcines-Caviedes, J.R.; Lucena, M.I.; Andrade, R.J. Acute liver failure following atorvastatin dose escalation: Is there a threshold dose for idiosyncratic hepatotoxicity? J. Hepatol. 2015, 62, 751–752. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Y.; Salem, J.-E.; Cohen, J.V.; Chandra, S.; Menzer, C.; Ye, F.; Zhao, S.; Das, S.; Beckermann, K.E.; Ha, L.; et al. Fatal toxic effects associated with immune checkpoint inhibitors: A systematic review and meta-analysis. JAMA Oncol. 2018, 4, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Vaddepally, R.K.; Kharel, P.; Pandey, R.; Garje, R.; Chandra, A.B. Review of indications of fda-approved immune checkpoint inhibitors per nccn guidelines with the level of evidence. Cancers 2020, 12, 738. [Google Scholar] [CrossRef] [PubMed]
- Peeraphatdit, T.; Wang, J.; Odenwald, M.A.; Hu, S.; Hart, J.; Charlton, M.R. Hepatotoxicity from immune checkpoint inhibitors: A systematic review and management recommendation. Hepatology 2020, 72, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Russmann, S.; Kullak-Ublick, G.A.; Grattagliano, I. Current concepts of mechanisms in drug-induced hepatotoxicity. Curr. Med. Chem. 2009, 16, 3041–3053. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.J.; Aithal, G.P.; Björnsson, E.S.; Kaplowitz, N.; Kullak-Ublick, G.A.; Larrey, D.; Karlsen, T.H.; European Association for the Study of the Liver. Easl clinical practice guidelines: Drug-induced liver injury. J. Hepatol. 2019, 70, 1222–1261. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, S.; Xu, Y.; Sun, W.; Feng, Y.; Liang, D.; Guan, Y. Integrated microbiome and metabolome analysis reveals correlations between gut microbiota components and metabolic profiles in mice with methotrexate-induced hepatoxicity. Drug Des. Dev. Ther. 2022, 16, 3877–3891. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Diaz, C.; Taminiau, B.; García-García, A.; Cueto, A.; Robles-Díaz, M.; Ortega-Alonso, A.; Martín-Reyes, F.; Daube, G.; Sanabria-Cabrera, J.; Jimenez-Perez, M.; et al. Microbiota diversity in nonalcoholic fatty liver disease and in drug-induced liver injury. Pharmacol. Res. 2022, 182, 106348. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Liu, M.; Wang, Q.; Chen, S.; Guo, X. Combined analysis of 16s rdna sequencing and metabolomics to find biomarkers of drug-induced liver injury. Sci. Rep. 2023, 13, 15138. [Google Scholar] [CrossRef]
- Wang, G.; Pan, R.; Liang, X.; Wu, X.; Wu, Y.; Zhang, H.; Zhao, J.; Chen, W. Perfluorooctanoic acid-induced liver injury is potentially associated with gut microbiota dysbiosis. Chemosphere 2021, 266, 129004. [Google Scholar] [CrossRef]
- Schneider, K.M.; Elfers, C.; Ghallab, A.; Schneider, C.V.; Galvez, E.J.; Mohs, A.; Gui, W.; Candels, L.S.; Wirtz, T.H.; Zuehlke, S.; et al. Intestinal dysbiosis amplifies acetaminophen-induced acute liver injury. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 909–933. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, B.; Tian, L.; Jiang, X.; Li, X.; Cai, D.; Sun, J.; Bai, W.; Jin, Y. Exposure to bisphenol a caused hepatoxicity and intestinal flora disorder in rats. Int. J. Mol. Sci. 2022, 23, 8042. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhao, F.; Lin, B.; Feng, J.; Wu, X.; Liu, Y.; Zhao, L.; Zhu, B.; Wei, Y. Gut microbiota participates in antithyroid drug induced liver injury through the lipopolysaccharide related signaling pathway. Front. Pharmacol. 2020, 11, 598170. [Google Scholar] [CrossRef]
- Fang, S.; Zhuo, Z.; Yu, X.; Wang, H.; Feng, J. Oral administration of liquid iron preparation containing excess iron induces intestine and liver injury, impairs intestinal barrier function and alters the gut microbiota in rats. J. Trace Elem. Med. Biol. 2018, 47, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Yip, L.Y.; Aw, C.C.; Lee, S.H.; Hong, Y.S.; Ku, H.C.; Xu, W.H.; Chan, J.M.X.; Cheong, E.J.Y.; Chng, K.R.; Ng, A.H.Q.; et al. The liver-gut microbiota axis modulates hepatotoxicity of tacrine in the rat. Hepatology 2018, 67, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Björnsson, E.S.; Bergmann, O.M.; Björnsson, H.K.; Kvaran, R.B.; Olafsson, S. Incidence, presentation, and outcomes in patients with drug-induced liver injury in the general population of iceland. Gastroenterology 2013, 144, 1419–1425, 1425.e1–e3; quiz e19–e20. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Bonkovsky, H.L.; Fontana, R.; Lee, W.; Stolz, A.; Talwalkar, J.; Reddy, K.R.; Watkins, P.B.; Navarro, V.; Barnhart, H.; et al. Features and outcomes of 899 patients with drug-induced liver injury: The dilin prospective study. Gastroenterology 2015, 148, 1340–1352. [Google Scholar] [CrossRef] [PubMed]
- Robles–Diaz, M.; Lucena, M.I.; Kaplowitz, N.; Stephens, C.; Medina–Cáliz, I.; González–Jimenez, A.; Ulzurrun, E.; Gonzalez, A.F.; Fernandez, M.C.; Romero–Gómez, M.; et al. Use of hy’s law and a new composite algorithm to predict acute liver failure in patients with drug-induced liver injury. Gastroenterology 2014, 147, 109–118. [Google Scholar] [CrossRef] [PubMed]
- De Abajo, F.J.; Montero, D.; Madurga, M.; Rodríguez, L.A.G. Acute and clinically relevant drug-induced liver injury: A population based case-control study. Br. J. Clin. Pharmacol. 2004, 58, 71–80. [Google Scholar] [CrossRef]
- Njoku, D.B. Drug-induced hepatotoxicity: Metabolic, genetic and immunological basis. Int. J. Mol. Sci. 2014, 15, 6990–7003. [Google Scholar] [CrossRef]
- Pessayre, D.; Fromenty, B.; Berson, A.; Robin, M.-A.; Lettéron, P.; Moreau, R.; Mansouri, A. Central role of mitochondria in drug-induced liver injury. Drug Metab. Rev. 2012, 44, 34–87. [Google Scholar] [CrossRef] [PubMed]
- Au, J.S.; Navarro, V.J.; Rossi, S. Review article: Drug-induced liver injury—Its pathophysiology and evolving diagnostic tools. Aliment. Pharmacol. Ther. 2011, 34, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.J.; Lucena, M.I.; Alonso, A.; García-Cortés, M.; García-Ruiz, E.; Benitez, R.; Fernández, M.C.; Pelaez, G.; Romero, M.; Corpas, R.; et al. Hla class ii genotype influences the type of liver injury in drug-induced idiosyncratic liver disease. Hepatology 2004, 39, 1603–1612. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Huo, Y.; Yin, S.; Hu, H. Mechanisms of acetaminophen-induced liver injury and its implications for therapeutic interventions. Redox Biol. 2018, 17, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.C.; Mao, Y.M.; Chen, C.W.; Chen, J.J.; Chen, J.; Cong, W.M.; Ding, Y.; Duan, Z.P.; Fu, Q.C.; Guo, X.Y.; et al. Csh guidelines for the diagnosis and treatment of drug-induced liver injury. Hepatol. Int. 2017, 11, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; O’Flaherty, S.; Claesson, M.J.; Turroni, F.; Klaenhammer, T.R.; van Sinderen, D.; O’Toole, P.W. Genome-scale analyses of health-promoting bacteria: Probiogenomics. Nat. Rev. Microbiol. 2009, 7, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Milosevic, I.; Vujovic, A.; Barac, A.; Djelic, M.; Korac, M.; Radovanovic Spurnic, A.; Gmizic, I.; Stevanovic, O.; Djordjevic, V.; Lekic, N.; et al. Gut-liver axis, gut microbiota, and its modulation in the management of liver diseases: A review of the literature. Int. J. Mol. Sci. 2019, 20, 395. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Bai, C.; Brown, T.D.; Hood, L.E.; Tian, Q. Human gut microbiota and gastrointestinal cancer. Genom. Proteom. Bioinform. 2018, 16, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 101943. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef]
- Kang, X.; Liu, C.; Ding, Y.; Ni, Y.; Ji, F.; Lau, H.C.H.; Jiang, L.; Sung, J.J.; Wong, S.H.; Yu, J. Roseburia intestinalis generated butyrate boosts anti-pd-1 efficacy in colorectal cancer by activating cytotoxic cd8(+) t cells. Gut 2023, 72, 2112–2122. [Google Scholar] [CrossRef]
- Doestzada, M.; Vila, A.V.; Zhernakova, A.; Koonen, D.P.; Weersma, R.K.; Touw, D.J.; Kuipers, F.; Wijmenga, C.; Fu, J. Pharmacomicrobiomics: A novel route towards personalized medicine? Protein Cell 2018, 9, 432–445. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.L.; Patterson, A.D. The gut microbiome: An orchestrator of xenobiotic metabolism. Acta Pharm. Sin. B 2020, 10, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Chen, L.; Shen, Z. Mechanisms of gastrointestinal microflora on drug metabolism in clinical practice. Saudi Pharm. J. 2019, 27, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nat. Rev. Mol. Cell Biol. 2014, 15, 19–33. [Google Scholar] [CrossRef]
- Chopyk, D.M.; Grakoui, A. Contribution of the intestinal microbiome and gut barrier to hepatic disorders. Gastroenterology 2020, 159, 849–863. [Google Scholar] [CrossRef]
- Raleigh, D.R.; Marchiando, A.M.; Zhang, Y.; Shen, L.; Sasaki, H.; Wang, Y.; Long, M.; Turner, J.R.; Higashi, T.; Miller, A.L.; et al. Tight junction-associated marvel proteins marveld3, tricellulin, and occludin have distinct but overlapping functions. Mol. Biol. Cell 2010, 21, 1200–1213. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Ambort, D.; Pelaseyed, T.; Schütte, A.; Gustafsson, J.K.; Ermund, A.; Subramani, D.B.; Holmén-Larsson, J.M.; Thomsson, K.A.; Bergström, J.H.; et al. Composition and functional role of the mucus layers in the intestine. Cell. Mol. Life Sci. 2011, 68, 3635–3641. [Google Scholar] [CrossRef]
- Hou, Q.; Huang, J.; Ayansola, H.; Masatoshi, H.; Zhang, B. Intestinal stem cells and immune cell relationships: Potential therapeutic targets for inflammatory bowel diseases. Front. Immunol. 2020, 11, 623691. [Google Scholar] [CrossRef]
- Inamine, T.; Schnabl, B. Immunoglobulin a and liver diseases. J. Gastroenterol. 2018, 53, 691–700. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory iga’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Ladinsky, M.S.; Yu, K.B.; Sanders, J.G.; Yoo, B.B.; Chou, W.-C.; Conner, M.E.; Earl, A.M.; Knight, R.; Bjorkman, P.J.; et al. Gut microbiota utilize immunoglobulin a for mucosal colonization. Science 2018, 360, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Chairatana, P.; Nolan, E.M. Defensins, lectins, mucins, and secretory immunoglobulin a: Microbe-binding biomolecules that contribute to mucosal immunity in the human gut. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Hooper, L.V. Antimicrobial defense of the intestine. Immunity 2015, 42, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Litvak, Y.; Mon, K.K.; Nguyen, H.; Chanthavixay, G.; Liou, M.; Velazquez, E.M.; Kutter, L.; Alcantara, M.A.; Byndloss, M.X.; Tiffany, C.R.; et al. Commensal enterobacteriaceae protect against salmonella colonization through oxygen competition. Cell Host Microbe 2019, 25, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.M.; Brummer, R.J.; Derrien, M.; MacDonald, T.T.; Troost, F.; Cani, P.D.; Theodorou, V.; Dekker, J.; Méheust, A.; de Vos, W.M.; et al. Homeostasis of the gut barrier and potential biomarkers. Am. J. Physiol.-Gastroint. Liver Physiol. 2017, 312, G171–G193. [Google Scholar] [CrossRef]
- Stacy, A.; Andrade-Oliveira, V.; McCulloch, J.A.; Hild, B.; Oh, J.H.; Perez-Chaparro, P.J.; Sim, C.K.; Lim, A.I.; Link, V.M.; Enamorado, M.; et al. Infection trains the host for microbiota-enhanced resistance to pathogens. Cell 2021, 184, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef]
- Wang, H.J.; Gao, B.; Zakhari, S.; Nagy, L.E. Inflammation in alcoholic liver disease. Annu. Rev. Nutr. 2012, 32, 343–368. [Google Scholar] [CrossRef]
- Hiippala, K.; Jouhten, H.; Ronkainen, A.; Hartikainen, A.; Kainulainen, V.; Jalanka, J.; Satokari, R. The potential of gut commensals in reinforcing intestinal barrier function and alleviating inflammation. Nutrients 2018, 10, 988. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Liu, R.; Wang, D. Research progress of glutathione. Chin. J. Pharm. (Online Ed.) 2019, 17, 141–148. [Google Scholar]
- Kaplowitz, N. Acetaminophen hepatoxicity: What do we know, what don’t we know, and what do we do next? Hepatology 2004, 40, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Reuben, A.; Tillman, H.; Fontana, R.J.; Davern, T.; McGuire, B.; Stravitz, R.T.; Durkalski, V.; Larson, A.M.; Liou, I.; Fix, O.; et al. Outcomes in adults with acute liver failure between 1998 and 2013: An observational cohort study. Ann. Intern. Med. 2016, 164, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.C.; Davidson, J.S.; Martin, K.; Baird, A.; Hayes, P.C.; Simpson, K.J. Acute liver failure in scotland: Changes in aetiology and outcomes over time (the scottish look-back study). Aliment. Pharmacol. Ther. 2017, 45, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Yang, X.; Won, K.-J.; Leone, V.A.; Chang, E.B.; Guzman, G.; Ko, Y.; Bae, O.-N.; Lee, H.; Jeong, H. Phenylpropionic acid produced by gut microbiota alleviates acetaminophen-induced hepatotoxicity. Gut Microbes 2023, 15, 2231590. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.; Morley, S.D.; Raja, A.K.; Vandeputte, M.; Samuel, K.; Waterfall, M.; Homer, N.Z.M.; Hayes, P.C.; Fallowfield, J.A.; Plevris, J.N. Metabolism of acetaminophen by enteric epithelial cells mitigates hepatocellular toxicity in vitro. J. Clin. Med. 2023, 12, 3995. [Google Scholar] [CrossRef] [PubMed]
- Joh, E.H.; Kim, D.H. A sensitive liquid chromatography-electrospray tandem mass spectrometric method for lancemaside a and its metabolites in plasma and a pharmacokinetic study in mice. J. Chromatogr. B 2010, 878, 1875–1880. [Google Scholar] [CrossRef]
- Gauhar, S.; Ali, S.A.; Naqvi, S.B.; Shoaib, M.H. Report: Pharmacokinetic and drug interaction studies of pefloxacin with paracetamol (nnaid) in healthy volunteers in pakistan. Pak. J. Pharm. Sci. 2014, 27, 389–395. [Google Scholar]
- Kim, D.H.; Kobashi, K. The role of intestinal flora in metabolism of phenolic sulfate esters. Biochem. Pharmacol. 1986, 35, 3507–3510. [Google Scholar] [CrossRef]
- Kwon, A.-R.; Oh, T.-G.; Kim, D.-H.; Choi, E.-C. Molecular cloning of the arylsulfate sulfotransferase gene and characterization of its product from enterobacter amnigenus ar-37. Protein Expr. Purif. 1999, 17, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., A human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Lancaster, E.M.; Hiatt, J.R.; Zarrinpar, A. Acetaminophen hepatotoxicity: An updated review. Arch. Toxicol. 2015, 89, 193–199. [Google Scholar] [CrossRef]
- Xia, J.; Lv, L.; Liu, B.; Wang, S.; Zhang, S.; Wu, Z.; Yang, L.; Bian, X.; Wang, Q.; Wang, K.; et al. Akkermansia muciniphila ameliorates acetaminophen-induced liver injury by regulating gut microbial composition and metabolism. Microbiol. Spectr. 2022, 10, e159621. [Google Scholar] [CrossRef]
- You, J.S.; Yong, J.H.; Kim, G.H.; Moon, S.; Nam, K.T.; Ryu, J.H.; Yoon, M.Y.; Yoon, S.S. Commensal-derived metabolites govern vibrio cholerae pathogenesis in host intestine. Microbiome 2019, 7, 132. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.-K.; Liu, H.-H.; Chen, G.-H.; Zhu, J.-Q.; Zheng, S.-Y.; Zhao, D.; Diao, J.; Jia, H.; Zhang, D.-D.; Chen, S.-X.; et al. Oridonin alters hepatic urea cycle via gut microbiota and protects against acetaminophen-induced liver injury. Oxid. Med. Cell. Longev. 2021, 2021, 3259238. [Google Scholar] [CrossRef]
- Kim, J.-K.; Choi, M.S.; Jeong, J.-J.; Lim, S.-M.; Kim, I.S.; Yoo, H.H.; Kim, D.-H. Effect of probiotics on pharmacokinetics of orally administered acetaminophen in mice. Drug. Metab. Dispos. 2018, 46, 122–130. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, R.L.; Kakkar, P. Modulation of bax/bcl-2 and caspases by probiotics during acetaminophen induced apoptosis in primary hepatocytes. Food. Chem. Toxicol. 2011, 49, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Neag, M.A.; Catinean, A.; Muntean, D.M.; Pop, M.R.; Bocsan, C.I.; Botan, E.C.; Buzoianu, A.D. Probiotic bacillus spores protect against acetaminophen induced acute liver injury in rats. Nutrients 2020, 12, 632. [Google Scholar] [CrossRef]
- Zheng, N.; Gu, Y.; Hong, Y.; Sheng, L.; Chen, L.; Zhang, F.; Hou, J.; Zhang, W.; Zhang, Z.; Jia, W.; et al. Vancomycin pretreatment attenuates acetaminophen-induced liver injury through 2-hydroxybutyric acid. J. Pharm. Anal. 2020, 10, 560–570. [Google Scholar] [CrossRef]
- Possamai, L.A.; McPhail, M.J.; Khamri, W.; Wu, B.; Concas, D.; Harrison, M.; Williams, R.; Cox, R.D.; Cox, I.J.; Anstee, Q.M.; et al. The role of intestinal microbiota in murine models of acetaminophen-induced hepatotoxicity. Liver Int. 2015, 35, 764–773. [Google Scholar] [CrossRef]
- Bone, E.; Tamm, A.; Hill, M. The production of urinary phenols by gut bacteria and their possible role in the causation of large bowel cancer. Am. J. Clin. Nutr. 1976, 29, 1448–1454. [Google Scholar] [CrossRef]
- Clayton, T.A.; Baker, D.; Lindon, J.C.; Everett, J.R.; Nicholson, J.K. Pharmacometabonomic identification of a significant host-microbiome metabolic interaction affecting human drug metabolism. Proc. Natl. Acad. Sci. USA 2009, 106, 14728–14733. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, B.J.; Liu, K.H.; Owens, J.A.; Hunter-Chang, S.; Camacho, M.C.; Eboka, R.U.; Chandrasekharan, B.; Baker, N.F.; Darby, T.M.; Robinson, B.S.; et al. Gut-resident lactobacilli activate hepatic nrf2 and protect against oxidative liver injury. Cell Metab. 2020, 31, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Jaeschke, H.; Mcgill, M.R.; Ramachandran, A. Oxidant stress, mitochondria, and cell death mechanisms in drug-induced liver injury: Lessons learned from acetaminophen hepatotoxicity. Drug Metab. Rev. 2012, 44, 88–106. [Google Scholar] [CrossRef] [PubMed]
- Kakan, X.; Chen, P.; Zhang, J. Clock gene mper2 functions in diurnal variation of acetaminophen induced hepatotoxicity in mice. Exp. Toxicol. Pathol. 2011, 63, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhu, L.; Jiang, L. Dynamic regulation of gut clostridium-derived short-chain fatty acids. Trends Biotechnol. 2022, 40, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. Scfa: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kong, C.; Mao, Y.; Huang, J.; Chen, H.; Han, B.; Wang, L. Ampicillin exacerbates acetaminophen-induced acute liver injury by inducing intestinal microbiota imbalance and butyrate reduction. Liver Int. 2023, 43, 865–877. [Google Scholar] [CrossRef]
- Hodgkinson, K.; El Abbar, F.; Dobranowski, P.; Manoogian, J.; Butcher, J.; Figeys, D.; Mack, D.; Stintzi, A. Butyrate’s role in human health and the current progress towards its clinical application to treat gastrointestinal disease. Clin. Nutr. 2023, 42, 61–75. [Google Scholar] [CrossRef]
- Li, Y.; Hong, X.; Liang, L.; Wang, X.; Ladd-Acosta, C. Association between acetaminophen metabolites and cyp2e1 dna methylation level in neonate cord blood in the boston birth cohort. Clin. Epigenet. 2023, 15, 132. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wu, R.; Wang, F.; Li, S.; Li, L.; Li, Y.; Qin, P.; Wei, M.; Yang, J.; Wu, J.; et al. Liberation of daidzein by gut microbial β-galactosidase suppresses acetaminophen-induced hepatotoxicity in mice. Cell Host Microbe 2023, 31, 766–780. [Google Scholar] [CrossRef] [PubMed]
- Tak, J.; Kim, Y.S.; Kim, T.H.; Park, G.-C.; Hwang, S.; Kim, S.G. Gα(12) overexpression in hepatocytes by er stress exacerbates acute liver injury via rock1-mediated mir-15a and alox12 dysregulation. Theranostics 2022, 12, 1570–1588. [Google Scholar] [CrossRef]
- Wang, M.; Liu, C.-Y.; Wang, T.; Yu, H.-M.; Ouyang, S.-H.; Wu, Y.-P.; Gong, H.-B.; Ma, X.-H.; Jiao, G.-L.; Fu, L.-L.; et al. (+)-clausenamide protects against drug-induced liver injury by inhibiting hepatocyte ferroptosis. Cell Death Dis. 2020, 11, 781. [Google Scholar] [CrossRef] [PubMed]
- Shojaie, L.; Iorga, A.; Dara, L. Cell death in liver diseases: A review. Int. J. Mol. Sci. 2020, 21, 9682. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Luo, Z.; Gong, S.; Win, S.; Kaplowitz, N.; Jiang, Y.; Chen, P. Intestinal epithelial chemokine (c-c motif) ligand 7 overexpression enhances acetaminophen-induced hepatotoxicity in mice. Am. J. Pathol. 2020, 190, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Chopyk, D.; Stuart, J.D.; Zimmerman, M.G.; Wen, J.; Gumber, S.; Suthar, M.S.; Thapa, M.; Czaja, M.J.; Grakoui, A. Acetaminophen intoxication rapidly induces apoptosis of intestinal crypt stem cells and enhances intestinal permeability. Hepatol. Commun. 2019, 3, 1435–1449. [Google Scholar] [CrossRef] [PubMed]
- Furin, J.; Cox, H.; Pai, M. Tuberculosis. Lancet 2019, 393, 1642–1656. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yang, Q.; Liu, B.; Dong, J.; Sun, L.; Zhu, Y.; Su, H.; Yang, J.; Yang, F.; Chen, X.; et al. Gut microbiota associated with pulmonary tuberculosis and dysbiosis caused by anti-tuberculosis drugs. J. Infect. 2019, 78, 317–322. [Google Scholar] [CrossRef]
- Cao, D.; Liu, W.; Lyu, N.; Li, B.; Song, W.; Yang, Y.; Zhu, J.; Zhang, Z.; Zhu, B. Gut mycobiota dysbiosis in pulmonary tuberculosis patients undergoing anti-tuberculosis treatment. Microbiol. Spectr. 2021, 9, e61521. [Google Scholar] [CrossRef]
- Chiu, C.-C.; Ching, Y.-H.; Li, Y.-P.; Liu, J.-Y.; Huang, Y.-T.; Huang, Y.-W.; Yang, S.-S.; Huang, W.-C.; Chuang, H.-L. Nonalcoholic fatty liver disease is exacerbated in high-fat diet-fed gnotobiotic mice by colonization with the gut microbiota from patients with nonalcoholic steatohepatitis. Nutrients 2017, 9, 1220. [Google Scholar] [CrossRef]
- Liu, N.; Liu, J.; Zheng, B.; Zeng, X.; Ye, Z.; Huang, X.; Liu, W.; Liu, Y.; Fang, Q.; Chen, L.; et al. Gut microbiota affects sensitivity to immune-mediated isoniazid-induced liver injury. Biomed. Pharmacother. 2023, 160, 114400. [Google Scholar] [CrossRef]
- Negi, S.; Pahari, S.; Bashir, H.; Agrewala, J.N. Intestinal microbiota disruption limits the isoniazid mediated clearance of mycobacterium tuberculosis in mice. Eur. J. Immunol. 2020, 50, 1976–1987. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Hou, M.; Gao, T.; Sun, J.; Luo, H.; Wang, F.; Zhong, F.; Ma, A.; Cai, J. Lactobacillus casei improve anti-tuberculosis drugs-induced intestinal adverse reactions in rat by modulating gut microbiota and short-chain fatty acids. Nutrients 2022, 14, 1668. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Study on the Relationship between Intestinal Flora Characteristics and Drug-Induced Liver Injury Induced by Antituberculous Drugs. Ph.D. Thesis, North China University of Science and Technology, Qinhuangdao, China, 2020. [Google Scholar]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via nkt cells. Science 2018, 360, eaan5931. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Wang, K.; Wang, Q.; Jiang, H.; Lu, Y.; Chen, X.; Zhang, H.; Su, X.; Du, Y.; Chen, L.; et al. Probiotic lactobacillus casei shirota prevents acute liver injury by reshaping the gut microbiota to alleviate excessive inflammation and metabolic disorders. Microb. Biotechnol. 2022, 15, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Sun, C.; Yang, J.; Zhang, X.; Dou, S.; Hua, Q.; Ma, A.; Cai, J. Regulation of gut microflora by lactobacillus casei zhang attenuates liver injury in mice caused by anti-tuberculosis drugs. Int. J. Mol. Sci. 2023, 24, 9444. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Chassard, C.; Rohner, F.; N’Goran, E.K.; Nindjin, C.; Dostal, A.; Utzinger, J.; Ghattas, H.; Lacroix, C.; Hurrell, R.F. The effects of iron fortification on the gut microbiota in african children: A randomized controlled trial in cote d’ivoire. Am. J. Clin. Nutr. 2010, 92, 1406–1415. [Google Scholar] [CrossRef]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef]
- Xu, J.; Chen, H.B.; Li, S.L. Understanding the molecular mechanisms of the interplay between herbal medicines and gut microbiota. Med. Res. Rev. 2017, 37, 1140–1185. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-T.; Hu, Y.-Q.; Wang, Y.-L.; Huang, K.; Chen, G.-F.; Zhou, H.; Liu, C.-H.; Yang, T. Antibiotic pretreatment promotes orally-administered triptolide absorption and aggravates hepatotoxicity and intestinal injury in mice. J. Ethnopharmacol. 2022, 292, 115224. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-F.; Zhao, Q.; Dai, M.-Y.; Xiao, X.-R.; Zhang, T.; Zhu, W.-F.; Li, F. Gut microbiota protects from triptolide-induced hepatotoxicity: Key role of propionate and its downstream signalling events. Pharmacol. Res. 2020, 155, 104752. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, X.; Yang, H.; Mao, X.; Wang, J.; Gao, W. Effect of natural β-glucosidase inhibitors in reducing toxicity of amygdalin in persicae semen. Phytother. Res. 2017, 31, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Yu-Feng, Z.; Feng-Rui, S.; Xin-Hua, G.; Shu-Ying, L. Studies on the biotransformation of aconitine in human intestinal bacteria using soft-ionization mass spectrometry. Chem. J. Chin. Univ.-Chin. 2008, 29, 55–59. [Google Scholar]
- Zuo, F.; Zhou, Z.-M.; Yan, M.-Z.; Liu, M.-L.; Xiong, Y.-L.; Zhang, Q.; Song, H.-Y.; Ye, W.-H. Metabolism of constituents in huangqin-tang, a prescription in traditional chinese medicine, by human intestinal flora. Biol. Pharm. Bull. 2002, 25, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H. Gut microbiota-mediated pharmacokinetics of ginseng saponins. J. Ginseng Res. 2018, 42, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ding, Q.; Wang, X.; Li, P.; Fan, N.; Zhou, Y.; Tong, L.; Zhang, W.; Zhang, W.; Tang, B. Visualization of dynamic changes in labile iron(II) pools in endoplasmic reticulum stress-mediated drug-induced liver injury. Anal. Chem. 2020, 92, 1245–1251. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Li, W.; Wang, G.; Fang, S.; Cai, J.; Wang, T.; Zhang, H.; Liu, P.; Wu, J.; Ma, Y. Hepatoprotective effects of huangqi decoction (astragali radix and glycyrrhizae radix et rhizoma) on cholestatic liver injury in mice: Involvement of alleviating intestinal microbiota dysbiosis. J. Ethnopharmacol. 2021, 267, 113544. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, T.; Chen, L.; Zhao, X.; Li, J.; Di, L. Total saponins of glycyrrhiza inflata and its decoction on intestinal flora in rats with liver injury. Chin. Tradit. Herb. Drugs 2020, 51, 101–108. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Zhang, Y.; Wu, K.; Tan, X.; Lan, T.; Wang, G. Role of Gut Microecology in the Pathogenesis of Drug-Induced Liver Injury and Emerging Therapeutic Strategies. Molecules 2024, 29, 2663. https://doi.org/10.3390/molecules29112663
Huang Y, Zhang Y, Wu K, Tan X, Lan T, Wang G. Role of Gut Microecology in the Pathogenesis of Drug-Induced Liver Injury and Emerging Therapeutic Strategies. Molecules. 2024; 29(11):2663. https://doi.org/10.3390/molecules29112663
Chicago/Turabian StyleHuang, Yuqiao, Yu Zhang, Kaireng Wu, Xinxin Tan, Tian Lan, and Guixiang Wang. 2024. "Role of Gut Microecology in the Pathogenesis of Drug-Induced Liver Injury and Emerging Therapeutic Strategies" Molecules 29, no. 11: 2663. https://doi.org/10.3390/molecules29112663
APA StyleHuang, Y., Zhang, Y., Wu, K., Tan, X., Lan, T., & Wang, G. (2024). Role of Gut Microecology in the Pathogenesis of Drug-Induced Liver Injury and Emerging Therapeutic Strategies. Molecules, 29(11), 2663. https://doi.org/10.3390/molecules29112663