
Omics-Based Approaches in Research on Textile Dye Microbial Decolorization

Abstract
:1. Introduction
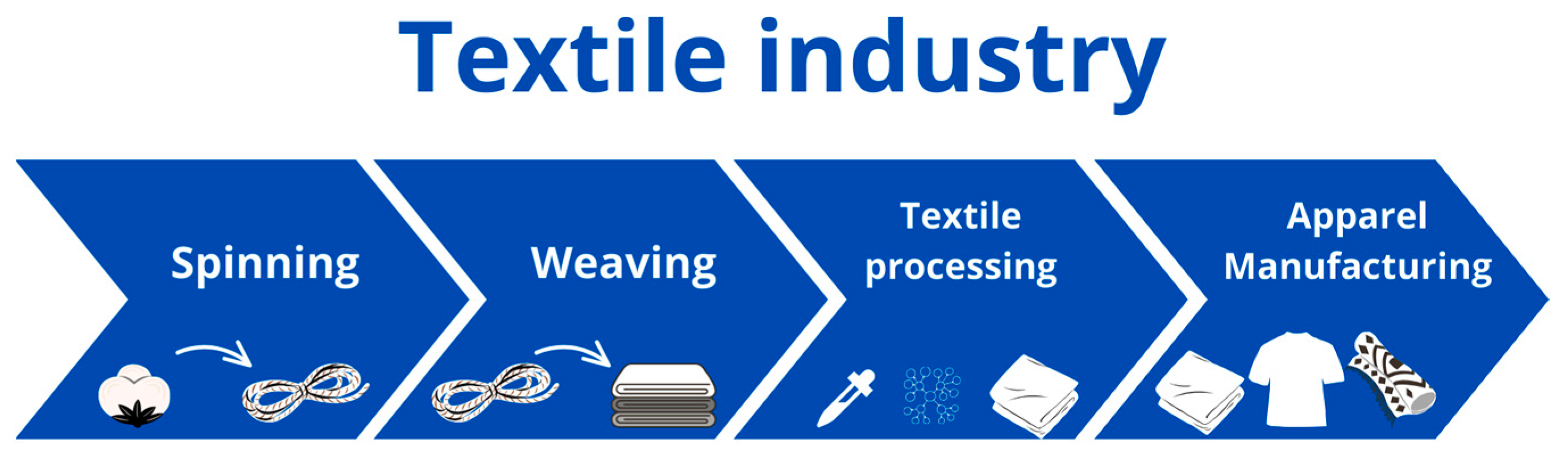
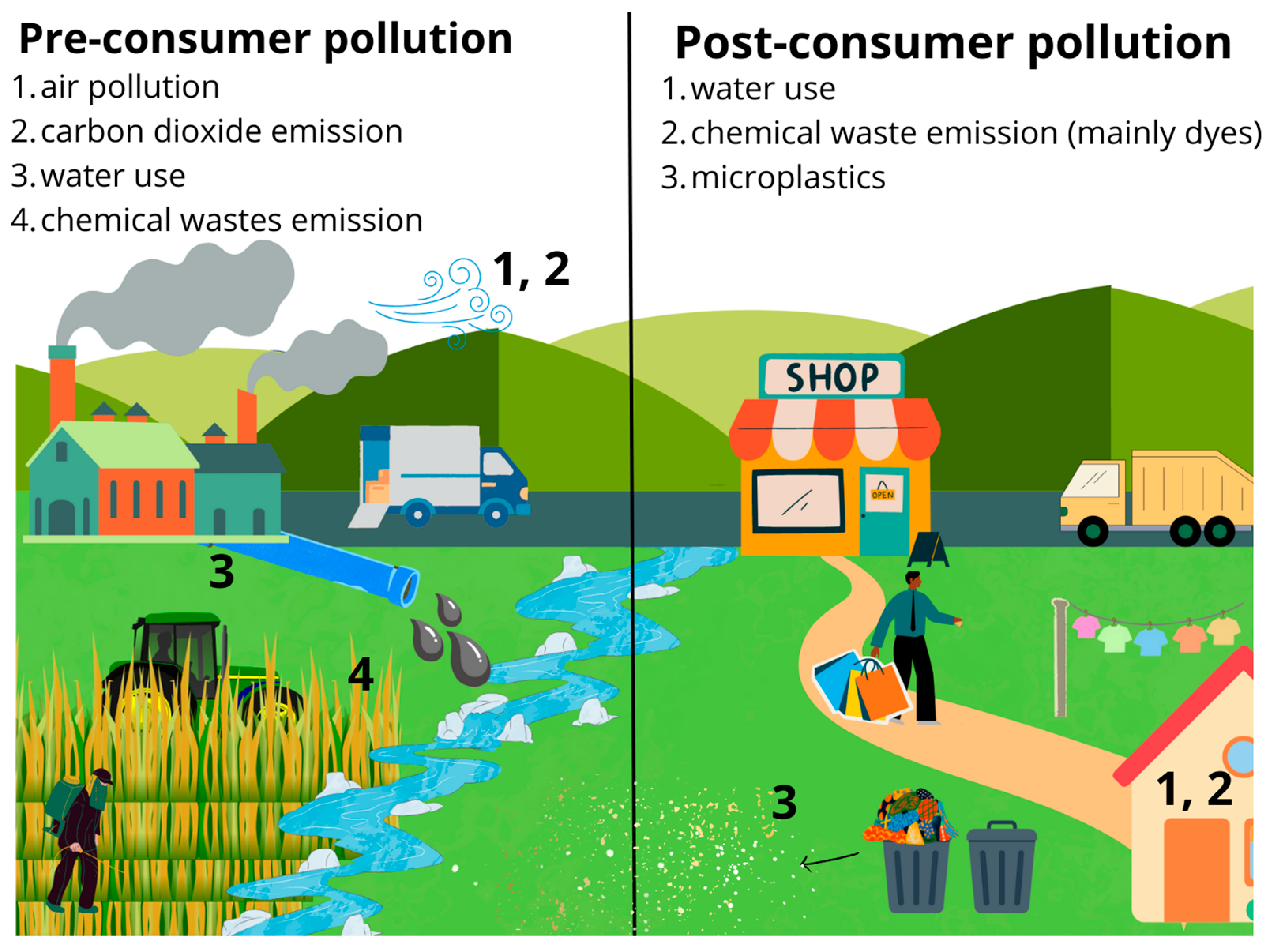
2. Global Textile Industry and Its Environmental Footprints
3. Dyes—Characteristics, Applications, and Danger
4. Non-Biological Treatment Methods for Textile Dye Elimination
5. Microorganisms as a Tool for Textile Dye Elimination
5.1. Conventional Workflow
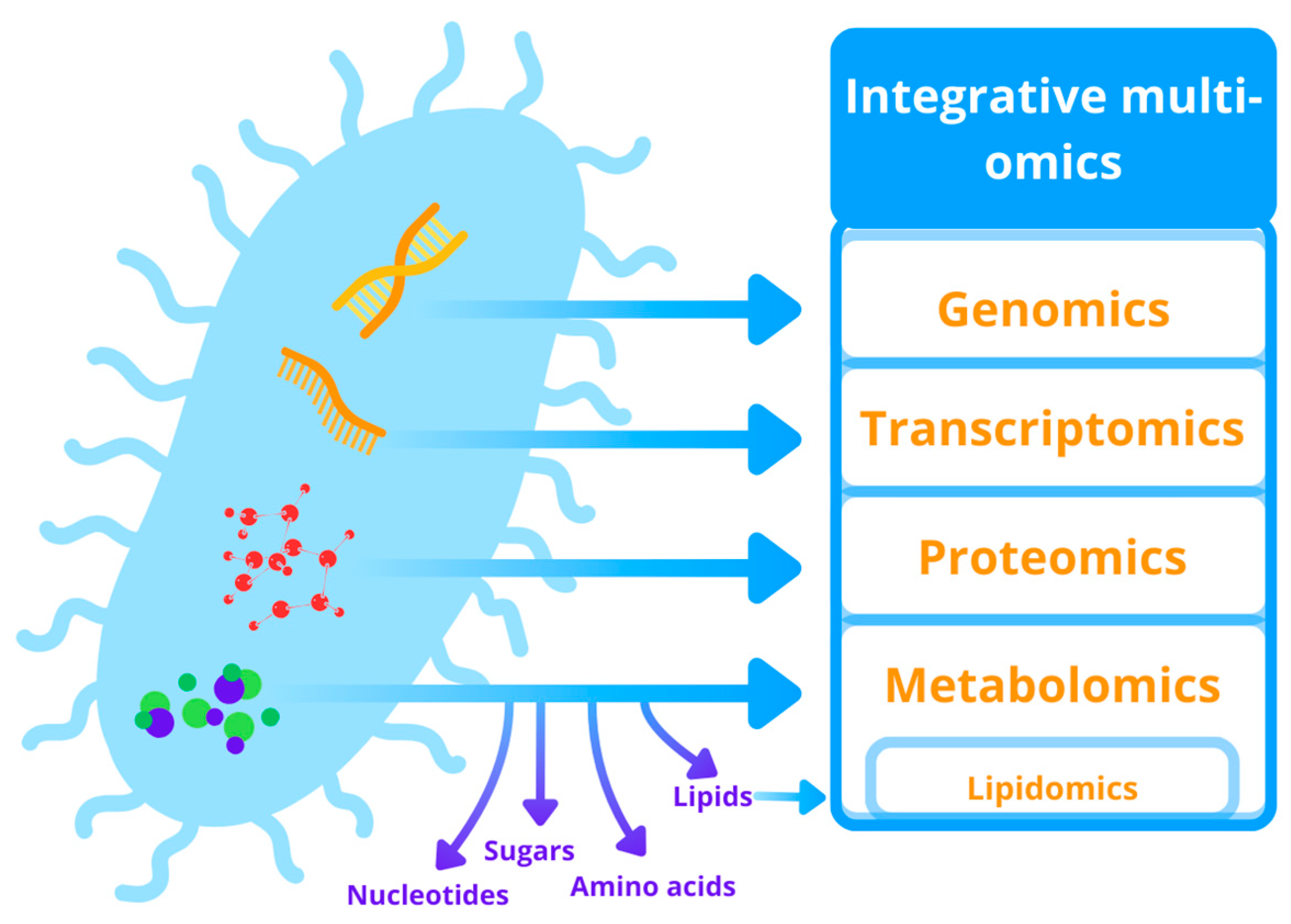
5.2. Omics-Based Approaches in the Dye Biodegradation Research
5.2.1. Genomics and Transcriptomics
5.2.2. Proteomics
5.2.3. Metabolomics
5.2.4. Integrative Omics
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- InkWorld. Global Textile Market to Register CAGR of 4.3% from 2020 to 2027: Grand View Research. 2020. Available online: https://www.inkworldmagazine.com/contents/view_breaking-news/2020-02-19/global-textile-market-to-register-cagr-of-43-from-2020-to-2027-grand-view-research/ (accessed on 2 June 2024).
- Costa, C.; Azoia, N.; Silva, C.; Marques, E. Textile Industry in a Changing World. U Porto J. Eng. 2020, 6, 86–97. [Google Scholar] [CrossRef]
- Filho, W.L.; Perry, P.; Heim, H.; Dinis, M.A.P.; Moda, H.; Ebhuoma, E.; Paço, A. An Overview of the Contribution of the Textiles Sector to Climate Change. Front. Environ. Sci. 2022, 10, 973102. [Google Scholar] [CrossRef]
- Plakantonaki, S.; Kiskira, K.; Zacharopoulos, N.; Chronis, I.; Coelho, F.; Togiani, A.; Kalkanis, K.; Priniotakis, G. A Review of Sustainability Standards and Ecolabeling in the Textile Industry. Sustainability 2023, 15, 11589. [Google Scholar] [CrossRef]
- Koussé, J.N.D.; Ilboudo, S.; Ouédraogo, J.C.R.P.; Hunsmann, M.; Ouédraogo, G.G.; Ouédraogo, M.; Kini, F.B.; Ouédraogo, S. Self-Reported Health Effects of Pesticides among Cotton Farmers from the Central-West Region in Burkina Faso. Toxicol. Rep. 2023, 11, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Memon, Q.U.A.; Wagan, S.A.; Chunyu, D.; Shuangxi, X.; Jingdong, L.; Damalas, C.A. Health Problems from Pesticide Exposure and Personal Protective Measures among Women Cotton Workers in Southern Pakistan. Sci. Total Environ. 2019, 685, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Douny, C.; Zoumenou, Y.M.B.G.; Aïna, M.; Toko, I.I.; Igout, A.; Guedegba, L.; Chabi, S.K.; Kestemont, P.; Scippo, M.-L. Contamination of Water, Sediment and Fish with Residues of Pesticides Used in Cotton Production in Northern Benin. Arch. Environ. Contam. Toxicol. 2021, 81, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wu, C.; Liu, W.; Teng, Y.; Luo, Y.; Christie, P.; Guo, D. Levels and Patterns of Organochlorine Pesticides in Agricultural Soils in an Area of Extensive Historical Cotton Cultivation in Henan Province, China. Environ. Sci. Pollut. Res. 2015, 23, 6680–6689. [Google Scholar] [CrossRef] [PubMed]
- Rafique, N.; Tariq, S.R.; Ahmed, D. Monitoring and Distribution Patterns of Pesticide Residues in Soil from Cotton/Wheat Fields of Pakistan. Environ. Monit. Assess. 2016, 188, 695. [Google Scholar] [CrossRef] [PubMed]
- Rajmohan, K.S.; Chandrasekaran, R.; Varjani, S. A Review on Occurrence of Pesticides in Environment and Current Technologies for Their Remediation and Management. Indian J. Microbiol. 2020, 60, 125–138. [Google Scholar] [CrossRef]
- Ellen McArthur Foundation; Morlet, A.; Opsomer, R.; Herrmann, S.; Balmond, L.; Gillet, C.; Fuchs, L. A New Textiles Economy: Redesigning Fashion’s Future; Ellen MacArthur Foundation: Cowes, UK, 2017; Available online: https://www.ellenmacarthurfoundation.org/a-new-textiles-economy (accessed on 2 June 2024).
- Lara, L.; Cabral, I.; Cunha, J. Ecological Approaches to Textile Dyeing: A Review. Sustainability 2022, 14, 8353. [Google Scholar] [CrossRef]
- Benkhaya, S.; M’rabet, S.; El Harfi, A. A Review on Classifications, Recent Synthesis and Applications of Textile Dyes. Inorg. Chem. Commun. 2020, 115, 107891. [Google Scholar] [CrossRef]
- Sarkar, S.; Chakraborty, P.; Bandopadhyay, R. Microbial Treatment for Removing Synthetic Dyes from Industrial Effluents; Springer Nature: Singapore, 2020; pp. 47–63. [Google Scholar] [CrossRef]
- Chung, K.-T. Azo Dyes and Human Health: A Review. J. Environ. Sci. Health Part C 2016, 34, 233–261. [Google Scholar] [CrossRef] [PubMed]
- Egan, J.; Salmon, S. Strategies and Progress in Synthetic Textile Fiber Biodegradability. SN Appl. Sci. 2021, 4, 22. [Google Scholar] [CrossRef]
- Periyasamy, A.P.; Tehrani-Bagha, A. A Review on Microplastic Emission from Textile Materials and Its Reduction Techniques. Polym. Degrad. Stab. 2022, 199, 109901. [Google Scholar] [CrossRef]
- Salvador Cesa, F.; Turra, A.; Baruque-Ramos, J. Synthetic Fibers as Microplastics in the Marine Environment: A Review from Textile Perspective with a Focus on Domestic Washings. Sci. Total Environ. 2017, 598, 1116–1129. [Google Scholar] [CrossRef]
- Bayo, J.; Olmos, S.; López-Castellanos, J.; Alcolea, A. Microplastics and Microfibers in the Sludge of a Municipal Wastewater Treatment Plant. Int. J. Sustain. Dev. 2016, 11, 812–821. [Google Scholar] [CrossRef]
- Bhatia, D.; Sharma, N.R.; Singh, J.; Kanwar, R.S. Biological Methods for Textile Dye Removal from Wastewater: A Review. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1836–1876. [Google Scholar] [CrossRef]
- Holkar, C.R.; Jadhav, A.J.; Pinjari, D.V.; Mahamuni, N.M.; Pandit, A.B. A Critical Review on Textile Wastewater Treatments: Possible Approaches. J. Environ. Manag. 2016, 182, 351–366. [Google Scholar] [CrossRef]
- Azanaw, A.; Birlie, B.; Teshome, B.; Jemberie, M. Textile Effluent Treatment Methods and Eco-Friendly Resolution of Textile Wastewater. Case Stud. Chem. Environ. Eng. 2022, 6, 100230. [Google Scholar] [CrossRef]
- Roy, M.; Saha, R. Dyes and Their Removal Technologies from Wastewater: A Critical Review. In Intelligent Environmental Data Monitoring for Pollution Management; Bhattacharyya, S., Mondal, N.K., Platos, J., Snášel, V., Krömer, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 127–160. [Google Scholar]
- Patti, A.; Cicala, G.; Acierno, D. Eco-Sustainability of the Textile Production: Waste Recovery and Current Recycling in the Composites World. Polymers 2020, 13, 134. [Google Scholar] [CrossRef]
- El Nemr, A. From Natural to Synthetic Fibers. In Textiles: Types, Uses and Production Methods; El Nemr, A., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2012. [Google Scholar]
- Reaserch and Markets. Textile Market Size, Share & Trends Analysis Report by Raw Material (Wool, Chemical, Silk), by Product (Natural Fibers, Polyester), by Application, by Region, and Segment Forecasts, 2023–2030; Reaserch and Markets: Dublin, Ireland, 2023. [Google Scholar]
- Nikolina, Š. Environmental Impact of the Textile and Clothing Industry. What Consumers Need to Know. Eur. Parliam. Res. Serv. 2019. Available online: https://www.europarl.europa.eu/RegData/etudes/BRIE/2019/633143/EPRS_BRI(2019)633143_EN.pdf (accessed on 16 February 2024).
- Degenhard, J. Total Consumer Spending on Clothing and Footwear Worldwide from 2014 to 2029. Available online: https://www.statista.com/forecasts/1161735/fashion-consumer-spending-forecast-in-the-world (accessed on 16 February 2024).
- Caro, F.; Martínez-de-Albéniz, V. Fast Fashion: Business Model Overview and Research Opportunities. Retail Supply Chain Manag. 2015, 223, 237–264. [Google Scholar] [CrossRef]
- Niinimäki, K.; Peters, G.; Dahlbo, H.; Perry, P.; Rissanen, T.; Gwilt, A. The Environmental Price of Fast Fashion. Nat. Rev. Earth Environ. 2020, 1, 189–200. [Google Scholar] [CrossRef]
- Kumar, P.S.; Prasanth, S.M.; Harish, S.; Rishikesh, M. Rishikesh Industrial Water Footprint: Case Study on Textile Industries. In Environmental Footprints and Eco-Design of Products and Processes; Muthu, S.S., Ed.; Springer: Singapore, 2021; pp. 35–60. [Google Scholar]
- EJF. The Deadly Chemicals in Cotton; Environmental Justice Foundation in Collaboration with Pesticide Action Network UK: London, UK, 2007; ISBN 1-904523-10-2. [Google Scholar]
- Khan, S.; Malik, A. Environmental and Health Effects of Textile Industry Wastewater. In Environmental Deterioration and Human Health; Malik, A., Grohmann, E., Akhtar, R., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 55–71. ISBN 9789400778894. [Google Scholar]
- Uddin, M.A.; Begum, M.S.; Ashraf, M.; Azad, A.K.; Adhikary, A.C.; Hossain, M.S. Water and Chemical Consumption in the Textile Processing Industry of Bangladesh. PLOS Sustain. Transform. 2023, 2, e0000072. [Google Scholar] [CrossRef]
- Vercalsteren, A.; Nicolau, M.; Lafond, E. Textiles and the Environment in a Circular Economy. Eionet Report 2019-ETC/WMGE2019/6. Available online: https://www.eionet.europa.eu/etcs/etc-wmge/products/etc-wmge-reports/textiles-and-the-environment-in-a-circular-economy (accessed on 16 February 2024).
- Ngo, A.C.R.; Tischler, D. Microbial Degradation of Azo Dyes: Approaches and Prospects for a Hazard-Free Conversion by Microorganisms. Int. J. Environ. Res. Public Health 2022, 19, 4740. [Google Scholar] [CrossRef] [PubMed]
- Manshoven, S.; Grossi, F. Textiles and the Environment the Role of Design in Europe’s Circular Economy. 2022. Available online: https://www.cscp.org/wp-content/uploads/2022/03/ETC_Design-of-Textiles.pdf (accessed on 2 June 2024).
- Papamichael, I.; Chatziparaskeva, G.; Pedreno, J.N.; Voukkali, I.; Almendro Candel, M.B.; Zorpas, A.A. Building a New Mind Set in Tomorrow Fashion Development through Circular Strategy Models in the Framework of Waste Management. Curr. Opin. Green Sustain. Chem. 2022, 36, 100638. [Google Scholar] [CrossRef]
- Volgare, M.; De Falco, F.; Avolio, R.; Castaldo, R.; Errico, M.E.; Gentile, G.; Ambrogi, V.; Cocca, M. Washing Load Influences the Microplastic Release from Polyester Fabrics by Affecting Wettability and Mechanical Stress. Sci. Rep. 2021, 11, 19479. [Google Scholar] [CrossRef]
- Sillanpää, M.; Sainio, P. Release of Polyester and Cotton Fibers from Textiles in Machine Washings. Environ. Sci. Pollut. Res. 2017, 24, 19313–19321. [Google Scholar] [CrossRef] [PubMed]
- Synthetic Dyes Market by Type (Acid Dyes, Reactive Dyes, Disperse Dyes, Direct Dyes, Solvent Dyes, Basic Dyes), Application, Form (Liquid, Powder), Structure (Anionic, Cationic, Non-Lonic), End-Use Industry, and Region-Global Forecast to 2027. Available online: https://www.marketsandmarkets.com/Market-Reports/synthetic-dyes-market-184747754.html (accessed on 16 February 2024).
- Benkhaya, S.; M’rabet, S.; Lgaz, H.; El Bachiri, A.; El Harfi, A. Dyes: Classification, Pollution, and Environmental Effects. In Dye Biodegradation, Mechanisms and Techniques; Senthilkannan Muthu, S., Khadir, A., Eds.; Springer: Singapore, 2021; pp. 1–50. [Google Scholar] [CrossRef]
- Bafana, A.; Devi, S.S.; Chakrabarti, T. Azo Dyes: Past, Present and the Future. Environ. Rev. 2011, 19, 350–371. [Google Scholar] [CrossRef]
- Fleischmann, C.; Lievenbrück, M.; Ritter, H. Polymers and Dyes: Developments and Applications. Polymers 2015, 7, 717–746. [Google Scholar] [CrossRef]
- Tkaczyk, A.; Mitrowska, K.; Posyniak, A. Synthetic Organic Dyes as Contaminants of the Aquatic Environment and Their Implications for Ecosystems: A Review. Sci. Total Environ. 2020, 717, 137222. [Google Scholar] [CrossRef]
- Topaç, F.O.; Dindar, E.; Uçaroğlu, S.; Başkaya, H.S. Effect of a Sulfonated Azo Dye and Sulfanilic Acid on Nitrogen Transformation Processes in Soil. J. Hazard. Mater. 2009, 170, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Stingley, R.L.; Zou, W.; Heinze, T.M.; Chen, H.; Cerniglia, C.E. Metabolism of Azo Dyes by Human Skin Microbiota. J. Med. Microbiol. 2010, 59, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.; Mahmoud, S.; Elgawish, R.; Elhady, K.A. Sudan III Azo Dye: Oxidative Stress with Possible Geno and Hepatotoxic Effects in Male Rats. Int. J. Sci. Res. 2016, 5, 1700–1704. [Google Scholar] [CrossRef]
- Bhatt, D.; Vyas, K.; Singh, S.; John, P.J.; Soni, I.P. Sunset Yellow Induced Biochemical and Histopathological Alterations in Rat Brain Sub-Regions. Acta Histochem. 2024, 126, 152155. [Google Scholar] [CrossRef] [PubMed]
- Brüschweiler, B.J.; Küng, S.; Bürgi, D.; Muralt, L.; Nyfeler, E. Identification of Non-Regulated Aromatic Amines of Toxicological Concern Which Can Be Cleaved from Azo Dyes Used in Clothing Textiles. Regul. Toxicol. Pharmacol. 2014, 69, 263–272. [Google Scholar] [CrossRef]
- Brüschweiler, B.J.; Merlot, C. Azo Dyes in Clothing Textiles Can Be Cleaved into a Series of Mutagenic Aromatic Amines Which Are Not Regulated Yet. Regul. Toxicol. Pharmacol. 2017, 88, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Bazin, I.; Ibn Hadj Hassine, A.; Haj Hamouda, Y.; Mnif, W.; Bartegi, A.; Lopez-Ferber, M.; De Waard, M.; Gonzalez, C. Estrogenic and Anti-Estrogenic Activity of 23 Commercial Textile Dyes. Ecotoxicol. Environ. Saf. 2012, 85, 131–136. [Google Scholar] [CrossRef]
- World Water Assessment Programme (Nations Unies), The United Nations World Water Development Report 2018 (United Nations Educational, Scientific and Cultural Organization, New York, United States). Available online: https://www.unwater.org/publications/world-water-development-report-2018 (accessed on 16 February 2024).
- Jasińska, A.; Różalska, S.; Bernat, P.; Paraszkiewicz, K.; Długoński, J. Malachite Green Decolorization by Non-Basidiomycete Filamentous Fungi of Penicillium Pinophilum and Myrothecium Roridum. Int. Biodeterior. Biodegrad. 2012, 73, 33–40. [Google Scholar] [CrossRef]
- Modi, H.A.; Rajput, G.; Ambasana, C. Decolorization of Water Soluble Azo Dyes by Bacterial Cultures, Isolated from Dye House Effluent. Bioresour. Technol. 2010, 101, 6580–6583. [Google Scholar] [CrossRef]
- Krishnan, J.; Arvind Kishore, A.; Suresh, A.; Madhumeetha, B.; Gnana Prakash, D. Effect of PH, Inoculum Dose and Initial Dye Concentration on the Removal of Azo Dye Mixture under Aerobic Conditions. Int. Biodeterior. Biodegrad. 2017, 119, 16–27. [Google Scholar] [CrossRef]
- Seyedi, Z.S.; Zahraei, Z.; Jookar Kashi, F. Decolorization of Reactive Black 5 and Reactive Red 152 Azo Dyes by New Haloalkaliphilic Bacteria Isolated from the Textile Wastewater. Curr. Microbiol. 2020, 77, 2084–2092. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Sadasivam, S.K. Biodegradation of Textile Azo Dyes by Textile Effluent Non-Adapted and Adapted Aeromonas Hydrophila. Environ. Res. J. 2021, 194, 110643. [Google Scholar] [CrossRef] [PubMed]
- Herath, I.S.; Udayanga, D.; Jayasanka, D.J.; Hewawasam, C. Textile Dye Decolorization by White Rot Fungi—A Review. Bioresour. Technol. Rep. 2024, 25, 101687. [Google Scholar] [CrossRef]
- Yadav, A.; Yadav, P.; Kumar Singh, A.; Kumar, V.; Chintaman Sonawane, V.; Markandeya; Naresh Bharagava, R.; Raj, A. Decolourisation of Textile Dye by Laccase: Process Evaluation and Assessment of Its Degradation Bioproducts. Bioresour. Technol. 2021, 340, 125591. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chen, Y.; Guan, J.; Ding, Y.; He, Y.; Zhang, X.; Shukurov, N.; Romanholo, F.; Liu, J.; Zhu, M. Biodecolorization and Ecotoxicity Abatement of Disperse Dye-Production Wastewater Treatment with Pycnoporus Laccase. Int. J. Environ. Res. Public Health 2022, 19, 7983. [Google Scholar] [CrossRef]
- Pandi, A.; Marichetti Kuppuswami, G.; Numbi Ramudu, K.; Palanivel, S. A Sustainable Approach for Degradation of Leather Dyes by a New Fungal Laccase. J. Clean. Prod. 2019, 211, 590–597. [Google Scholar] [CrossRef]
- Sugano, Y.; Yoshida, T. DyP-Type Peroxidases: Recent Advances and Perspectives. Int. J. Mol. Sci. 2021, 22, 5556. [Google Scholar] [CrossRef] [PubMed]
- Misal, S.A.; Gawai, K.R. Azoreductase: A Key Player of Xenobiotic Metabolism. Bioresour. Bioprocess 2018, 5, 17. [Google Scholar] [CrossRef]
- Jasińska, A.; Paraszkiewicz, K.; Słaba, M.; Długoński, J. Microbial Decolorization of Triphenylmethane Dyes. In Microbial Degradation of Synthetic Dyes in Wastewaters. Environmental Science and Engineering; Springer: Cham, Switzerland, 2014; pp. 169–186. [Google Scholar] [CrossRef]
- Yu, J.; Lai, J.; Neal, B.M.; White, B.J.; Banik, M.T.; Dai, S.Y. Genomic Diversity and Phenotypic Variation in Fungal Decomposers Involved in Bioremediation of Persistent Organic Pollutants. J. Fungi 2023, 9, 418. [Google Scholar] [CrossRef]
- Yang, P.; Shi, W.; Wang, H.; Liu, H. Screening of Freshwater Fungi for Decolorizing Multiple Synthetic Dyes. Braz. J. Microbiol. 2016, 47, 828–834. [Google Scholar] [CrossRef]
- Roy, U.; Sengupta, S.; Banerjee, P.; Das, P.; Bhowal, A.; Datta, S. Assessment on the Decolourization of Textile Dye (Reactive Yellow) Using Pseudomonas Sp. Immobilized on Fly Ash: Response Surface Methodology Optimization and Toxicity Evaluation. J. Environ. Manag. 2018, 223, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Kıvanç, M.R.; Yönten, V. A statistical optimization of methylene blue removal from aqueous solutions by Agaricus Campestris using multi-step experimental design with response surface methodology: Isotherm, kinetic and thermodynamic studies. Surf. Interfaces 2020, 18, 100414. [Google Scholar] [CrossRef]
- Singh, G.; Dwivedi, S.K. Decolorization and Degradation of Direct Blue-1 (Azo Dye) by Newly Isolated Fungus Aspergillus terreus GS28, from Sludge of Carpet Industry. Environ. Technol. Innov. 2020, 18, 100751. [Google Scholar] [CrossRef]
- Góralczyk-Bińkowska, A.; Długoński, A.; Bernat, P.; Długoński, J.; Jasińska, A. Environmental and Molecular Approach to Dye Industry Waste Degradation by the Ascomycete Fungus Nectriella pironii. Sci. Rep. 2021, 11, 23829. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, F.; Li, J.; Li, D.-B.; Liu, D.; Li, W.; Yu, H. Exclusive Extracellular Bioreduction of Methyl Orange by Azo Reductase-Free Geobacter Sulfurreducens. Environ. Sci. Technol. 2017, 51, 8616–8623. [Google Scholar] [CrossRef]
- Saratale, R.G.; Saratale, G.D.; Chang, J.S.; Govindwar, S.P. Bacterial Decolorization and Degradation of Azo Dyes: A Review. J. Taiwan Inst. Chem. Eng. 2011, 42, 138–157. [Google Scholar] [CrossRef]
- Danouche, M.; EL Arroussi, H.; El Ghachtouli, N. Mycoremediation of Synthetic Dyes by Yeast Cells: A Sustainable Biodegradation Approach. Environ. Sustain. 2021, 4, 5–22. [Google Scholar] [CrossRef]
- Kishor, R.; Purchase, D.; Saratale, G.D.; Saratale, R.G.; Ferreira, L.F.R.; Bilal, M.; Chandra, R.; Bharagava, R.N. Ecotoxicological and Health Concerns of Persistent Coloring Pollutants of Textile Industry Wastewater and Treatment Approaches for Environmental Safety. J. Environ. Chem. Eng. 2021, 9, 105012. [Google Scholar] [CrossRef]
- Rawat, D.; Sharma, R.S.; Karmakar, S.; Arora, L.S.; Mishra, V. Ecotoxic Potential of a Presumably Non-Toxic Azo Dye. Ecotoxicol. Environ. Saf. 2018, 148, 528–537. [Google Scholar] [CrossRef]
- Almeida, E.J.R.; Corso, C.R. Decolorization and Removal of Toxicity of Textile Azo Dyes Using Fungal Biomass Pelletized. Int. J. Environ. Sci. Technol. 2018, 16, 1319–1328. [Google Scholar] [CrossRef]
- Ceretta, M.B.; Nercessian, D.; Wolski, E.A. Current Trends on Role of Biological Treatment in Integrated Treatment Technologies of Textile Wastewater. Front. Microbiol. 2021, 12, 651025. [Google Scholar] [CrossRef] [PubMed]
- Malla, M.A.; Dubey, A.; Yadav, S.; Kumar, A.; Hashem, A.; Abd Allah, E.F. Understanding and Designing the Strategies for the Microbe-Mediated Remediation of Environmental Contaminants Using Omics Approaches. Front. Microbiol. 2018, 9, 1132. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, N.; Bansal, M.; Santhiya, D.; Sharma, J.G. Insights into Microbial Diversity on Plastisphere by Multi-Omics. Arch. Microbiol. 2022, 204, 216. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, S.; Ngo, A.C.R.; Schultes, F.P.J.; Tischler, D. Draft Genome Sequence of Kocuria indica DP-K7, a Methyl Red Degrading Actinobacterium. 3 Biotech 2020, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, I.; Ccorahua, R.; Tinoco, O.; León, O.; Ramírez, P. Draft Genome Sequences of Two Textile Azo Dye-Degrading Shewanella Sp. Strains Isolated from a Textile Effluent in Peru. Microbiol. Resour. Announc. 2019, 8, e00836-19. [Google Scholar] [CrossRef] [PubMed]
- Faldu, P.R.; Kothari, V.V.; Kothari, C.R.; Rawal, C.M.; Domadia, K.K.; Patel, P.A.; Bhimani, H.D.; Raval, V.H.; Parmar, N.R.; Nathani, N.M.; et al. Draft Genome Sequence of Textile Azo Dye-Decolorizing and -Degrading Pseudomonas aeruginosa Strain PFK10, Isolated from the Common Effluent Treatment Plant of the Ankleshwar Industrial Area of Gujarat, India. Genome Announc. 2014, 2, e00019-14. [Google Scholar] [CrossRef]
- Li, Y.; Ng, I.-S.; Zhang, X.; Wang, N. Draft Genome Sequence of the Dye-Decolorizing and Nanowire-Producing Bacterium Shewanella Xiamenensis BC01. Genome Announc. 2014, 2, e00721-14. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Ng, I.-S.; Chen, P.T.; Li, Y.; Chen, Y.-C.; Chen, B.-Y.; Lu, Y. Draft Genome Sequence of the Bioelectricity-Generating and Dye-Decolorizing Bacterium Proteus hauseri Strain ZMd44. Genome Announc. 2014, 2, e00992-13. [Google Scholar] [CrossRef]
- Nsa, I.Y.; Odunsi, A.A.; Amund, O.O. Draft Genome Sequence and Annotation of the Synthetic Textile Dye-Decolorizing Strain Bacillus amyloliquefaciens AD20, Isolated from a Dye Waste Pond. Microbiol. Resour. Announc. 2021, 10, e00645-21. [Google Scholar] [CrossRef]
- Kunadia, K.; Nathani, N.M.; Kothari, V.; Kotadia, R.J.; Kothari, C.R.; Joshi, A.; Rank, J.K.; Faldu, P.R.; Shekar, M.C.; Viroja, M.J.; et al. Draft Genome Sequence of Commercial Textile Dye-Decolorizing and -Degrading Bacillus subtilis Strain C3 Isolated in India. Genome Announc. 2016, 4, e00104-16. [Google Scholar] [CrossRef]
- Nsa, I.Y.; Omolere, B.M. Draft Genome Sequence and Annotation of Priestia aryabhattai Strain BD1 Isolated from a Dye Sediment. Microbiol. Resour. Announc. 2023, 12, e01175-22. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Wang, Y.; Yang, G.; Luo, H.; Zhuang, L.; Yu, Z.; Zhou, S. Complete Genome Sequence of the Dissimilatory Azo Reducing Thermophilic Bacterium Novibacillus thermophiles SG-1. J. Biotechnol. 2018, 284, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; An, X.; Feng, L.; Xia, X.; Zhang, Q. Genome and Transcriptome Analysis of a Newly Isolated Azo Dye Degrading Thermophilic Strain Anoxybacillus Sp. Ecotoxicol. Environ. Saf. 2020, 203, 111047. [Google Scholar] [CrossRef] [PubMed]
- Lizárraga, W.C.; Mormontoy, C.G.; Calla, H.; Castañeda, M.; Taira, M.; Garcia, R.; Marín, C.; Abanto, M.; Ramirez, P. Complete Genome Sequence of Shewanella Algae Strain 2NE11, a Decolorizing Bacterium Isolated from Industrial Effluent in Peru. Biotechnol. Rep. 2022, 33, e00704. [Google Scholar] [CrossRef] [PubMed]
- Nanjani, S.; Paul, D.; Keharia, H. Genome Analysis to Decipher Syntrophy in the Bacterial Consortium “SCP” for Azo Dye Degradation. BMC Microbiol. 2021, 21, 177. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Wang, Y.; Guo, G.; Ding, K.; Yang, F.; Wang, C.; Wang, H.; Yan, M. Meta-Genome Analysis of a Newly Enriched Azo Dyes Detoxification Halo-Thermophilic Bacterial Consortium. Environ. Res. J. 2023, 237, 116828. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Liu, Q.; Ma, J.; Liao, H.; Xiong, X.; Zhang, K.-K.; Wang, T.; Liu, X.; Xu, T.; Yuan, S.; et al. An Acid-Stable Bacterial Laccase Identified from the Endophyte Pantoea ananatis Sd-1 Genome Exhibiting Lignin Degradation and Dye Decolorization Abilities. Biotechnol. Lett. 2015, 37, 2279–2288. [Google Scholar] [CrossRef]
- Qu, W.; Hong, G.-L.; Zhao, J. Degradation of Malachite Green Dye by Tenacibaculum Sp. HMG1 Isolated from Pacific Deep-Sea Sediments. Acta Oceanol. Sin. 2018, 37, 104–111. [Google Scholar] [CrossRef]
- Pei, S.; Niu, S.; Xie, F.; Wang, W.; Zhang, S.; Zhang, G. Brevibacterium limosum Sp. Nov., Brevibacterium pigmenatum Sp. Nov., and Brevibacterium atlanticum Sp. Nov., Three Novel Dye Decolorizing Actinobacteria Isolated from Ocean Sediments. J. Microbiol. 2021, 59, 898–910. [Google Scholar] [CrossRef]
- Zhang, C.; Xin, Y.; Wang, Y.; Guo, T.; Lu, S.; Kong, J. Identification of a Novel Dye-Decolorizing Peroxidase, EfeB, Translocated by a Twin-Arginine Translocation System in Streptococcus Thermophilus CGMCC 7.179. Appl. Environ. Microbiol. 2015, 81, 6108–6119. [Google Scholar] [CrossRef]
- Alghamdi, M.A.; Ayed, L.; Aljarad, M.R.; Altayeb, H.N.; Abbes, S.; Chaieb, K. Whole Genome Sequencing Analysis and Box-Behnken Design for the Optimization of the Decolourization of Mixture Textile Dyes by Halotolerant Microbial Consortium. Microbiol. Res. 2023, 276, 127481. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.U.; Hinsu, A.T.; Kotadiya, R.J.; Rank, J.K.; Andharia, K.N.; Kothari, R.K. Decolorization and Biodegradation of Textile Di-Azo Dye Acid Blue 113 by Pseudomonas stutzeri AK6. 3 Biotech 2020, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- John, J.; Dineshram, R.; Hemalatha, K.R.; Dhassiah, M.P.; Gopal, D.; Kumar, A. Bio-Decolorization of Synthetic Dyes by a Halophilic Bacterium Salinivibrio Sp. Front. Microbiol. 2020, 11, 594011. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; Park, H.; Jang, Y.; Kim, J.-J.; Kim, K.H.; Pangilinan, J.; Lipzen, A.; Riley, R.; Grigoriev, I.V.; Spatafora, J.W.; et al. Genome Sequence of a White Rot Fungus Schizopora paradoxa KUC8140 for Wood Decay and Mycoremediation. J. Biotechnol. 2015, 211, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-W.; Nam, J.; Nguyen, H.T.K.; Lee, J.; Choi, Y.; Choi, J. Draft Genome Sequence of the White-Rot Fungus Schizophyllum commune IUM1114-SS01. J. Mycobiol. 2020, 49, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Li, G.; Liang, W.Q.; Liu, Y.H. Molecular Cloning and Characterization of a Novel Metagenome-Derived Multicopper Oxidase with Alkaline Laccase Activity and Highly Soluble Expression. Appl. Microbiol. Biotechnol. 2010, 87, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.H.; Ye, M.; Lu, Y.; Zhang, X.; Li, G. Improving the Decolorization for Textile Dyes of a Metagenome-Derived Alkaline Laccase by Directed Evolution. Appl. Microbiol. Biotechnol. 2011, 91, 667–675. [Google Scholar] [CrossRef]
- Sun, S.; Xie, S.; Cheng, Y.; Yu, H.; Zhao, H.; Li, M.; Li, X.; Zhang, X.; Yuan, J.S.; Dai, S.Y. Enhancement of Environmental Hazard Degradation in the Presence of Lignin: A Proteomics Study. Sci. Rep. 2017, 7, 11356. [Google Scholar] [CrossRef]
- Zhang, Q.; Xie, X.; Liu, Y.; Zheng, X.; Wang, Y.; Cong, J.; Yu, C.; Liu, N.; Sand, W.; Liu, J. Co-Metabolic Degradation of Refractory Dye: A Metagenomic and Metaproteomic Study. Environ. Pollut. 2020, 256, 113456. [Google Scholar] [CrossRef]
- Guo, G.; Liu, C.; Hao, J.; Tian, F.; Ding, K.; Zhang, C.; Yang, F.; Liu, T.; Xu, J.; Guan, Z. Development and Characterization of a Halo-Thermophilic Bacterial Consortium for Decolorization of Azo Dye. Chemosphere 2021, 272, 129916. [Google Scholar] [CrossRef]
- Zhuang, M.; Sanganyado, E.; Xu, L.; Zhu, J.; Li, P.; Liu, W. High Throughput Sediment DNA Sequencing Reveals Azo Dye Degrading Bacteria Inhabit Nearshore Sediments. Microorganisms 2020, 8, 233. [Google Scholar] [CrossRef]
- Tan, L.; Qu, Y.; Zhou, J.; Ma, F.; Li, A. Dynamics of Microbial Community for X-3B Wastewater Decolorization Coping with High-Salt and Metal Ions Conditions. Bioresour. Technol. 2009, 100, 3003–3009. [Google Scholar] [CrossRef] [PubMed]
- Copete, L.S.; Chanagá, X.; Barriuso, J.; López-Lucendo, M.F.; Martínez, M.J.; Camarero, S. Identification and Characterization of Laccase-Type Multicopper Oxidases Involved in Dye-Decolorization by the Fungus Leptosphaerulina Sp. BMC Biotechnol. 2015, 15, 74. [Google Scholar] [CrossRef] [PubMed]
- de Souza, N.A.; Ramaiah, N.; Damare, S.R.; Furtado, B.; Mohandass, C.; Patil, A.; De Lima, M. Differential Protein Expression in Shewanella seohaensis Decolorizing Azo Dyes. Curr. Proteom. 2019, 16, 156–164. [Google Scholar] [CrossRef]
- Mathur, A.; Dubey, S.; Prasad, R.; Singh, R.P. Mycelial and Secretome Proteomic Dynamics of L. squarrosulus AF5 in Azo Dye Degradation. J. Environ. Chem. Eng. 2023, 11, 109374. [Google Scholar] [CrossRef]
- Jasińska, A.; Soboń, A.; Góralczyk-Bińkowska, A.; Długoński, J. Analysis of Decolorization Potential of Myrothecium roridum in the Light of Its Secretome and Toxicological Studies. Environ. Sci. Pollut. Res. 2019, 26, 26313–26323. [Google Scholar] [CrossRef]
- Wang, B.; Xu, M.; Sun, G. Comparative Analysis of Membranous Proteomics of Shewanella Decolorationis S12 Grown with Azo Compound or Fe (III) Citrate as Sole Terminal Electron Acceptor. Appl. Microbiol. Biotechnol. 2010, 86, 1513–1523. [Google Scholar] [CrossRef]
- Zhang, S.; An, X.; Gong, J.; Xu, Z.; Wang, L.; Xia, X.; Zhang, Q. Molecular Response of Anoxybacillus Sp. PDR2 under Azo Dye Stress: An Integrated Analysis of Proteomics and Metabolomics. J. Hazard. Mater. 2022, 438, 129500. [Google Scholar] [CrossRef]
- Ng, I.-S.; Zheng, X.; Chen, B.-Y.; Chi, X.; Lu, Y.; Chang, C.-S. Proteomics Approach to Decipher Novel Genes and Enzymes Characterization of a Bioelectricity-Generating and Dye-Decolorizing Bacterium Proteus Hauseri ZMd44. Biotechnol. Bioprocess Eng. 2013, 18, 8–17. [Google Scholar] [CrossRef]
- An, X.; Chen, Y.; Chen, G.; Feng, L.; Zhang, Q. Integrated Metagenomic and Metaproteomic Analyses Reveal Potential Degradation Mechanism of Azo Dye-Direct Black G by Thermophilic Microflora. Ecotoxicol. Environ. Saf. 2020, 196, 110557. [Google Scholar] [CrossRef]
- Lin, L.; Wang, X.; Cao, L.; Xu, M. Lignin Catabolic Pathways Reveal Unique Characteristics of Dye-Decolorizing Peroxidases in Pseudomonas Putida. Environ. Microbiol. 2019, 21, 1847–1863. [Google Scholar] [CrossRef] [PubMed]
- Salvachúa, D.; Martínez, A.T.; Tien, M.; López-Lucendo, M.F.; García, F.; de los Ríos, V.; Martínez, M.J.; Prieto, A. Differential Proteomic Analysis of the Secretome of Irpex lacteus and Other White-Rot Fungi during Wheat Straw Pretreatment. Biotechnol. Biofuels 2013, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Ng, I.-S.; Xu, F.; Ye, C.; Chen, B.-Y.; Lu, Y. Exploring Metal Effects and Synergistic Interactions of Ferric Stimulation on Azo-Dye Decolorization by New Indigenous Acinetobacter guillouiae Ax-9 and Rahnella aquatilis DX2b. Bioprocess Biosyst. Eng. 2013, 37, 217–224. [Google Scholar] [CrossRef]
- Sharma, B.; Tiwari, S.; Kumar, R.; Kumar, M.; Tewari, L. Eco-Friendly Detoxification of Hazardous Congo Red Dye Using Novel Fungal Strain Trametes flavida WTFP2: Deduced Enzymatic Biomineralization Process through Combinatorial In-Silico and In-Vitro Studies. J. Hazard. Mater. 2023, 455, 131503. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Chinnathambi, V.; Arumugam, P.; Suresh, P.K. In silico and in vitro Physicochemical Screening of Rigidoporus Sp. Crude Laccase-Assisted Decolorization of Synthetic Dyes—Approaches for a Cost-Effective Enzyme-Based Remediation Methodology. Appl. Biochem. Biotechnol. 2013, 169, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Shanmugam, G.; Surwase, S.V.; Jadhav, J.P. Senthil Kumar Sadasivam in silico Analysis of Bacterial Systems for Textile Azo Dye Decolorization and Affirmation with Wetlab Studies. Clean Soil Air Water 2017, 45, 1600734. [Google Scholar] [CrossRef]
- Srinivasan, S.; Sadasivam, S.K.; Gunalan, S.; Shanmugam, G.; Kothandan, G. Application of Docking and Active Site Analysis for Enzyme Linked Biodegradation of Textile Dyes. Environ. Pollut. 2019, 248, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Haghshenas, H.; Kay, M.; Dehghanian, F.; Tavakol, H. Molecular Dynamics Study of Biodegradation of Azo Dyes via Their Interactions with AzrC Azoreductase. J. Biomol. Struct. Dyn. 2015, 34, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Thakuria, B.; Jungai, N.; Adhikari, S. Catalytic Site Prediction of Azoreductase Enzyme of E. coli with Potentially Important Industrial Dyes Using Molecular Docking Tools. Int. J. Biosci. Biochem. Bioinform. 2015, 5, 91–99. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Kong, L.-F.; Li, Y.-Y.; Xie, Z.-X. Environmental Microbial Community Proteomics: Status, Challenges and Perspectives. Int. J. Mol. Sci. 2016, 17, 1275. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Sharma, K. Microbial Biodiversity and Bioremediation Assessment through Omics Approaches. Front. Environ. Chem. 2020, 1, 570326. [Google Scholar] [CrossRef]
- Mishra, S.; Lin, Z.; Pang, S.; Zhang, W.; Bhatt, P.; Chen, S. Recent Advanced Technologies for the Characterization of Xenobiotic-Degrading Microorganisms and Microbial Communities. Front. Bioeng. Biotechnol. 2021, 9, 632059. [Google Scholar] [CrossRef]
- Villas-Bôas, S.G.; Bruheim, P. The potential of metabolomics tools in bioremediation studies. OMICS A J. Integr. Biol. 2007, 11, 305–313. [Google Scholar] [CrossRef]
- Rodríguez, A.; Castrejón-Godínez, M.L.; Salazar-Bustamante, E.; Gama-Martínez, Y.; Sánchez-Salinas, E.; Mussali-Galante, P.; Tovar-Sánchez, E.; Ortiz-Hernández, M.L. Omics Approaches to Pesticide Biodegradation. Curr. Microbiol. 2020, 77, 545–563. [Google Scholar] [CrossRef]
- Lin, C.Y.; Viant, M.R.; Tjeerdema, R.S. Metabolomics: Methodologies and applications in the environmental sciences. J. Pestic. Sci. 2006, 31, 245–251. [Google Scholar] [CrossRef]
- Callaghan, A.V. Metabolomic investigations of anaerobic hydrocarbon-impacted environments. Curr. Opin. Biotechnol. 2013, 24, 506–515. [Google Scholar] [CrossRef]
- Sun, J.; Jin, J.; Beger, R.D.; Cerniglia, C.E.; Chen, H. Evaluation of metabolism of azo dyes and their effects on Staphylococcus aureus metabolome. J. Ind. Microbiol. Biotechnol. 2017, 44, 1471–1481. [Google Scholar] [CrossRef]
- Zheng, X.; Xie, X.; Liu, Y.; Cong, J.; Fan, J.; Fang, Y.; Liu, N.; He, Z.; Liu, J. Deciphering the mechanism of carbon sources inhibiting recolorization in the removal of refractory dye: Based on an untargeted LC-MS metabolomics approach. Bioresour. Technol. 2020, 307, 123248. [Google Scholar] [CrossRef]
- Cao, X.; Qi, Y.; Xu, C.; Yang, Y.; Wang, J. Transcriptome and metabolome responses of Shewanella oneidensis MR-1 to methyl orange under microaerophilic and aerobic conditions. Appl. Microbiol. Biotechnol. 2017, 101, 3463–3472. [Google Scholar] [CrossRef]
- An, X.; Li, N.; Zhang, S.; Han, Y.; Zhang, Q. Integration of proteome and metabolome profiling to reveal heat stress response and tolerance mechanisms of Serratia sp. AXJ-M for the bioremediation of papermaking black liquor. J. Hazard. Mater. 2023, 450, 131092. [Google Scholar] [CrossRef]
- Li, R.; Li, T.; Wan, Y.; Zhang, X.; Liu, X.; Li, R.; Pu, H.; Gao, T.; Wang, X.; Zhou, Q. Efficient decolorization of azo dye wastewater with polyaniline/graphene modified anode in microbial electrochemical systems. J. Hazard. Mater. 2021, 421, 126740. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Database | Accession Number | The Place of Isolation | Decolorization Capability | References |
|---|---|---|---|---|---|
| Kocuria indica DP-K7 | DDBJ/ENA/GenBank | WMHZ00000000 | Sediment sample; Chorao Island, India | Decolorization of 68% Methyl Red after 160 h of incubation. | [81] |
| Shewanella sp. LC2 | GenBank | VFSJ00000000 | Textile industry wastewater effluent; Peru | Decolorization of 90% to 97% Direct Blue 71, Remazol Red, and Remazol Yellow Gold after 24 h of incubation | [82] |
| Shewanella sp. LC6 | GenBank | VFSK00000000 | |||
| Pseudomonas aeruginosa PFK10 | GenBank | AZBM00000000 | Effluent treatment plant; Gujarat, India | Capable of degrading various mono-azo, di-azo, tri-azo, and poly-azo textile dyes utilized in dyeing and printing industries | [83] |
| Shewanella xiamenensis BC01 | GenBank | JGVI00000000 | Seawater sample; Xiamen, China | Decolorization of textile azo dyes and production of nanowires in cells, responsive to electron transfer, thereby accelerating azo dye reduction | [84] |
| Proteus hauseri ZMd44 | GenBank | AWXP00000000 | Lanyang Plain, Taiwan | Utilized for applications in microbe fuel cells with wastewater using azo dye Reactive Blue 160 | [85] |
| Bacillus amyloliquefaciens AD20 | PATRICbrc | 1390.742 | Dye waste pond near a textile factory; Nigeria | Exhibited Azo dye decolorization ability | [86] |
| Bacillus subtilis C3 | DDBJ/EMBL/GenBank | JYOG00000000 | Effluent treatment plant; Gujarat, India | Displayed approximately 95% to 100% decolorization of various azo dyes during 24 to 48 h of incubation under static culture conditions | [87] |
| Priestia aryabhattai BD1 | DDBJ/ENA/GenBank | JANIPC000000000 | Dye waste sediment behind a textile company; Lagos, Nigeria, | Decolorization of Cibacron dyes—Reactive Blue 4% Red and Brilliant Yellow 6% Green | [88] |
| Novibacillus thermophiles SG-1 | GenBank | CP019699.1 | Saline soil sample; Guangdong Province, China | Orange I decolorization | [89] |
| Anoxybacillus sp. PDR2 | GenBank | CP047158 | Soil near a textile dye manufacturing factory; China | Decolorization of 82–98% of Direct Black G (100–600 mg/L) within 48 h | [90] |
| Shewanella algae 2NE11 | GenBank | CP055159 | Olive processing company effluent; Tacna, Peru | Decolorization of azo and anthraquinone dyes with a decolorization rate of 89–97% | [91] |
| Consortium of bacteria: Stenotrophomonas acidaminiphila APG1, Pseudomonas stutzeri APG2, and Cellulomonas sp. APG4 | GenBank | APG1: JAACYG000000000 APG2: JAACYH000000000 APG4: JAACYI000000000 | Sediment of the Alang ship recycling yard, Gujarat, India | Degradation of the mono-azo dye, Reactive Blue 28, and the aromatic amines released upon the cleavage of azo bond | [92] |
| Consortium of halo-thermophilic bacteria Alteribacillus as the dominant genus | nf. | nf. | nf. | Significant decolorization ability under a wide range of salinity (1–10%), pH (7–9), and temperature (45–60 °C) | [93] |
| Pantoea ananatis Sd-1 | GenBank | AZTE00000000 | Endophytic bacteria from rice seeds; China | Decolorization of various synthetic dyes | [94] |
| Tenacibaculum sp. HMG1 | GenBank | LDOD00000000 | Deep sea sediment; Pacific Ocean | Almost complete decolorization of Malachite Green within 12 h | [95] |
| Brevibacterium limosum sp. Nov., Brevibacterium pigmenatum sp. Nov., Brevibacterium atlanticum sp. Nov. | GenBank | B. limosum: KU560320 B. pigmenatum: KU560298 B. atlanticum: MN463008 | Sediment samples; South Atlantic and Western Pacific | Decolorization of Congo Red, Toluidine Blue, and Reactive Blue | [96] |
| Streptococcus thermophilus CGMCC 7.179 | GenBank | KR106994 | Fermented dairy products; Inner Mongolia | Production of peroxidase with the ability to decolorize Reactive Blue 5. | [97] |
| Halomonas pacifica M31 and Shewanella algae M69b | GenBank | M31: JAQQYI000000000 M69b: JAQQYH000000000 | Uninhabited coastal Red Sea region; Saudi Arabia | A 77% decolorization of dyes in artificial textile effluent derived from a mixture of Indigo carmine, Malachite Green, Cotton Blue, Bromocresol Green, and Reactive Red 66 | [98] |
| Pseudomonas stutzeri, AK6 | GenBank | PZYR00000000 | Textile effluent collected from the dye manufacturing industry; Gujarat, India. | Decolorization of 86% of Acid Blue 113 within 96 h. | [99] |
| Salinivibrio kushneri HTSP | GenBank | PXUD00000000 | Salt pan; Marakkanam, India | More than 80% decolorization within 48 h for Coomassie Brilliant Blue G-250 (500–3000 mg/L), Safranin, and Congo Red (50–800 mg/L) | [100] |
| Schizopora paradoxa KUC8140 | DDBJ/EMBL/GenBank | LBNM00000000 | Oakwood decay; Korea | Remazol Brilliant Blue R decolorization activity | [101] |
| Schizophyllum commune IUM1114-SS01 | GenBank | JAATOI000000000 | The Culture Collection of Mushrooms at Incheon National University | Dye decolorization ability from 50% up to 97% with the dyes: Crystal Violet, Amaranth Red, Brilliant Blue G, and Congo Red | [102] |
| Microorganism | Dye(s) | Technique Used | Identified Proteins Involved in Decolorization | References |
|---|---|---|---|---|
| Anoxybacillus sp. | Direct Black G | Tandem Mass Tag quantitative proteomics technology | Thioredoxin reductase | [115] |
| Proteus hauseri ZMd44 | Reactive Blue 160 | nano-LC-MS/MS | Laccase, dehydrogenase, and porin | [116] |
| Shewanella seohaensis NIODMS14 | Reactive Black 5, Reactive Green 19 Reactive Red 120 | LC-MS-QTOF | Oxidoreductases and an alkyl hydroperoxide reductase | [111] |
| Leptosphaerulina sp. CECT (20913) | Reactive Black 5 | Nano-LC-ion trap mass spectrometry | Oxidoreductases, peroxidases, flavo-oxidases, and laccases | [110] |
| Irpex lacteus CD2 | Direct Red 5 | Multidimensional Protein Identification Technology | Peroxidases | [105] |
| Lentinus squarrosulus AF5 | Dye mix consisting of: Acid Black 10B, Reactive Black 5, Reactive Blue 160 | LC-QTOF-MS | Peroxidase and oxidoreductase | [112] |
| Myrothecium roridum IM6482 | Acid Blue 113, Acid Red 27, Direct Blue 14, Acid Orange 7 | 2D-LC-MS/MS | Laccase | [113] |
| A thermophilic microflora | Direct Black G | 2D-LC-MS/MS | NADH-ubiquinone reductase and NADH-quinone oxidoreductase | [117] |
| Pseudomonas putida A514 | na. | LC-MS - | Peroxidases | [118] |
| I. lacteus SSF and I. lacteus SmF | Azo and anthraquinone dyes | MALDI-TOF-MS/ MS | Dye-decolorizing and manganese-oxidizing peroxidases | [119] |
| S. decolorationis S12 | Amaranth | 2-DE and MALDI-TOF/TOF/MS | nucleoside diphosphate kinase, ATP-dependent Clp protease proteolytic subunit, and N-acetyl-gamma-glutamylphosphate reductase | [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasińska, A.; Walaszczyk, A.; Paraszkiewicz, K. Omics-Based Approaches in Research on Textile Dye Microbial Decolorization. Molecules 2024, 29, 2771. https://doi.org/10.3390/molecules29122771
Jasińska A, Walaszczyk A, Paraszkiewicz K. Omics-Based Approaches in Research on Textile Dye Microbial Decolorization. Molecules. 2024; 29(12):2771. https://doi.org/10.3390/molecules29122771
Chicago/Turabian StyleJasińska, Anna, Aleksandra Walaszczyk, and Katarzyna Paraszkiewicz. 2024. "Omics-Based Approaches in Research on Textile Dye Microbial Decolorization" Molecules 29, no. 12: 2771. https://doi.org/10.3390/molecules29122771
APA StyleJasińska, A., Walaszczyk, A., & Paraszkiewicz, K. (2024). Omics-Based Approaches in Research on Textile Dye Microbial Decolorization. Molecules, 29(12), 2771. https://doi.org/10.3390/molecules29122771