
Antioxidant Properties of Platycladus orientalis Flavonoids for Treating UV-Induced Damage in Androgenetic Alopecia Hair


Abstract
:1. Introduction
2. Results
2.1. Analysis of Components of Extracts from Platycladus orientalis Leaves
2.2. The Impact of UV Radiation on the Hair of Individuals with AGA and Healthy Individuals
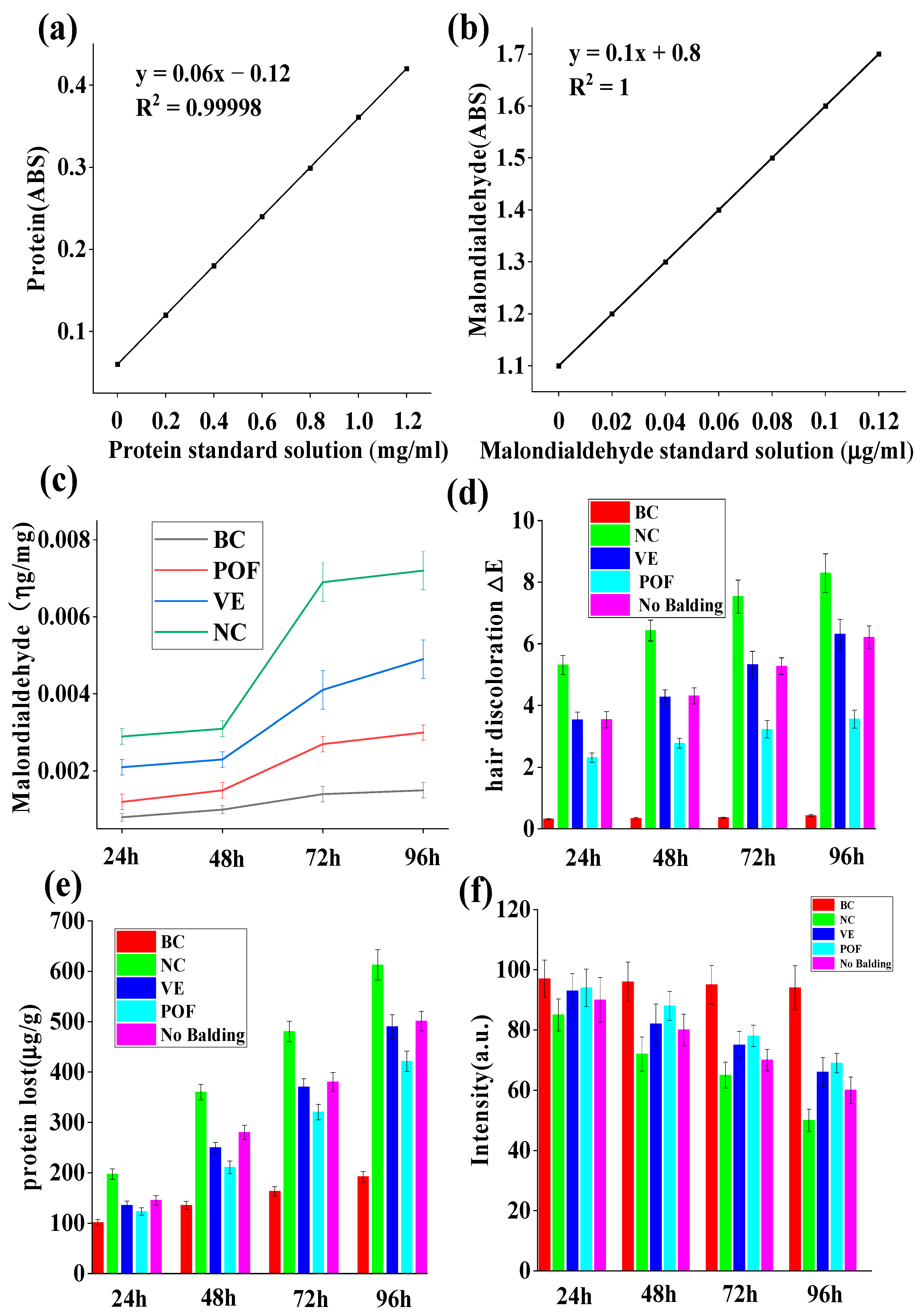
2.3. Hair’s Lipid Protection
2.4. Hair Discoloration
2.5. Protein Loss Protection
2.6. Tryptophan Protection
2.7. Tensile Properties
2.8. Preventing UV Damage of AGA Hair Using POFs
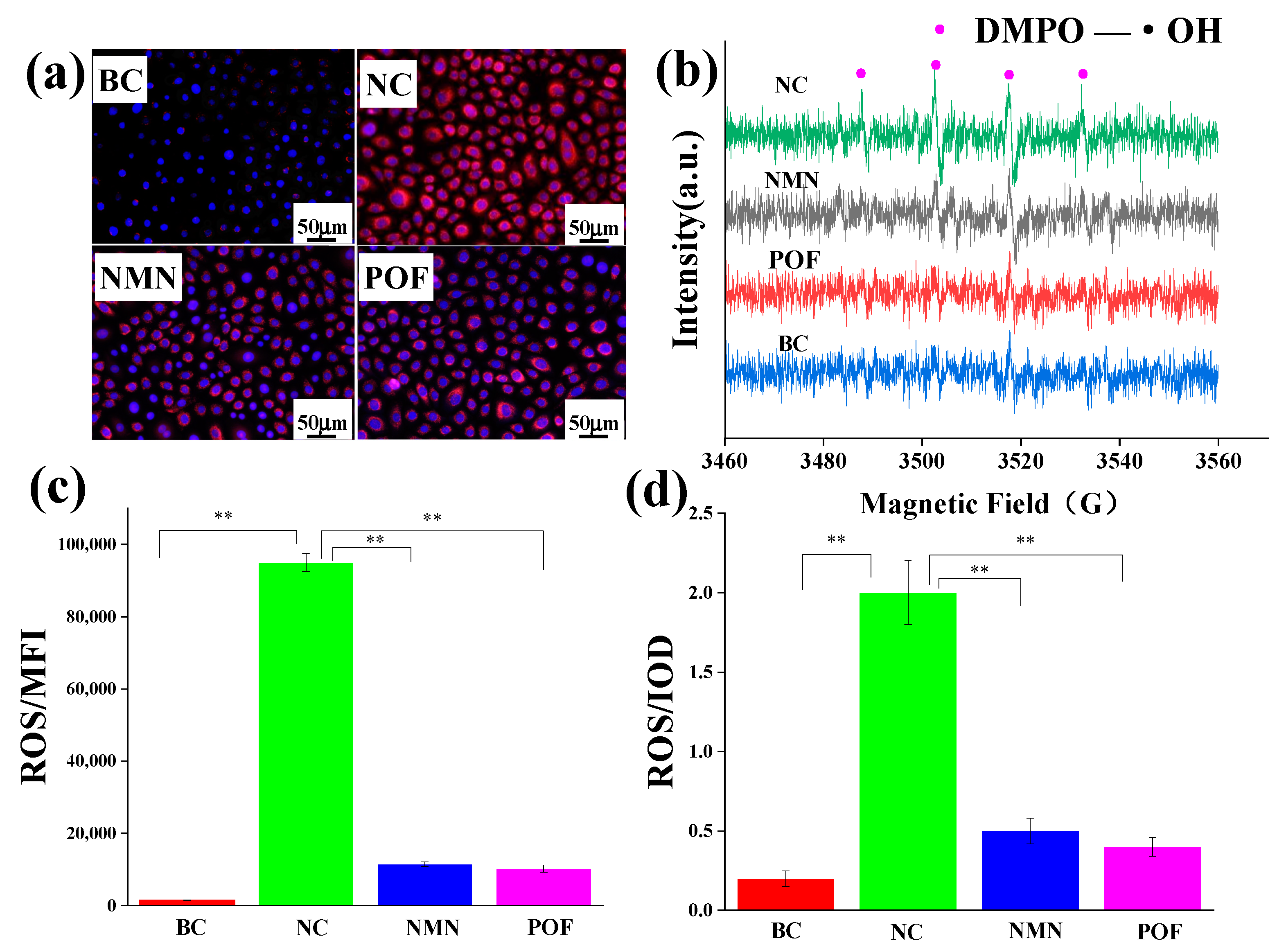
2.9. The Scavenging Ability of Free Radicals (ROS)
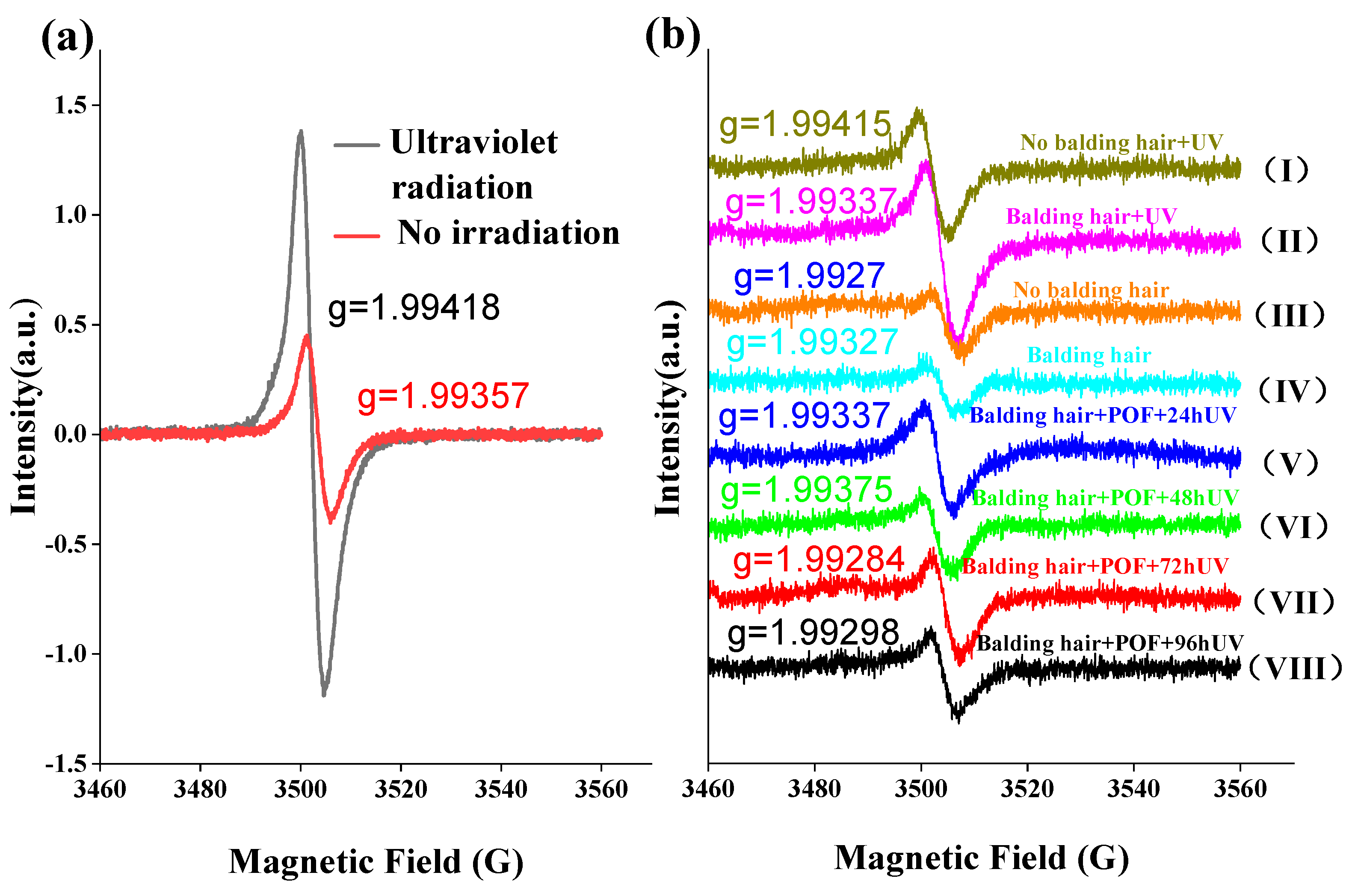
2.10. The Effect of UV Irradiation on Free Radicals in AGA Hair
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Extraction and Testing of POF Components
4.3. Preparation of Hair Samples and SEM Scanning
4.4. UV Irradiation of Hair
4.5. Lipid Peroxide Test
4.6. Chromatism
4.7. Protein Loss
4.8. Tryptophan Content Test
4.9. Mechanical Properties of Hair
4.10. Antioxidant Test
4.11. EPR
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chang, E.; Guo, W.; Xie, Y.; Jiang, Z.; Dong, Y.; Jia, Z.; Zhao, X.; Liu, J.; Zhang, J. Changes of lignified-callus and wound-induced adventitious rooting in ancient Platycladus orientalis cuttings as affected by tree age. Ind. Crop. Prod. 2023, 203, 117183. [Google Scholar] [CrossRef]
- Anshiba, J.; Poonkothai, M.; Karthika, P.; Mythili, R. Green route to a novel and ecofriendly phytosynthesis of silver nanoparticles using Platycladus orientalis L. leaf extract. Mater. Lett. 2022, 309, 131347. [Google Scholar] [CrossRef]
- Chen, L.; Ding, L.; Yu, A.; Yang, R.; Wang, X.; Li, J.; Jin, H.; Zhang, H. Continuous determination of total flavonoids in Platycladus orientalis (L.) Franco by dynamic microwave-assisted extraction coupled with on-line derivatization and ultraviolet–visible detection. Anal. Chim. Acta 2007, 596, 164–170. [Google Scholar] [CrossRef]
- Fan, S.-Y.; Zeng, H.-W.; Pei, Y.-H.; Li, L.; Ye, J.; Pan, Y.-X.; Zhang, J.-G.; Yuan, X.; Zhang, W.-D. The anti-inflammatory activities of an extract and compounds isolated from Platycladus orientalis (Linnaeus) Franco in vitro and ex vivo. J. Ethnopharmacol. 2012, 141, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Mu, K.; Liu, Y.; Liu, G.; Ran, F.; Zhou, L.; Wu, Y.; Peng, L.; Shao, M.; Li, C.; Zhang, Y. A review of hemostatic chemical components and their mechanisms in traditional Chinese medicine and ethnic medicine. J. Ethnopharmacol. 2023, 307, 116200. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Qiang, W.; Jiang, J.; Hu, X.; Chen, Y.; Guo, Y.; Liu, H.; Sun, S.; Gao, H.; Zhang, Y.; et al. Safflower oil body nanoparticles deliver hFGF10 to hair follicles and reduce microinflammation to accelerate hair regeneration in androgenetic alopecia. Int. J. Pharmaceut. 2022, 616, 121537. [Google Scholar]
- Wei, J.; Zhao, G.; Wu, G.; Bo, Y.; Yang, D.; Guo, J.; Ma, Y.; An, M. An ultrasound-assisted extraction using an alcohol-based hydrophilic natural deep eutectic solvent for the determination of five flavonoids from Platycladi Cacumen. Microchem. J. 2024, 198, 110076. [Google Scholar] [CrossRef]
- Hobo, Y.; Nishikawa, J.; Miyashiro, Y.; Fujikata, A. Analysis of hair steroid hormone concentrations at different parts of the head by liquid chromatography-tandem mass spectrometry. Clin. Chim. Acta 2021, 523, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Ntshingila, S.; Oputu, O.; Arowolo, A.T.; Khumalo, N.P. Androgenetic alopecia: An update. JAAD Int. 2023, 13, 150–158. [Google Scholar] [CrossRef]
- Adav, S.S.; Ng, K.W. Recent omics advances in hair aging biology and hair biomarkers analysis. Ageing Res. Rev. 2023, 91, 102041. [Google Scholar] [CrossRef]
- Geisler, A.N.; Austin, E.; Nguyen, J.; Hamzavi, I.; Jagdeo, J.; Lim, H.W. Visible light. Part II: Photoprotection against visible and ultraviolet light. J. Am. Acad. Dermatol. 2021, 84, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Richena, M.; Rezende, C.A. Effect of photodamage on the outermost cuticle layer of human hair. J. Photochem. Photobiol. B Biol. 2015, 153, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Richena, M.; Rezende, C.A. Morphological degradation of human hair cuticle due to simulated sunlight irradiation and washing. J. Photochem. Photobiol. B Biol. 2016, 161, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, T.B.; Hudari, F.; Munnia, A.; Peluso, M.; Godschalk, R.W.; Zanoni, M.V.B.; den Hartog, G.J.M.; Bast, A.; Barros, S.B.M.; Maria-Engler, S.S.; et al. The oxidation of p-phenylenediamine, an ingredient used for permanent hair dyeing purposes, leads to the formation of hydroxyl radicals: Oxidative stress and DNA damage in human immortalized keratinocytes. Toxicol. Lett. 2015, 239, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Mokrzyński, K.; Sarna, M.; Sarna, T. Photoreactivity and phototoxicity of experimentally photodegraded hair melanosomes from individuals of different skin phototypes. J. Photochem. Photobiol. B Biol. 2023, 243, 112704. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Zhou, X.; McCallum, N.C.; Hu, Z.; Ni, Q.Z.; Kapoor, U.; Heil, C.M.; Cay, K.S.; Zand, T.; Mantanona, A.J.; et al. Unraveling the Structure and Function of Melanin through Synthesis. J. Am. Chem. Soc. 2021, 143, 2622–2637. [Google Scholar] [CrossRef] [PubMed]
- Chiarelli-Neto, O.; Pavani, C.; Ferreira, A.S.; Uchoa, A.F.; Severino, D.; Baptista, M.S. Generation and suppression of singlet oxygen in hair by photosensitization of melanin. Free Radic. Biol. Med. 2011, 51, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ji, X.; Mukherjee, S.; Yang, B.; Ren, Y.; Wang, C.; Chen, Y. A Bioinspired Skin UV Filter with Broadband UV Protection, Photostability, and Resistance to Oxidative Damage. ACS Appl. Mater. Interfaces 2023, 15, 10383–10397. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, R.; Kolling, D.R.J.; Shoup, D.; Smythers, A.L.; Nickel, S.; Westfall, T.D.; Epperly, C.; Coplin, M. Comparing Free Radicals in Sunscreen-Treated Pig Skin by Using Electron Paramagnetic Resonance Spectroscopy. J. Chem. Educ. 2019, 96, 2021–2028. [Google Scholar] [CrossRef]
- Prasertpol, T.; Tiyaboonchai, W. Nanostructured lipid carriers: A novel hair protective product preventing hair damage and discoloration from UV radiation and thermal treatment. J. Photochem. Photobiol. B Biol. 2020, 204, 111769. [Google Scholar] [CrossRef]
- Eggen, M.D.; Merboth, P.; Neukirchner, H.; Glomb, M.A. Lipid Peroxidation Has Major Impact on Malondialdehyde-Derived but Only Minor Influence on Glyoxal and Methylglyoxal-Derived Protein Modifications in Carbohydrate-Rich Foods. J. Agric. Food Chem. 2022, 70, 10271–10283. [Google Scholar] [CrossRef]
- Adams, A.; De Kimpe, N.; van Boekel, M.A.J.S. Modification of Casein by the Lipid Oxidation Product Malondialdehyde. J. Agric. Food Chem. 2008, 56, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Matamá, T.; Costa, C.; Fernandes, B.; Araújo, R.; Cruz, C.F.; Tortosa, F.; Sheeba, C.J.; Becker, J.D.; Gomes, A.; Cavaco-Paulo, A. Changing human hair fibre colour and shape from the follicle. J. Adv. Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Fernández, E.; Barba, C.; Alonso, C.; Martí, M.; Parra, J.L.; Coderch, L. Photodamage determination of human hair. J. Photochem. Photobiol. B Biol. 2012, 106, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Wang, Y.; Cui, W. Advanced electrospun hydrogel fibers for wound healing. Compos. Part B Eng. 2021, 223, 109101. [Google Scholar] [CrossRef]
- Hirai, T.; Ikeda-Imafuku, M.; Tasaka, N.; Chuang, V.T.G.; Xian, M.; Ishida, T.; Akaike, T.; Ishima, Y. Human hair keratin responds to oxidative stress via reactive sulfur and supersulfides. Adv. Redox Res. 2024, 10, 100091. [Google Scholar] [CrossRef]
- Huang, Y.; Song, B.; Chen, K.; Kong, D.; Yuan, J. Time-gated luminescent probes for lysosomal singlet oxygen: Synthesis, characterizations and bioimaging applications. Anal. Chim. Acta 2024, 1287, 342063. [Google Scholar] [CrossRef] [PubMed]
- Boulebd, H.; Pereira, D.M. Examination of the Antioxidant Activity of Psoralidin: Computational Mechanistic Study and Impact on the ROS Level in Human Keratinocytes. J. Org. Chem. 2023, 88, 5745–5751. [Google Scholar] [CrossRef] [PubMed]
- Beberok, A.; Zdybel, M.; Pilawa, B.; Buszman, E.; Wrześniok, D. EPR characteristics of free radicals in DOPA–melanin–moxifloxacin complexes at ambient level of UVA radiation. Chem. Phys. Lett. 2014, 592, 41–46. [Google Scholar] [CrossRef]
- Demir, E.; Acaralı, N. Comparison on Quality Performance of Human Hair Types with Herbal Oils (Grape Seed/Safflower Seed/Rosehip) by Analysis Techniques. ACS Omega 2023, 8, 8293–8302. [Google Scholar] [CrossRef]
- Fernández, E.; Martínez-Teipel, B.; Armengol, R.; Barba, C.; Coderch, L. Efficacy of antioxidants in human hair. J. Photochem. Photobiol. B Biol. 2012, 117, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Truong, V.-L.; Jeong, W.-S. Red ginseng (Panax ginseng Meyer) oil: A comprehensive review of extraction technologies, chemical composition, health benefits, molecular mechanisms, and safety. J. Ginseng Res. 2022, 46, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Dario, M.F.; Viana, A.S.; Augusto, O.; Linares, E.; Gama, R.M.; Baby, A.R.; Velasco, M.V.R. Dyed hair photoprotection efficacy of a quercetin-loaded cationic nanoemulsion. J. Photochem. Photobiol. B Biol. 2020, 204, 111788. [Google Scholar] [CrossRef] [PubMed]
- Roets, B. Potential application of PBM use in hair follicle organoid culture for the treatment of androgenic alopecia. Mater. Today Bio 2023, 23, 100851. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.-D.; Fan, H.-L.; Jiao, D.-Q.; Li, F.; He, Y.-N.; Wu, Q.-L.; Liu, X.-L.; Wang, C.-H. Rational engineering S1’ substrate binding pocket to enhance substrate specificity and catalytic activity of thermal-stable keratinase for efficient keratin degradation. Int. J. Biol. Macromol. 2024, 263, 130688. [Google Scholar] [CrossRef] [PubMed]
- Harland, D.P.; Popescu, C.; Richena, M.; Deb-Choudhury, S.; Wichlatz, C.; Lee, E.; Plowman, J.E. The susceptibility of disulfide bonds to modification in keratin fibers undergoing tensile stress. Biophys. J. 2022, 121, 2168–2179. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.; Martins, M.; Tinoco, A.; Cavaco-Paulo, A. Hair resistance to mechanical wear. Wear 2021, 470–471, 203612. [Google Scholar] [CrossRef]
- Fedorkova, M.V.; Smolina, N.V.; Mikhalchik, E.V.; Balabushevich, N.G.; Ibragimova, G.A.; Gadzhigoroeva, A.G.; Dmitrieva, E.I.; Dobretsov, G.E. Effects of ultra violet radiation on the soluble proteins of human hair. J. Photochem. Photobiol. B Biol. 2014, 140, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Dario, M.F.; Baby, A.R.; Velasco, M.V.R. Effects of solar radiation on hair and photoprotection. J. Photochem. Photobiol. B Biol. 2015, 153, 240–246. [Google Scholar] [CrossRef]
- Breakspear, S.; Smith, J.R.; Luengo, G. Effect of the covalently linked fatty acid 18-MEA on the nanotribology of hair’s outermost surface. J. Struct. Biol. 2005, 149, 235–242. [Google Scholar] [CrossRef]
- Di Mascio, P.; Martinez, G.R.; Miyamoto, S.; Ronsein, G.E.; Medeiros, M.H.G.; Cadet, J. Singlet Molecular Oxygen Reactions with Nucleic Acids, Lipids, and Proteins. Chem. Rev. 2019, 119, 2043–2086. [Google Scholar] [CrossRef] [PubMed]
- Bando, N.; Hayashi, H.; Wakamatsu, S.; Inakuma, T.; Miyoshi, M.; Nagao, A.; Yamauchi, R.; Terao, J. Participation of singlet oxygen in ultraviolet-a-induced lipid peroxidation in mouse skin and its inhibition by dietary β-carotene: An ex vivo study. Free Radic. Biol. Med. 2004, 37, 1854–1863. [Google Scholar] [CrossRef] [PubMed]
- Bacellar, I.O.L.; Baptista, M.S. Mechanisms of Photosensitized Lipid Oxidation and Membrane Permeabilization. ACS Omega 2019, 4, 21636–21646. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Carroll, L.; Rasmussen, L.M.; Davies, M.J. Oxidation of protein disulfide bonds by singlet oxygen gives rise to glutathionylated proteins. Redox Biol. 2021, 38, 101822. [Google Scholar] [CrossRef] [PubMed]
- Buchner, O.; Roach, T.; Gertzen, J.; Schenk, S.; Karadar, M.; Stöggl, W.; Miller, R.; Bertel, C.; Neuner, G.; Kranner, I. Drought affects the heat-hardening capacity of alpine plants as indicated by changes in xanthophyll cycle pigments, singlet oxygen scavenging, α-tocopherol and plant hormones. Environ. Exp. Bot. 2017, 133, 159–175. [Google Scholar] [CrossRef]
- Kim, E.; Panzella, L.; Napolitano, A.; Payne, G.F. Redox Activities of Melanins Investigated by Electrochemical Reverse Engineering: Implications for their Roles in Oxidative Stress. J. Investig. Dermatol. 2020, 140, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Qu, H.; Xiong, C.; Liu, C.; Zheng, L. A novel method for non-destructive determination of hair photo-induced damage based on multispectral imaging technology. Sci. Rep. (Nat. Publ. Group) 2017, 7, 45544. [Google Scholar] [CrossRef] [PubMed]
- Carolina Botelho, L.; Gasparin, R.M.; Thomaz, F.M.; da Silva, G.C.; Airton Abrahao, M.; Paiva-Santos, A.C.; Priscila Gava, M. Impact of Hair Damage on the Penetration Profile of Coconut, Avocado, and Argan Oils into Caucasian Hair Fibers. Cosmetics 2024, 11, 64. [Google Scholar] [CrossRef]
- Busch-Kschiewan, K.; Zentek, J.; Wortmann, F.J.; Biourge, V. UV Light, Temperature, and Humidity Effects on White Hair Color in Dogs. J. Nutr. 2004, 134, 2053S–2055S. [Google Scholar] [CrossRef]
- Draelos, Z.D. Commentary: Healthy hair and protein loss. J. Am. Acad. Dermatol. 2010, 62, 409–410. [Google Scholar] [CrossRef]
- Bertazzo, A.; Biasiolo, M.; Costa, C.V.L.; Cardin de Stefani, E.; Allegri, G. Tryptophan in human hair: Correlation with pigmentation. Ill. Farm 2000, 55, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.H.; Debnath, S.; Mahapatra, R.K.; Nayak, L.K.; Baruah, S.; Das, A.; Banik, S.; Tamuli, M.K. Tensile properties of hair fibres obtained from different breeds of pigs. Biosyst. Eng. 2014, 119, 35–43. [Google Scholar] [CrossRef]
- Xu, C.; Dai, J.; Ai, H.; Du, W.; Ji, H. β-Nicotinamide Mononucleotide Promotes Cell Proliferation and Hair Growth by Reducing Oxidative Stress. Molecules 2024, 29, 798. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, S.W.; Moseley, P.L.; Buettner, G.R. Increased flux of free radicals in cells subjected to hyperthermia: Detection by electron paramagnetic resonance spin trapping. FEBS Lett. 1998, 431, 285–286. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Time (min) | Component | Content (%) | CAS |
|---|---|---|---|
| 3.824 | Myricetin | 5.15 | 529-44-2 |
| 4.490 | Quercetin | 1.34 | 522-12-3 |
| 5.113 | Afzelin | 1.54 | 482-39-3 |
| 5.770 | Aromadendrin | 2.28 | 480-20-6 |
| 6.277 | Hinokiflavone | 4.83 | 19202-36-9 |
| 7.061 | Amentoflavone | 4.37 | 1617-53-4 |
| 10.226 | Kaempferol | 2.41 | 520-18-3 |
| 13.342 | Rutin | 2.35 | 153-18-4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Dai, J.; Du, W.; Ji, H. Antioxidant Properties of Platycladus orientalis Flavonoids for Treating UV-Induced Damage in Androgenetic Alopecia Hair. Molecules 2024, 29, 2876. https://doi.org/10.3390/molecules29122876
Xu C, Dai J, Du W, Ji H. Antioxidant Properties of Platycladus orientalis Flavonoids for Treating UV-Induced Damage in Androgenetic Alopecia Hair. Molecules. 2024; 29(12):2876. https://doi.org/10.3390/molecules29122876
Chicago/Turabian StyleXu, Chuntao, Jiawei Dai, Weian Du, and Hongbing Ji. 2024. "Antioxidant Properties of Platycladus orientalis Flavonoids for Treating UV-Induced Damage in Androgenetic Alopecia Hair" Molecules 29, no. 12: 2876. https://doi.org/10.3390/molecules29122876
APA StyleXu, C., Dai, J., Du, W., & Ji, H. (2024). Antioxidant Properties of Platycladus orientalis Flavonoids for Treating UV-Induced Damage in Androgenetic Alopecia Hair. Molecules, 29(12), 2876. https://doi.org/10.3390/molecules29122876