Abstract
Angomonas deanei belongs to Trypanosomatidae family, a family of parasites that only infect insects. It hosts a bacterial endosymbiont in a mutualistic relationship, constituting an excellent model for studying organelle origin and cellular evolution. A lipidomic approach, which allows for a comprehensive analysis of all lipids in a biological system (lipidome), is a useful tool for identifying and measuring different expression patterns of lipid classes. The present study applied GC-MS and NMR techniques, coupled with principal component analysis (PCA), in order to perform a comparative lipidomic study of wild and aposymbiotic A. deanei grown in the presence or absence of FBS. Unusual contents of branched-chain iso C17:0 and C19:0-cis-9,10 and-11,12 fatty acids were identified in A. deanei cultures, and it was interesting to note that their content slightly decreased at the log phase culture, indicating that in the latter growth stages the cell must promote the remodeling of lipid synthesis in order to maintain the fluidity of the membrane. The combination of analytical techniques used in this work allowed for the detection and characterization of lipids and relevant contributors in a variety of A. deanei growth conditions.
1. Introduction
When it comes to protozoan–host interactions, the cell membrane, which constitutes the main interface between cells, provides not only a site for contact and invasion but also a mechanism for evading host defense mechanisms. Particularly in the case of symbiotic associations, the membrane environment plays a specific role in the exchange of metabolites and nutrients. Angomonas deanei (previously known as Crithidia deanei) [1,2] is a trypanosomatid that infects only insects. It may be grown in vitro with low nutritional requirements and hosts a β-proteobacterial endosymbiont [3]. The endosymbiont maintains a mutualistic relationship with the host protozoan [4], thus constituting an excellent model for studying organelle origin and cellular evolution [2,5,6,7]. Recently, Motta et al. [8] published the A. deanei genome and analyzed the predicted protein sequences of A. deanei and its symbiont, clarifying evolutionary aspects and providing information that led to a better understanding of this symbiotic relationship. In this symbiotic relationship, intense metabolic exchanges occur between both partners, the bacterium providing essential enzymes that complete important biosynthetic pathways of the protozoa (such as heme and ornithine synthesis) and in exchange receiving from the host suitable physical conditions and energy supply [1,9,10,11,12]. The symbiont is enclosed by two membranes and exhibits a reduced cell wall, essential for bacterial division and morphological maintenance [13]. Sterols are absent in symbiont membranes whose major phospholipid is cardiolipin, followed by similar amounts of phosphatidylcholine, phosphatidylethanolamine and a lesser proportion of phosphatidylinositol [14]. The endosymbiont enhances protozoan phospholipid production and depends in part on its host cell to obtain phosphatidylcholine [15]. A study conducted by Freitas-Junior et al. [16] showed the effects of a phosphatidylcholine biosynthesis inhibitor on mitochondrion and symbiotic fractions of A. deanei, reinforcing the idea that an intense metabolic exchange occurs between the host trypanosomatid and structures of symbiotic origin.
To date, few studies have focused on evaluating the lipid composition of A. deanei under various nutritional conditions; moreover, little information is available regarding how the lipid composition of this trypanosomatid changes when it is grown in the absence of this endosymbiont. Since A. deanei is not able to synthesize cholesterol, analyzing the lipid changes when grown in the absence of FBS would also provide significant insights regarding lipid metabolism. As a non-destructive technique, Nuclear Magnetic Resonance (NMR) can be used in the preliminary characterization of lipid metabolites, while multivariate analysis methodologies such as principal component analysis (PCA) can be used subsequently to point out the main contributors that distinguish cell lines or experimental data sets [17]. Furthermore, the current available data applying modern analytical techniques to lipid composition of Trypanosomatidae representants are insufficient, resulting in a need for researchers in this area to provide up-to-date data that can be used as a reference for comparative studies. The growing field encompassing neural networks and machine learning could greatly benefit from these analytical data to perform automated quantitative and qualitative lipidomic analysis with NMR raw data from different matrices. [18,19]. This study applied GC-MS and NMR techniques, coupled with principal component analysis (PCA), in order to characterize the lipidomic profile of A. deanei grown in the presence and absence of fetal bovine serum, as well as in aposymbiotic A. deanei, stressing the usability of multivariate analysis in discriminating biomarkers among the lipid classes.
2. Results
2.1. GC-MS of Derivatizated Fatty Acids
Total fatty acid composition of the A. deanei was determined after growth in a variety of conditions (Table S1). Choanomastigotes containing the symbiotic organism were harvested at the exponential growth phase, both without FBS (0% Log) and with 10% FBS (10% Log), as well as at the stationary growth phase without FBS (0% Sta). Aposymbiotic choanomastigotes were also harvested at log phase without FBS (0% Apo) and subjected to lipid extraction and derivatization, as described in the Materials and Methods section. Table 1 shows a predominance of unsaturated fatty acids, namely, C18:3, C18:1, C18:2, C19:1 and C20:3. As shown in Table S1, between the saturated fatty acids, the main component was the unusual branched fatty acid C17:0 (iso-heptadecanoic acid, as confirmed by mass spectrometry and chromatographic analysis) [20,21,22,23], along with C18:0 and another unusual fatty acid, C19 Cyclic (cis-9,10- methylene-octadecanoic acid).

Table 1.
The relative proportion of the fatty acid class of total lipid extracts from A. deanei under the four growth conditions, namely, 0% FBS collected at the exponential growth phase (0% Log); 10% FBS collected at the exponential growth phase (10% Log); 0% FBS aposymbiotic (0% Apo) and 0% FBS collected at the stationary growth phase (0% Sta).
The main variations found among the four conditions analyzed were related to MUFA and PUFA levels. As shown in Table 1, the MUFA levels increased for the 10% FBS (10% Log) and aposymbiotic (0% Apo) conditions, whereas the PUFA levels were greater for the 0% FBS Log (0% Log) and 0% FBS collected at the stationary phase (0% Sta). No variations were observed among the four conditions for overall levels of branched-chain fatty acids. This stands in contrast to cyclic fatty acid levels, which increased continuously among the 0% Log, 10% Log, 0% Apo and 0% Sta growth conditions, an increase that occurred with both isomers (cis-9,10-methylene-octadecanoic acid and cis-11,12-methylene-octadecanoic acid). The condition containing 10% FBS in the growth medium exhibited very similar behavior to that of the 0% Apo.
A comparison of the results of fatty acid analyses for 0% Sta and 0% Log growth conditions reveals changes in the proportion of several individual fatty acids, taking into account that both were grown in the same nutritional conditions. The stationary A. deanei culture showed lower proportions of C14, C15Bra, C16, C16:1, C16Bra, C17:1, C18, C20:3, C22, C22:1 and C24 fatty acids, despite the fact that the overall proportion of unsaturated fatty acids remained similar. On the other hand, C17Bra, C18:3, C19Bra and C19Cyc (both isomers) increased in proportion. These results indicate that in the latter stages of growth, the cell must promote the remodeling of lipid synthesis in order to maintain the fluidity of the membrane. Therefore, the fatty acid profile obtained at this phase of growth characterizes the lipidome of this microorganism when the cells are not undergoing division.
2.2. NMR Analysis of Lipid Extracts
One-dimensional and two-dimensional NMR analysis of the lipid extracts allowed for the identification of the main lipid assignments (Figure 1 and Table S2). As shown in Table 1, the conditions 0% Log and 0% Sta exhibited higher amounts of PUFA, an observation that is confirmed by examining the 1H NMR spectra of the four samples (Figure 1). NMR analysis also detected the cyclopropane fatty acid (CFA) at δ −0.32 ppm (Figure 2), which is characteristic of a cis CFA ring [24,25,26]. These data were backed by 1D selective TOCSY (Figure 2C), which showed the assignments of CFA ring characteristics of a CFA 19:0 cis-9,10-methylene-octadecanoic acid and the cis-11,12 isomer [25,26]. Two-dimensional 1H-13C multiplicity-edited HSQC NMR analyses allowed for the assignment of both 1H and 13C chemical shift of main lipid classes on the lipid extracts, as shown in Figure 3.
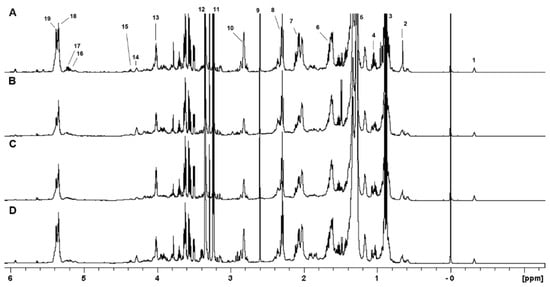
Figure 1.
The 1H NMR spectra of lipid extracts obtained from A. deanei grown in the absence of FBS (A), 10% FBS (B), aposymbiotic (C) and 0% FBS collected at the stationary growth phase (D). The chemical shifts are relative to the internal standard TMS (δ = 0.00 ppm). 1−CFA; 2−Erg−C18; 3−R−CH3; 4−Erg−C19; 5−(CH2)n; 6−Fβ:R−CH2−CH2−CO; 7−CH=CH−CH2−CH−CH (18:1); 8−Fα:R−CH2−CH2−CO; 9−Suc; 10−CH=CH−CH2−CH=CH (18:2); 11−N+(Me3)3; 12−CHD2OD; 13−Gly−C3 (PC/PE); 14−PC−1′; 15−Gly−C1 (DG/TG/PC/PE/PI); 16−Gly−C2 (DG); 17−Gly−C2 (PC/PE/PI); 18−Mono−UFA; 19−Poly−UFA.
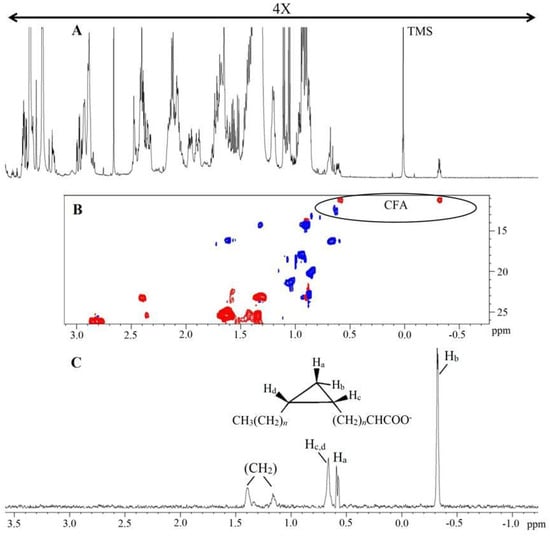
Figure 2.
1H NMR spectrum of lipid extract obtained from A. deanei (A), HSQC−ed slice of fingerprint region of CFA (B) and 1D selective TOCSY (60 ms) irradiated at Hb of CFA (C). Chemical shifts are relative to internal standard TMS (δ = 0.00 ppm). The positive phase (blue) corresponds to CH and CH3 first-order correlations and the negative phase (red) corresponds to CH2 first-order correlations.
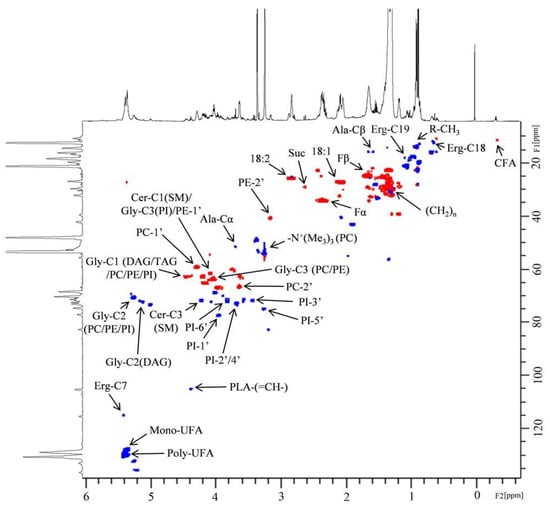
Figure 3.
Two-dimensional 1H-13C multiplicity-edited HSQC NMR correlation map from the lipid extract of A. deanei grown in the absence of FBS and the principal lipid assignments. The positive phase (blue) corresponds to CH and CH3 first-order correlations and the negative phase (red) corresponds to CH2 first-order correlations.
2.3. PCA
In order to further explore the 1H NMR data obtained from the lipid extracts, the authors applied multivariate analysis with the aim of identifying less intense 1H NMR assignments that could indicate lipid classes contributing to differentiation. Principal component analysis is a multivariate analysis technique which allows for the correlation between analytical data and experimental conditions [27,28]. PCA was carried out with 1H NMR spectra from lipid extracts of A. deanei (Figure 4). The four experimental conditions were separated into three distinct groups on the Scores plot (Figure 4A), and both the first (PC1, horizontal axis) and second (PC2, vertical axis) principal components contributed to the differentiation, resulting in a cumulative explained variance of 28.52% and 48.67%, respectively. The 0%Sta experimental condition showed a great PC2 discrimination, and the 1D loadings plot (Figure 4C) indicated the main 1H chemical shifts contributing to this separation. The 0%Log condition was separated from the other three conditions mainly by PC1, with chemical shifts assigned to PUFA and MUFA contributing to separation.
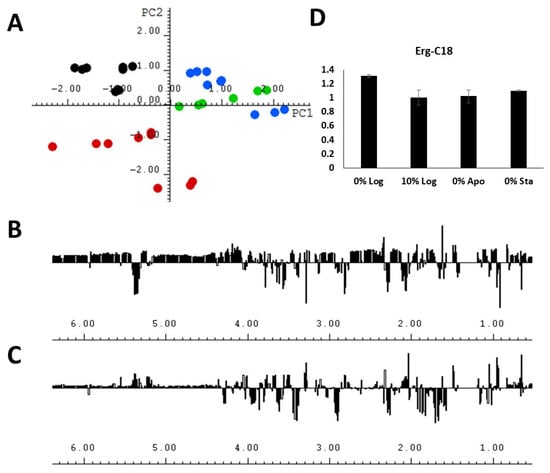
Figure 4.
(A) The score plots (PC1 vs. PC2) of the PCA analysis performed with the 1H NMR spectra of lipid extracts obtained from A. deanei grown in the absence of FBS (black dots), 10% FBS (blue dots), aposymbiotic condition (green dots) and 0% FBS collected at the stationary growth phase (red dots); (B) 1D loadings plot of PC1; (C) 1D loadings plot of PC2; (D) relative integral area of the chemical shift of Erg−C18 obtained from the lipid extracts under 4 experimental conditions.
An interesting observation was that it was not possible to fully discriminate the conditions 10% Log and 0% Apo by means of PC analysis, whether by PC1 or PC2, or even when a third principal component was plotted (Figure S1). As seen on the Score plot (Figure 4A) for both PC1 and PC2, one of the main chemical shifts that contributed to the differentiation was the signal near 0.65 ppm, which was assigned to Erg−C18, and the integration of this signal (Figure 4D and Table S3) showed no statistically significant difference in the area of this compound for the 10% Log and 0% Apo experimental conditions (Table S4), thus highlighting the usability of multivariate analysis in discriminating biomarkers among the lipid classes.
2.4. Sterol Analysis
The relative sterol composition showed a predominance of ergosterol (Supplementary Table S5). A. deanei grown in the absence of FBS showed a value of 93.9% for this sterol, whereas A. deanei grown in the presence of FBS and aposymbiotic A. deanei showed values of 87.0% and 67.5%, respectively (Supplementary Table S5). Low levels of cholesterol were detected—7.8% for A. deanei cultivated in the presence of FBS and 11.4% in the aposymbiotic condition. Fecosterol was found only in aposymbiotic A. deanei (11.2%).
3. Discussion
The current study used NMR and GC-MS techniques to evaluate lipidomic profiles of A. deanei with the aim of identifying lipid biochemical changes under different growth conditions. The NMR technique has been used widely for metabolite characterization in the Trypanosomatidae family in studies relating to the following: analysis of cell death due to visible mobile lipids in T. cruzi epimastigote [29]; metabolic studies of T. cruzi [30] and Crithidia luciliae [31]; 13C NMR analysis of glucose metabolism in Crithidia fasciculata [32] and T. cruzi [33]; analysis of metabolic end products in T. cruzi [34]; the structural characterization of a flagellar calcium-binding protein in T. brucei [35]; excreted product analysis using 1H NMR of trypanosome isolates [36]; high-resolution 31P NMR of T. brucei, T. cruzi and Leishmania major [37] and the characterization of the metabolic responses of mice to T. brucei brucei infection using 1H NMR [38].
The great similarity between the lipid profiles of A. deanei grown with 10% Log and 0% Apo, an observation that was verified by 1H NMR and GC-MS analysis, may be explained by the low nutritional requirements of the parasite, which does not even need FBS for growth. Unlike most trypanosomatids, A. deanei is not a nutrition-demanding protozoan, requiring only a few amino acids and vitamins supplemented in LIT medium for growth in vitro. Hemin or purine, indispensable for the growth of other trypanosomatids, are unnecessary for A. deanei growth [1,9,39]. However, the absence of the endosymbiont influenced the content of lipid components such as ergosterol and fatty acids. McLaughlin [40] did not detect significant changes in fatty acid composition of symbiotic and aposymbiotic cultures of Crithidia oncopeleti, although high percentages of unsaturated fatty acids were observed in lipid extracts from Blastocrithidia culicis cultures.
A comparison of the 0% Log and 0% Sta cultures (both without FBS) reveals that only the CFA levels increased significantly for the second condition, and the main saturated fatty acids decreased proportionally for the stationary cultures. This result contrasts with that of Bronia [41], who observed an increase in saturated fatty acids and a decrease in mainly polyunsaturated fatty acids in Trypanosoma cruzi stationary growth phase cultures.
An analysis of the fatty acid composition of A. deanei lipid extracts revealed the main fatty acids to be C18:3, C18:1, iso C17:0, C18:2 and C18:0, which represent nearly 75% of all fatty acids in the four growth conditions analyzed (Table S1). Korn et al. [42] described the fatty acid composition of Crithidia sp. (grown in a different culture media) and detected mainly C18:1 (18%), C18:2 (16%) and C18:3 (21%) fatty acids in the lipid extracts. They also found 16% of a (probably) branched-chain fatty acid in Crithidia sp., which was not characterized. Similarly, Meyer and Holz [43] showed that the main fatty acids found in some members of the Kinetoplastid order (including Crithidia species) were C18:1, C18:2 and C18:3. These authors first reported the presence of CFA C19:0 in Crithidia and detected a high percentage (20%) of iso 17:0 fatty acid in the trypanosomatid Leishmania tarentolae.
No studies involving the total fatty acid composition of A. deanei were found in the literature. The results presented here are the first to reveal and detect a high iso C17:0 fatty acid content in Crithidia (formerly A. deanei) genera. As mentioned above, a high C17:0 fatty acid content was detected in L. tarentolae and a low C17:0 fatty acid content in epimastigote, trypomastigote and amastigote forms of T. cruzi [44]. The authors also identified and quantified other branched-chain fatty acids such as C13, C14, C15, C16 and C19, although smaller amounts of these are present in the lipid extracts. Iso- and anteiso- branched-chain fatty acids are present in the lipid membrane of several bacteria species [45], as well as in some genera, such as Bacillus and Listeria; they are associated with adaptations involving the maintenance of membrane fluidity at low temperatures or alkaline/acidic pH conditions [46,47]. Although it has been suggested that iso- and anteiso- branched-chain fatty acids play important functions in the development of some Eukaryote models [48], the unusually high iso-C17:0 branched-chain fatty acid may represent a strategy used by A. deanei to adapt to specific characteristics of insect hosts [49].
In their study involving the presence of CFA C19:0 (cis-9,10-methyleneoctadecanoic acid) among many species of the Trypanosomatidae family, Fish et al. [25] detected a value of 44% of the CFA in lipid extract from A. deanei cultures, thus concluding that this CFA does not seem to be associated with the endosymbiont. Their data were confirmed by the present study, which detected CFA even in aposymbiotic A. deanei (Table 1 and Table S1). The presence of the cis-11,12 isomer was also observed for the four conditions analyzed.
C19:0 CFA values for the lipid extracts from A. deanei obtained during the logarithmic growth phase were considerably smaller than those of the lipid extract obtained from the same organism at the stationary growth phase. Similarly, a high CFA 17:0 percentage was detected in Pseudomonas putida at the stationary phase but not at the logarithmic phase [50]. CFAs seem to stabilize membrane lipids against turnover and degradation; in addition, some reports indicate that they contribute to stress tolerance [50,51]. Generally, organisms in which CFAs are found contain greater contents during the later stages of growth [51], thus corroborating the results obtained in the present study. The hypothesis that a correlation exists between high CFA content and stress resistance was tested in E. coli., the results showing that CFA synthetase mutants exhibited the same behavior as wild bacteria with regard to stress resistance [25,52]. Therefore, the function of CFA in bacteria and trypanosomatids remains uncertain.
The results for the sterols from A. deanei grown in the absence and presence of FBS (Supplementary Table S5) agreed with those published by Korn et al. [53], who observed that Crithidia fasciculata contained mainly ergosterol when grown in the absence of FBS, small quantities (6%) of cholesterol when grown in the presence of 20% of FBS and 25% of another sterol (probably 22,23-dihydroergosterol) when grown in the presence of FBS. Palmié-Peixoto et al. [14], who analyzed sterols of A. deanei grown with 10% FBS, detected only ergosterol (99%). Fecosterol, methyl-episterol and 4-methyl-ergosta-8,14,24-trienol (detected in this work) were not detected by the authors mentioned above. A. deanei is not able to synthesize cholesterol, and a high cholesterol uptake from the culture medium was described for another member of Trypanosomatidae family: T. cruzi. Epimastigote forms of T. cruzi are able to store and mobilize high amounts of cholesterol (52.9%) in reservosome lipid inclusions; however, the cellular implications of lipid uptake are unknown [54]. The present study found that A. deanei grown in the presence of FBS exhibits a cholesterol uptake of around 7.8%, a behavior that was slightly affected (increasing to 11.4%) by the removal of the symbiont (Supplementary Table S5). The high ergosterol content observed for aposymbiotic A. deanei, together with the high levels of monounsaturated fatty acids, may suggest compensation of the liquid-ordered state of the membrane at physiological temperatures [55].
The authors of this study were able to successfully identify and quantify fatty acids from lipid extracts of A. deanei grown in different nutritional and physiological conditions. The GC-MS analysis revealed unusual branched-chain and cyclic fatty acids and provided some potential insights regarding the function of these fatty acids on the physiology of this trypanosomatid. 1H NMR analysis confirmed these results, and 2D 1H-13C HSQC multiplicity-edited NMR analysis provided correlation maps that allowed for the identification of intact lipid classes in the extracts. These NMR fingerprints will greatly aid other research involving lipid analysis in trypanosomatids. Principal component analysis using 1H NMR was shown to be a useful tool for identifying the main contributors that distinguish the lipid extracts from various physiological and nutritional conditions. The GC-MS fatty acid profile presented in this work was the first to identify significant amounts of branched-chain fatty acids and cyclic fatty acid isomers in A. deanei lipid extracts, and it could point the biochemical synthetic pathways related to these molecules as targets for therapeutic research on protozoan parasites.
4. Materials and Methods
4.1. Parasite Culture
The choanomastigote form of A. deanei was cultivated in LIT medium [56] supplemented with 10% heat-inactivated fetal bovine serum (FBS) (or, for some experiments, in the absence of FBS) with penicillin and streptomycin, at 28 °C without agitation. Aposymbiotic A. deanei, kindly provided by Dr. Stenio Fragoso from the Instituto Carlos Chagas (Curitiba, Paraná, Brazil), was cultured in LIT media in the absence of FBS. Approximately 100 mL of cultures at 1 × 108 choanomastigotes/mL (early stationary phase) and 4 × 107 choanomastigotes/mL (logarithmic phase) were separated by centrifugation at 2000× g for 10 min, washed twice with PBS and frozen at −20 °C. All A. deanei cultures (except in the log phase experiment) were collected at a density of 1 × 108 choanomastigotes/mL. All cultivation experiments were performed in triplicate.
4.2. Lipid Extraction
Parasite pellets were subjected to lysis, being frozen with liquid nitrogen (−196 °C) and returned to a water bath (37 °C) three times, after which they were freeze-dried overnight. A predetermined amount (20 mg for culture) was subsequently transferred to a glass tube with a screw cap and teflon. The parasites were then extracted using a (1:1 v/v) chloroform–methanol mixture (both obtained from Tedia, Fairfield, OH, USA) at 100 °C for 2 h [57,58], after which the lipid extracts were centrifuged at 8000× g for 15 min, the organic solvent in the supernatant was collected and the analyses were performed.
4.3. Nuclear Magnetic Resonance (NMR)
After drying by a N2 stream, the samples were deuterium-exchanged by repeated dissolution in MeOD-D2O (2:1) and freeze-dried. Their spectra were obtained from solutions in MeOD-CDCl3 (1:1) at 30 °C, using tetramethylsilane (TMS) as reference (δ = 0). All spectra were obtained using either a Bruker 400 MHz Avance III NMR spectrometer with a 5 mm BBI inverse probe with a Z gradient or a Bruker 600 MHz Ascend NMR spectrometer with a 5 mm QXI inverse probe with a Z gradient. One-dimensional 1H-NMR was performed using 64 scans to give a Signal/Noise (S/N) ratio of at least 2000/1 (90° pulse, relaxation delay = 4.0 s, number of time domain points = 65,536 and acquisition time = 7.7 s); experiments were performed without tube rotation, with the TMS signal at a medium width varying from 0.8 to 1.0 Hz, the 1H measurements being obtained in triplicate. Two-dimensional NMR experiments were carried out using edited-HSQC, heteronuclear correlation via double inept transfer with decoupling during acquisition, using trim pulses in inept transfer with multiplicity editing during the selection step (hsqcedetgpsisp2.2), TOCSY, total homonuclear correlation via Hartman-Hahn transfer using the MLEV17 sequence for mixing, using a mixing time of 0.06 s (mlevphpr.2) and HMBC heteronuclear correlation via zero and double quantum coherence, optimized on long-range couplings’ (hmbcgplpndqf) pulse sequences. The 2D experiments were recorded for quadrature detection in the indirect dimension, and edited-HSQC spectra were acquired using 128 scans per series of 1 K × 256 W data points with zero filling in F1 (4 K) prior to Fourier transformation [28,59]. Data processing and integration were performed using the software TOPSPIN 3.1 (Bruker Biospin, Rheinstetten, Germany).
4.4. Principal Component Analysis (PCA) and Data Reduction
After the NMR spectral phase and baseline correction, each spectrum was data-reduced to 440 regions of equal width (0.01 ppm) using the AMIX (Analysis of MIXtures) software package, version 3.8 (Bruker Biospin, Rheinstetten, Germany). The spectral width considered was from 0.5 to 6.50 ppm. The spectral region close to the CHD2OD resonances (δ 3.32–3.36) was removed from all data sets prior to normalization and multivariate data analysis in order to eliminate variation due to water suppression efficiency or solvent multiplicity in homogeneities. Following a preliminary PCA, the regions corresponding to R-CH3 (δ 0.84–0.91), (CH2)n (δ 1.2–1.4), Ala-Cβ (δ 1.50–1.57), succinate (δ 2.58–2.61) and N+(CH3)3 (δ 3.19–3.25) resonances were also removed from some of the spectra, mainly because minor peak shape deformations in this region interfere with PCA due to the high intensity of the signals in this region. All remaining frequency regions of the spectra were analyzed with Pareto scaling.
4.5. Lipid Derivatizations
Aliquots of 100 µL of samples dissolved in CHCl3-MeOH (1:1, v/v) were first dried under a gentle N2 stream, after which methanolysis was carried out with 1N methanolic HCl (Sigma-Aldrich, St. Louis., MO, USA) at 100 °C for 2 h to obtain the fatty acid methyl esters (FAMEs). The resulting FAMEs were extracted by a partition between n-hexane (1 mL) and distilled water (0.5 mL). The organic (upper) phase was collected and evaporated under a stream of N2. In order to analyze the sterols, 50 µL of the lipid extract was dried under a gentle N2 stream and derivatized for 30 min in a sealed vial with N,N-bis(trimethylsilyl)-2,2,2-trifluoroacetamide (BSTFA)-pyridine (2:1, v/v) (Sigma-Aldrich) at 100 °C. The trimethylsilyl derivatives were then subjected to GC-MS analysis.
4.6. Gas Chromatography Coupled with Mass Spectrometry (GC-MS)
GC-MS analysis was carried out using a Varian 3800 gas chromatograph coupled to a 4000 MS detector (ion trap), equipped with a 30 m × 0.25 mm i.d. low-bleed/MS capillary column (VF-5ms) (both from Agilent/Varian, Santa Clara, CA, USA). The temperature ramp to fatty acid analysis was as follows: injector 250 °C, oven initially at 100 °C, maintained for 2 min, heated to 280 °C (5 °C/min) and then maintained for 2 min. The temperature ramp was altered as follows for sterol analysis: injector 250 °C, column maintained at 200 °C for 1 min, increased to 300 °C at a rate of 15 °C/min and finally maintained at 300 °C for 15 min [60]. For all analyses, the injection volume was 1 µL, with a split ratio of 1:10. Post-run analysis was performed using a Saturn Workstation 5.1 [61]. GC-MS experiments were performed in triplicate.
4.7. Cell Microscopy
Coanomastigotes grown under different conditions were fixed with methanol at room temperature for 15 min and stained with Giemsa. The slides were then mounted and analyzed under an Olympus microscope equipped with a 100× Oil objective.
4.8. Data Analysis
Raw data values obtained from 1H NMR integration and GC-MS chromatograms were analyzed using the data analysis tool from Microsoft Excel 2019 software. The TMS signal was normalized to the value of 1.00 on all 1H NMR spectra, and the relative fatty acid proportion was calculated by means of the integration and sum of the peaks corresponding to all fatty acids on each GC-MS chromatogram, normalized to 100%. Unless otherwise stated, the data in the tables are shown as mean minus/plus standard deviation. The statistical significance of the differences among growth conditions was calculated using one-way ANOVA. The p-values obtained were adjusted to account for multiple comparisons.
Supplementary Materials
The following supporting information may be downloaded at https://www.mdpi.com/article/10.3390/molecules29143352/s1: Table S1: Changes in fatty acid methyl ester ratios obtained from lipid extracts from A. deanei under the four growth conditions; Table S2: 1H NMR and 13C NMR lipid assignments performed on the lipid extracts of A. deanei; Table S3: 1H NMR lipid assignments and integration values from lipid extracts from A. deanei under the four growth conditions; Figure S1: Scores plot of PCA performed using 1H NMR spectra from lipid extracts of A. deanei under four experimental conditions; Table S4. Statistical significance of relative levels of Ergosterol obtained through integration of 1H NMR chemical shift corresponding to lipid assignment on each growth condition; Table S5: The sterol composition percentages of A. deanei grown in the presence and absence of FBS and in the aposymbiotic condition.
Author Contributions
Conceptualization, G.L.S. and W.D.D.; methodology, A.J.P., L.A.L., N.S.-M. and A.P.S.-F.; software, A.P.S.-F.; writing—original draft preparation, A.J.P.; writing—review and editing, A.P.S.-F. and G.L.S.; funding acquisition, G.L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CNPq, grant number 311177/2021-2.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available upon request due to privacy.
Acknowledgments
The authors would like to thank the Brazilian funding agencies Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and the UFPR NMR Center.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Motta, M.C.M.; Catta-Preta, C.M.C.; Schenkman, S.; Martins, A.C.d.A.; Miranda, K.; de Souza, W.; Elias, M.C. The Bacterium Endosymbiont of Crithidia deanei Undergoes Coordinated Division with the Host Cell Nucleus. PLoS ONE 2010, 5, e12415. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; Borghesan, T.C.; Ferreira, R.C.; Santos, M.A.; Takata, C.S.; Campaner, M.; Nunes, V.L.; Milder, R.V.; de Souza, W.; Camargo, E.P. Phylogenetic Validation of the Genera Angomonas and Strigomonas of Trypanosomatids Harboring Bacterial Endosymbionts with the Description of New Species of Trypanosomatids and of Proteobacterial Symbionts. Protist 2011, 162, 503–524. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.; Ehret, G.; Poschmann, G.; Reinicke, T.; Maurya, A.K.; Kröninger, L.; Zanini, D.; Wolters, R.; Kalyanaraman, D.; Krakovka, M.; et al. Host-symbiont interactions in Angomonas deanei include the evolution of a host-derived dynamin ring around the endosymbiont division site. Curr. Biol. 2023, 33, 28–40.e7. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.S.; Catta-Preta, C.M.C.; Repolês, B.; Mottram, J.C.; De Souza, W.; Machado, C.R.; Motta, M.C.M. Importance of Angomonas Deanei KAP4 for KDNA Arrangement, Cell Division and Maintenance of the Host-Bacterium Relationship. Sci. Rep. 2021, 11, 9210. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Hoffmeister, M.; Rotte, C.; Henze, K. An Overview of Endosymbiotic Models for the Origins of Eukaryotes, Their ATP-Producing Organelles (Mitochondria and Hydrogenosomes), and Their Heterotrophic Lifestyle. Biol. Chem. 2001, 382, 1521–1539. [Google Scholar] [CrossRef]
- Du, Y.; McLaughlin, G.; Chang, K.P. 16S Ribosomal DNA Sequence Identities of β-Proteobacterial Endosymbionts in Three Crithidia Species. J. Bacteriol. 1994, 176, 3081–3084. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Du, Y.; A Maslov, D.; Chang, K.P. Monophyletic origin of β-division proteobacterial endosymbionts and their coevolution with insect trypanosomatid protozoa Blastocrithidia culicis and Crithidia spp. Proc. Natl. Acad. Sci. USA 1994, 91, 8437–8441. [Google Scholar] [CrossRef] [PubMed]
- Motta, M.C.M.; Martins, A.C.d.A.; de Souza, S.S.; Catta-Preta, C.M.C.; Silva, R.; Klein, C.C.; de Almeida, L.G.P.; Cunha, O.d.L.; Ciapina, L.P.; Brocchi, M.; et al. Predicting the Proteins of Angomonas deanei, Strigomonas culicis and Their Respective Endosymbionts Reveals New Aspects of the Trypanosomatidae Family. PLoS ONE 2013, 8, e60209. [Google Scholar] [CrossRef]
- Salzman, T.A.; Batlle, A.D.C.; Angluster, J.; de Souza, W. Heme synthesis in Crithidia deanei: Influence of the endosymbiote. Int. J. Biochem. 1985, 17, 1343–1347. [Google Scholar] [CrossRef]
- Alves, J.M.P.; Voegtly, L.; Matveyev, A.V.; Lara, A.M.; da Silva, F.M.; Serrano, M.G.; Buck, G.A.; Teixeira, M.M.G.; Camargo, E.P. Identification and Phylogenetic Analysis of Heme Synthesis Genes in Trypanosomatids and Their Bacterial Endosymbionts. PLoS ONE 2011, 6, e23518. [Google Scholar] [CrossRef]
- Camargo, E.P.; Freymuller, E. Endosymbiont as supplier of ornithine carbamoyltransferase in a trypanosomatid. Nature 1977, 270, 52–53. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Chang, C.S.; Sassa, S. Heme biosynthesis in bacterium-protozoon symbioses: Enzymic defects in host hemoflagellates and complemental role of their intracellular symbiotes. Proc. Natl. Acad. Sci. USA 1975, 72, 2979–2983. [Google Scholar] [CrossRef] [PubMed]
- Motta, M.C.M.; Leal, L.H.M.; DE Souza, W.; DE Almeida, D.F.; Ferreira, L.C.S. Detection of Penicillin-binding Proteins in the Endosymbiont of the Trypanosomatid Crithidia deanei. J. Eukaryot. Microbiol. 1997, 44, 492–496. [Google Scholar] [CrossRef]
- Palmié-Peixoto, I.V.; Rocha, M.R.; Urbina, J.A.; Souza, W.; Einicker-Lamas, M.; Motta, M.C.M. Effects of sterol biosynthesis inhibitors on endosymbiont-bearing trypanosomatids. FEMS Microbiol. Lett. 2006, 255, 33–42. [Google Scholar] [CrossRef] [PubMed]
- de Azevedo-Martins, A.C.; Frossard, M.L.; de Souza, W.; Einicker-Lamas, M.; Motta, M.C.M. Phosphatidylcholine synthesis in Crithidia deanei: The influence of the endosymbiont. FEMS Microbiol. Lett. 2007, 275, 229–236. [Google Scholar] [CrossRef][Green Version]
- De Freitas-Junior, P.R.; Catta-Preta, C.M.; Andrade, I.d.S.; Cavalcanti, D.P.; Souza, W.; Einicker-Lamas, M.; Motta, M.C.M. Effects of miltefosine on the proliferation, ultrastructure, and phospholipid composition of Angomonas deanei, a trypanosomatid protozoan that harbors a symbiotic bacterium. FEMS Microbiol. Lett. 2012, 333, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Coen, M.; Lenz, E.M.; Nicholson, J.K.; Wilson, I.D.; Pognan, F.; Lindon, J.C. An Integrated Metabonomic Investigation of Acetaminophen Toxicity in the Mouse Using NMR Spectroscopy. Chem. Res. Toxicol. 2003, 16, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.; Puppa, M.; van der Merwe, M.; Tipirneni-Sajja, A. Rapid and automated lipid profiling by nuclear magnetic resonance spectroscopy using neural networks. NMR Biomed. 2023, 36, e5010. [Google Scholar] [CrossRef] [PubMed]
- Castañé, H.; Baiges-Gaya, G.; Hernández-Aguilera, A.; Rodríguez-Tomàs, E.; Fernández-Arroyo, S.; Herrero, P.; Delpino-Rius, A.; Canela, N.; Menendez, J.A.; Camps, J.; et al. Coupling Machine Learning and Lipidomics as a Tool to Investigate Metabolic Dysfunction-Associated Fatty Liver Disease. A General Overview. Biomolecules 2021, 11, 473. [Google Scholar] [CrossRef]
- Dubois, N.; Barnathan, G.; Gouygou, J.; Bergé, J. Gas chromatographic behavior of fatty acid derivatives for mass spectrometry on low-polarity capillary columns. Eur. J. Lipid Sci. Technol. 2009, 111, 688–697. [Google Scholar] [CrossRef]
- Haken, J. Retention time relationships in the gas chromatography of the methyl esters of branched chain fatty acids. J. Chromatogr. A 1967, 26, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Hauff, S.; Vetter, W. Quantification of Branched Chain Fatty Acids in Polar and Neutral Lipids of Cheese and Fish Samples. J. Agric. Food Chem. 2010, 58, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Ran-Ressler, R.R.; Lawrence, P.; Brenna, J.T. Structural characterization of saturated branched chain fatty acid methyl esters by collisional dissociation of molecular ions generated by electron ionization. J. Lipid Res. 2012, 53, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Oudejans, R.C.H.M.; Van der Horst, D.J.; Van Dongen, J.P.C.M. Isolation and identification of cyclopropane fatty acids from the millipede Graphidostreptus tumuliporus (Myriapoda:Diplopoda). Biochemistry 1971, 10, 4938–4941. [Google Scholar] [CrossRef] [PubMed]
- Fish, W.R.; Holz, G.G.; Beach, D.H.; Owen, E.; Anekwe, G.E. The cyclopropane fatty acid of trypanosomatids. Mol. Biochem. Parasitol. 1981, 3, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.; Reiser, R. Cyclopropane fatty acid metabolism: Physical and chemical identification of propane ring metabolic products in the adipose tissue. J. Am. Oil Chem. Soc. 1965, 42, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Nádvorníková, J.; Pitthard, V.; Kurka, O.; Kučera, L.; Barták, P. Egg vs. Oil in the Cookbook of Plasters: Differentiation of Lipid Binders in Wall Paintings Using Gas Chromatography–Mass Spectrometry and Principal Component Analysis. Molecules 2024, 29, 1520. [Google Scholar] [CrossRef] [PubMed]
- de Santana-Filho, A.P.; Jacomasso, T.; Riter, D.S.; Barison, A.; Iacomini, M.; Winnischofer, S.M.B.; Sassaki, G.L. NMR metabolic fingerprints of murine melanocyte and melanoma cell lines: Application to biomarker discovery. Sci. Rep. 2017, 7, srep42324. [Google Scholar] [CrossRef]
- Benitez, D.; Pezaroglo, H.; Martínez, V.; Casanova, G.; Cabrera, G.; Galanti, N.; González, M.; Cerecetto, H. Study of Trypanosoma cruzi epimastigote cell death by NMR-visible mobile lipid analysis. Parasitology 2012, 139, 506–515. [Google Scholar] [CrossRef]
- Sanchez-Moreno, M.; Fernandez-Becerra, M.; Castilla-Calvente, J.; Osuna, A. Metabolic studies by 1H NMR of different forms of Trypanosoma cruzi as obtained by ‘in vitro’ culture. FEMS Microbiol. Lett. 1995, 133, 119–125. [Google Scholar] [CrossRef]
- Gilroy, F.V.; Edwards, M.R.; Norton, R.S.; O’Sullivan, W.J. Metabolic studies of the protozoan parasite, Crithidia luciliae, using proton nuclear magnetic resonance spectroscopy. Mol. Biochem. Parasitol. 1988, 31, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.d.L.; Buldain, G.; Frydman, B.; Cannata, J.J.B.; Cazzulo, J.J. Carbon-13 Nuclear Magnetic Resonance Analysis of [1-13C] Glucose Metabolism in Crithidia Fasciculata Evidence of CO2 Fixation by Phosphoenolpyruvate Carboxykinase. Eur. J. Biochem. 1985, 149, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Frydman, B.; Santos, C.d.L.; Cannata, J.J.B.; Cazzulo, J.J. Carbon-13 Nuclear Magnetic Resonance Analysis of [1-13C]Glucose Metabolism in Trypanosoma Cruzi Evidence of the Presence of Two Alanine Pools and of Two CO2 Fixation Reactions. Eur. J. Biochem. 1990, 192, 363–368. [Google Scholar] [CrossRef]
- Penin, P.; Sanchez-Moreno, M.; de Diego, J. Proton nuclear magnetic resonance analysis of metabolic end products of the Bolivia strain of Trypanosoma cruzi and three of its clones. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 120, 571–574. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, X.; Olson, C.L.; Engman, D.M.; Ames, J.B. NMR structure of the calflagin Tb24 flagellar calcium binding protein of Trypanosoma brucei. Protein Sci. 2012, 21, 1942–1947. [Google Scholar] [CrossRef] [PubMed]
- Miralles, D.; Marın, C.; Magán, R.; Fernández-Ramos, C.; Entrala, E.; Cordova, O.; Vargas, F.; Sánchez-Moreno, M. In vitro culture and biochemical characterization of six trypanosome isolates from Peru and Brazil. Exp. Parasitol. 2002, 102, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Moreno, B.; Urbina, J.A.; Oldfield, E.; Bailey, B.N.; Rodrigues, C.O.; Docampo, R. 31P NMR Spectroscopy of Trypanosoma Brucei, Trypanosoma Cruzi, and Leishmania Major. Evidence for High Levels of Condensed Inorganic Phosphates. J. Biol. Chem. 2000, 275, 28356–28362. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Utzinger, J.; Saric, J.; Li, J.V.; Burckhardt, J.; Dirnhofer, S.; Nicholson, J.K.; Singer, B.H.; Brun, R.; Holmes, E. Global metabolic responses of mice to Trypanosoma brucei brucei infection. Proc. Natl. Acad. Sci. USA 2008, 105, 6127–6132. [Google Scholar] [CrossRef] [PubMed]
- Mundim, M.H.; Roitman, I.; Hermans, M.A.; Kitajima, E.W. Simple Nutrition of Crithidia deanei, a Reduviid Trypanosomatid with an Endosymbiont. J. Protozool. 1974, 21, 518–521. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, G.L.; Wood, D.L.; Cain, G.D. Lipids and carbohydrates in symbiotic and aposymbiotic Crithidia oncopelti and Blastocrithidia culicis. Comp. Biochem. Physiol. Part B Comp. Biochem. 1983, 76, 143–152. [Google Scholar] [CrossRef]
- Bronia, D.H.; Aguerri, A.M.; Bertetto, S.T.; Broña, D.H.; Aguerri, A.M.; Bertetto, A. Trypanosoma cruzi: Changes in lipid composition during aging in culture. Exp. Parasitol. 1986, 61, 151–159. [Google Scholar] [CrossRef]
- Korn, E.D.; Greenblatt, C.L.; Lees, A.M. Synthesis of unsaturated fatty acids in the slime mold Physarum polycephalum and the zooflagellates Leishmania tarentolae, Trypanosoma lewisi, and Crithidia sp.: A comparative study. J. Lipid Res. 1965, 6, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Meyers, H.; Holz, G.G. Biosynthesis of Lipids by Kinetoplastid Flagellates. J. Biol. Chem. 1966, 241, 5000–5007. [Google Scholar] [CrossRef]
- Kaneda, Y.; Nagakura, K.; Goutsu, T. Lipid composition of three morphological stages of Trypanosoma cruzi. Comp. Biochem. Physiol. Part B Comp. Biochem. 1986, 83, 533–536. [Google Scholar] [CrossRef]
- Kaneda, T. Iso-and Anteiso-Fatty Acids in Bacteria: Biosynthesis, Function, and Taxonomic Significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef]
- Mansilla, M.C.; Cybulski, L.E.; Albanesi, D.; De Mendoza, D. Control of Membrane Lipid Fluidity by Molecular Thermosensors Temperature-Dependent Changes in the Content of UFAS of Glycerophospholipids In E. coli. J. Bacteriol. 2004, 186, 6681–6688. [Google Scholar] [CrossRef] [PubMed]
- Giotis, E.S.; McDowell, D.A.; Blair, I.S.; Wilkinson, B.J. Role of Branched-Chain Fatty Acids in pH Stress Tolerance in Listeria monocytogenes. Appl. Environ. Microbiol. 2007, 73, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Kniazeva, M.; Crawford, Q.T.; Seiber, M.; Wang, C.-Y.; Han, M. Monomethyl Branched-Chain Fatty Acids Play an Essential Role in Caenorhabditis elegans Development. PLoS Biol. 2004, 2, e257. [Google Scholar] [CrossRef]
- Holz, G.G.; Beach, D.H.; Singh, B.N.; Fish, W.R. Biosynthesis of the novel fatty acid, 17-methyl-cis-9,10-methyleneoctadecanoic acid, by the parasitic protozoan, Herpetomonas megaseliae. Lipids 1983, 18, 607–610. [Google Scholar] [CrossRef]
- Muñoz-Rojas, J.; Bernal, P.; Duque, E.; Godoy, P.; Segura, A.; Ramos, J.-L. Involvement of Cyclopropane Fatty Acids in the Response of Pseudomonas putida KT2440 to Freeze-Drying. Appl. Environ. Microbiol. 2006, 72, 472–477. [Google Scholar] [CrossRef]
- Macdonald, P.M.; Sykes, B.D.; McElhaney, R.N. Fluorine-19 nuclear magnetic resonance studies of lipid fatty acyl chain order and dynamics in Acholeplasma laidlawii B membranes. A direct comparison of the effects of cis- and trans-cyclopropane ring and double-bond substituents on orientational order. Biochemistry 1985, 24, 4651–4659. [Google Scholar] [CrossRef] [PubMed]
- Taylor, F.; E Cronan, J. Selection and properties of Escherichia coli mutants defective in the synthesis of cyclopropane fatty acids. J. Bacteriol. 1976, 125, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Korn, E.D.; Von Brand, T.; Tobie, E.J. The sterols of Trypanosoma cruzi and Crithidia fasciculata. Comp. Biochem. Physiol. 1969, 30, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.G.; Nakayasu, E.S.; Sant’Anna, C.; De Cicco, N.N.T.; Atella, G.C.; de Souza, W.; Almeida, I.C.; Cunha-E-Silva, N. Trypanosoma cruzi Epimastigotes Are Able to Store and Mobilize High Amounts of Cholesterol in Reservosome Lipid Inclusions. PLoS ONE 2011, 6, e22359. [Google Scholar] [CrossRef] [PubMed]
- Dufourc, E.J. Sterols and membrane dynamics. J. Chem. Biol. 2008, 1, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Camargo, E.P. Growth and Differentiation in Trypanosoma cruzi. I. Origin of Metacyclic Trypanosomes in Liquid Media. Rev. Inst. Med. Trop. Sao Paulo 1964, 6, 93–100. [Google Scholar] [PubMed]
- Cabrini, L.; Landi, L.; Stefanelli, C.; Barzanti, V.; Maria, S.A. Extraction of lipids and lipophilic antioxidants from fish tissues: A comparison among different methods. Comp. Biochem. Physiol. Part B Comp. Biochem. 1992, 101, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-H.; Liu, L.-Y.; Yang, M.-H.; Lee, M.-H. Ethyl Acetate/Ethyl Alcohol Mixtures as an Alternative to Folch Reagent for Extracting Animal Lipids. J. Agric. Food Chem. 2004, 52, 4984–4986. [Google Scholar] [CrossRef] [PubMed]
- Sassaki, G.L.; Riter, D.S.; Filho, A.P.S.; Guerrini, M.; Lima, M.A.; Cosentino, C.; Souza, L.M.; Cipriani, T.R.; Rudd, T.R.; Nader, H.B.; et al. A robust method to quantify low molecular weight contaminants in heparin: Detection of tris(2-n-butoxyethyl) phosphate. Analyst 2011, 136, 2330–2338. [Google Scholar] [CrossRef]
- Doyle, P.S.; Chen, C.-K.; Johnston, J.B.; Hopkins, S.D.; Leung, S.S.F.; Jacobson, M.P.; Engel, J.C.; McKerrow, J.H.; Podust, L.M. A Nonazole CYP51 Inhibitor Cures Chagas’ Disease in a Mouse Model of Acute Infection. Antimicrob. Agents Chemother. 2010, 54, 2480–2488. [Google Scholar] [CrossRef]
- Sassaki, G.L.; Souza, L.M.; Serrato, R.V.; Cipriani, T.R.; Gorin, P.A.; Iacomini, M. Application of acetate derivatives for gas chromatography–mass spectrometry: Novel approaches on carbohydrates, lipids and amino acids analysis. J. Chromatogr. A 2008, 1208, 215–222. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).