A Novel Kunitz Trypsin Inhibitor from Enterolobium gummiferum Seeds Exhibits Antibiofilm Properties against Pathogenic Yeasts

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
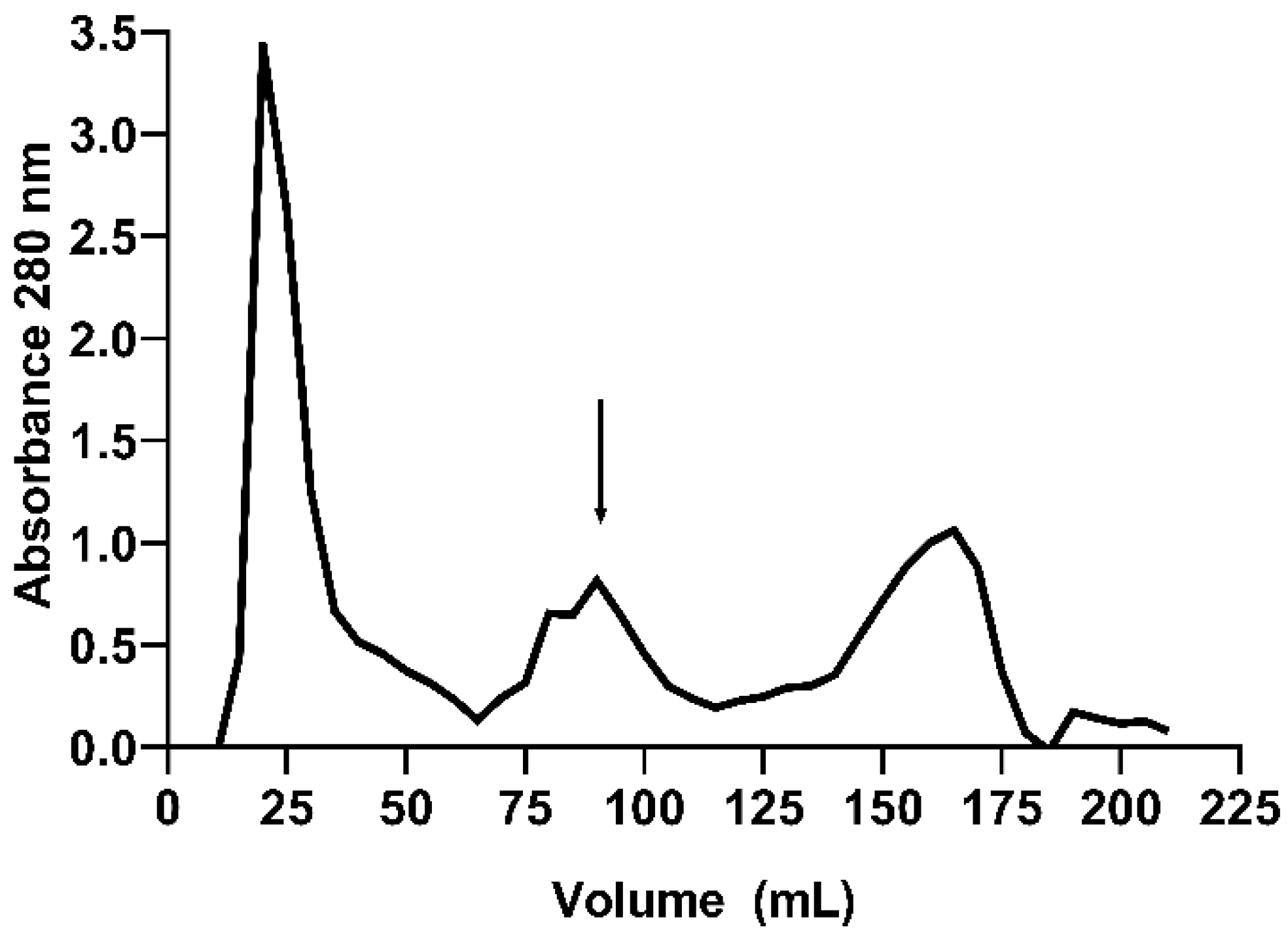
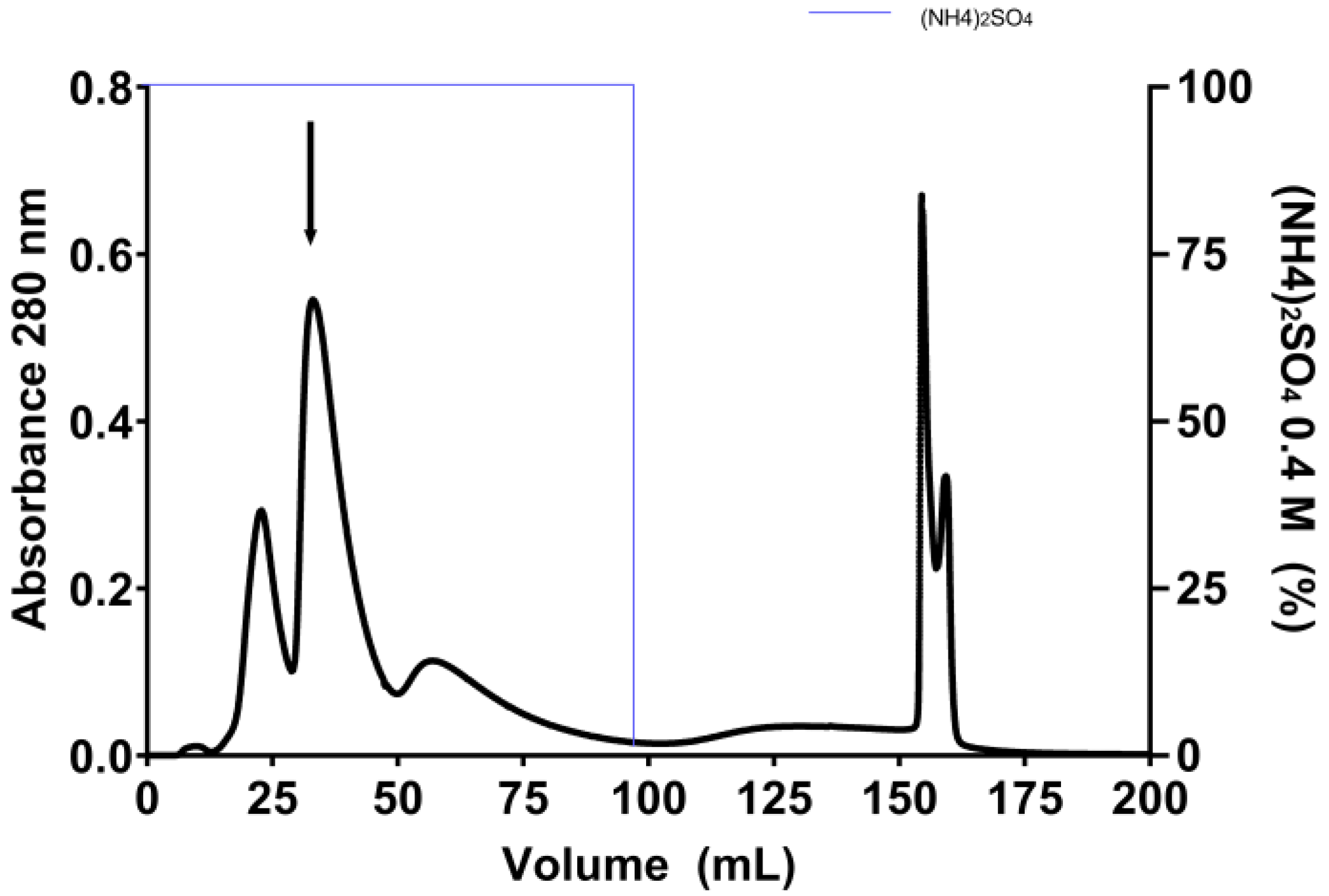
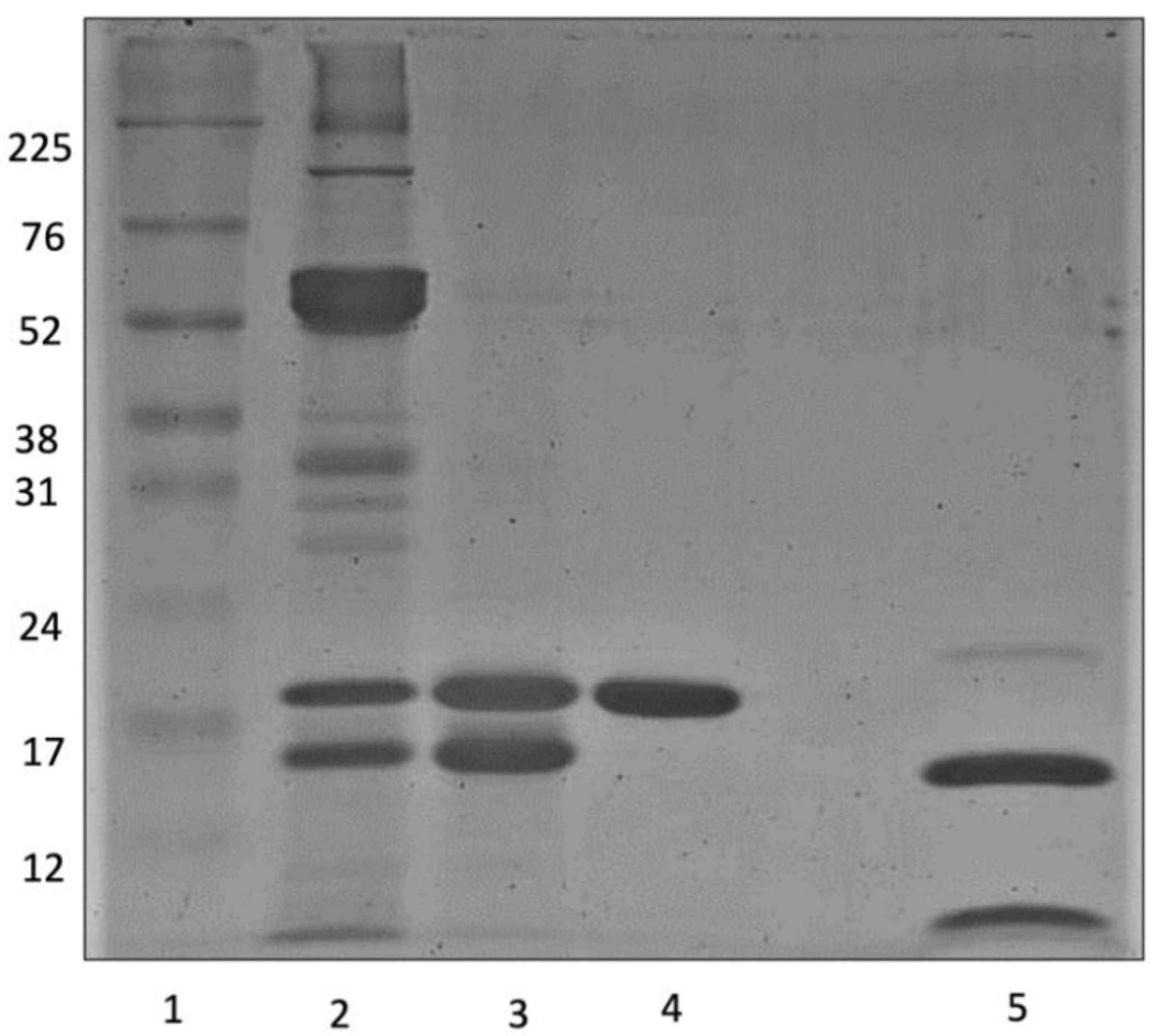
2.1. Purification of E. gummiferum Peptidase Inhibitor
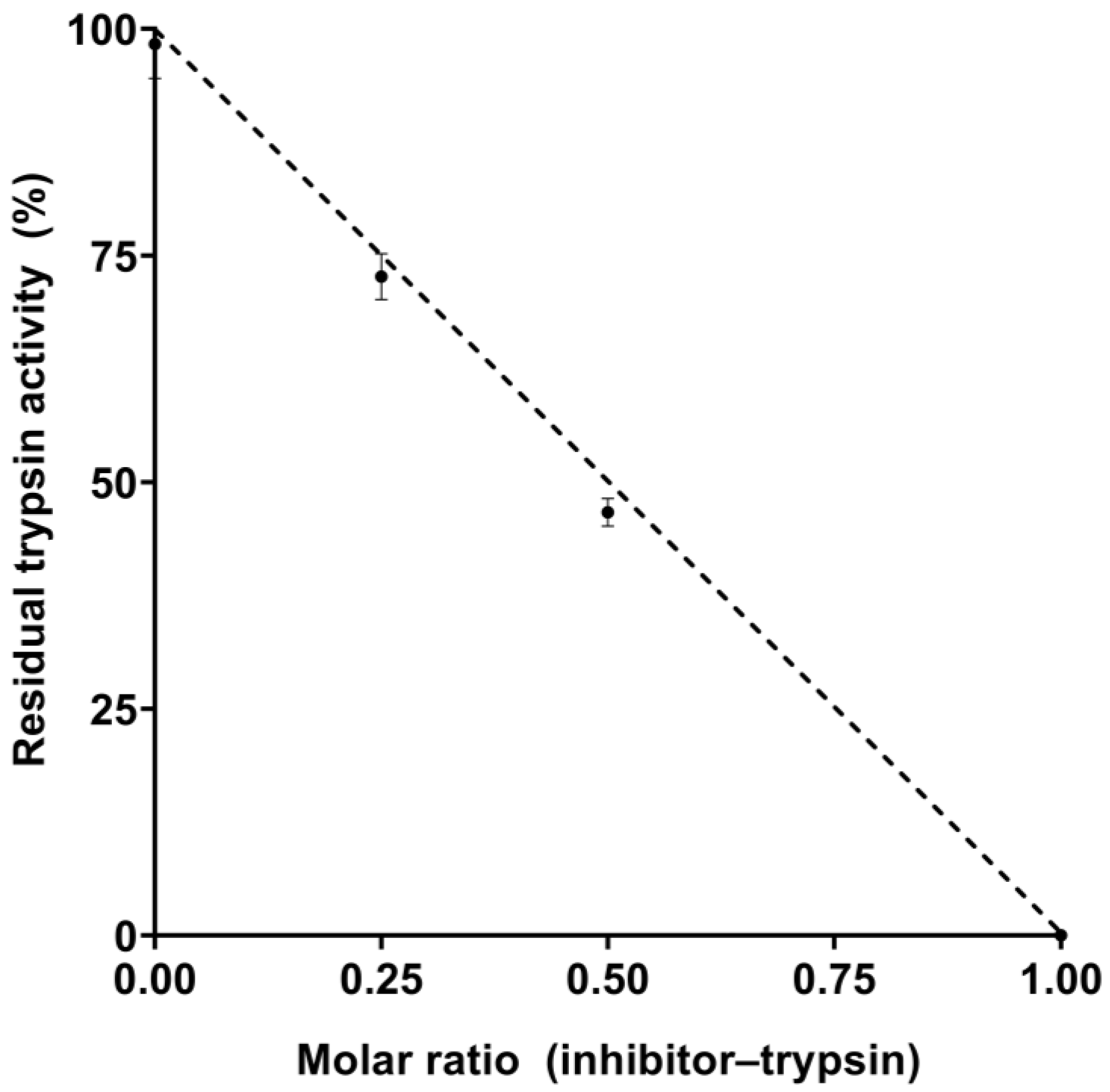
2.2. Inhibitory Properties of EgPI
2.3. Amino-Terminal Sequencing
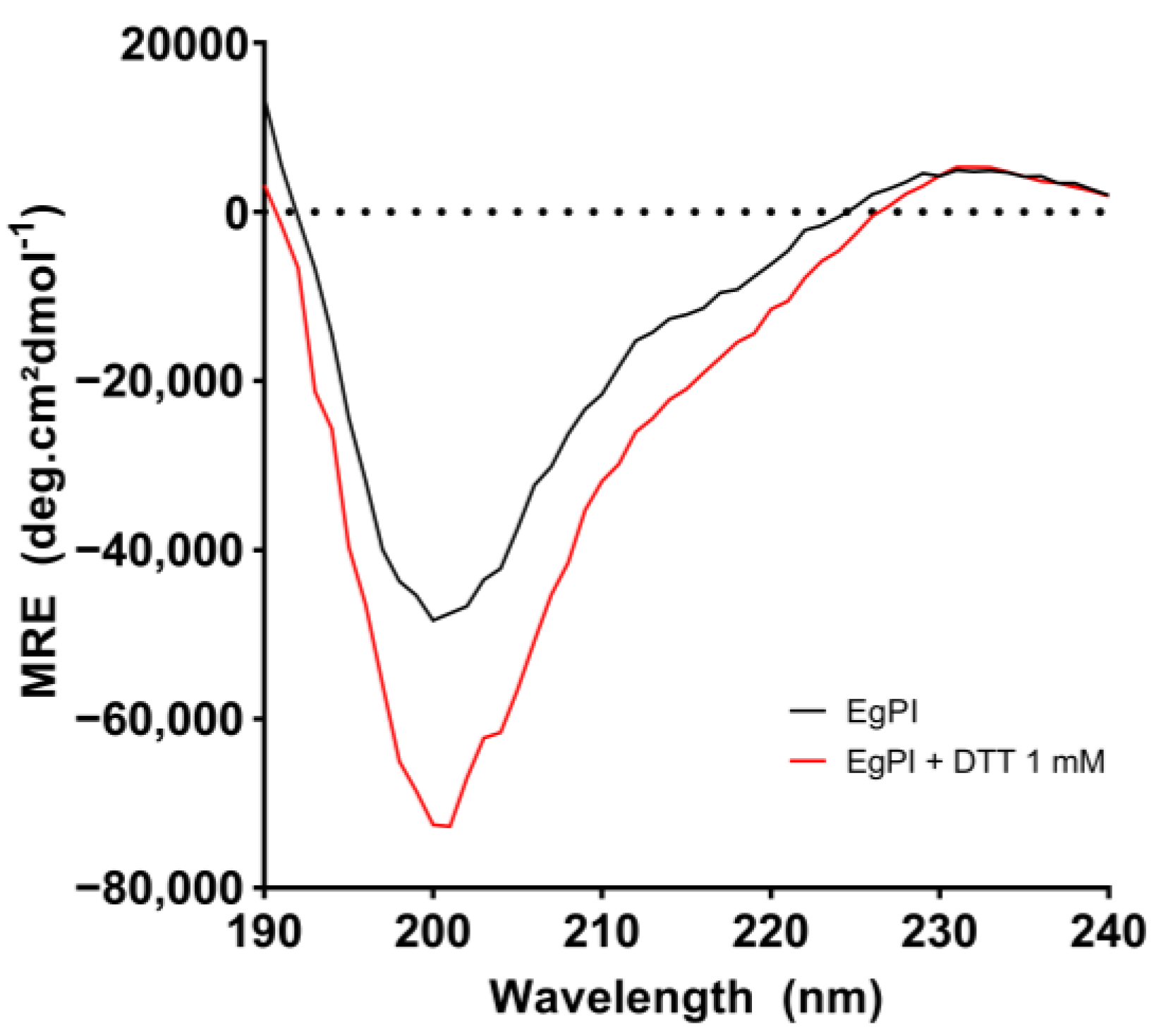
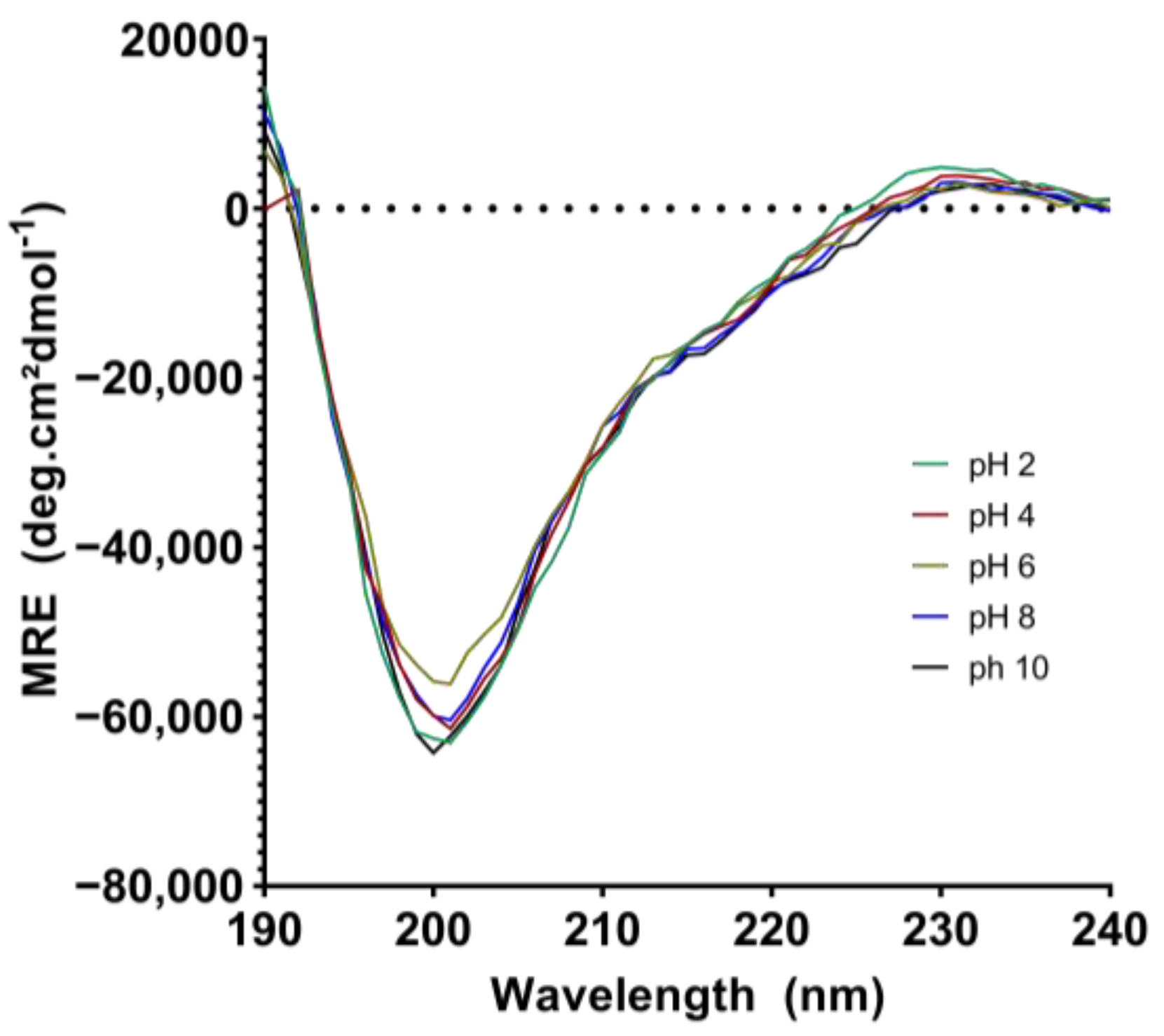
2.4. Analysis of the Secondary Structure and Stability of EgPI Using Circular Dichroism
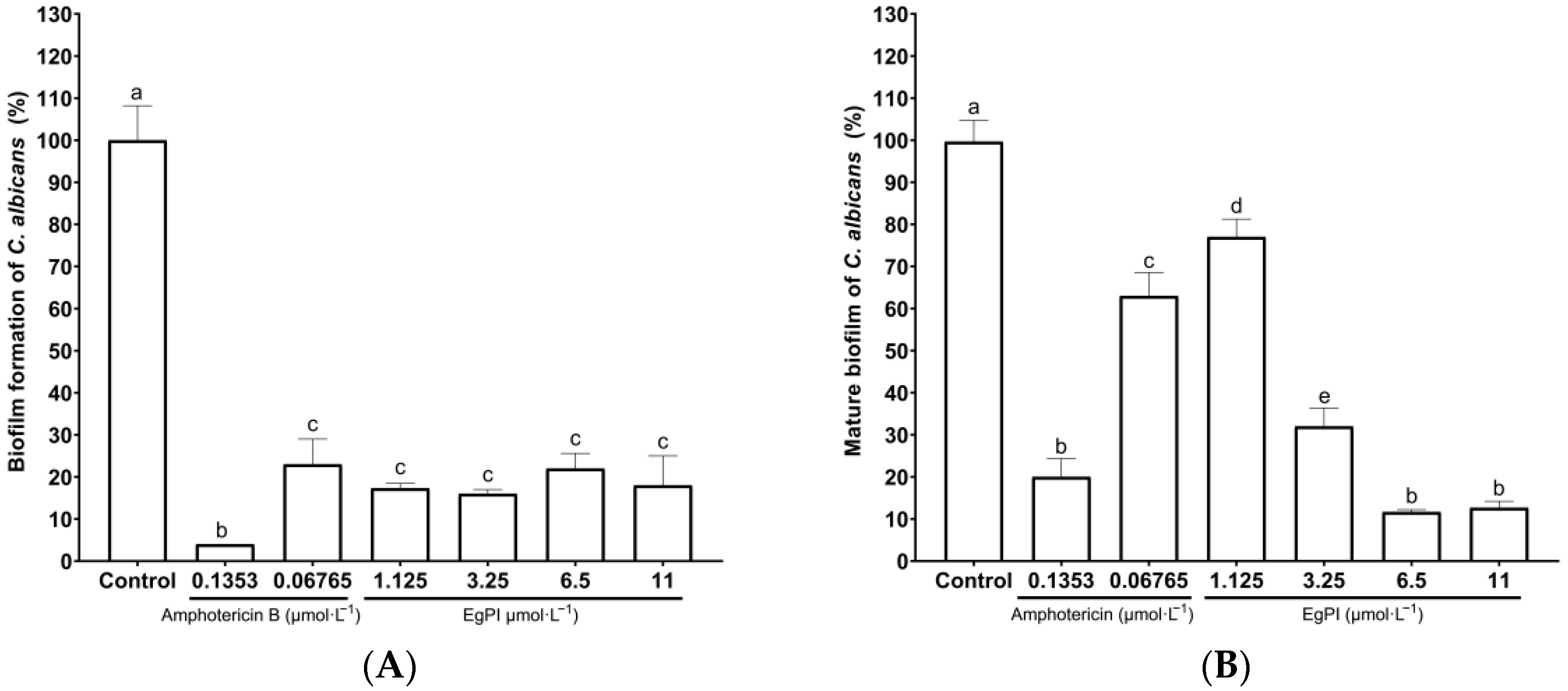
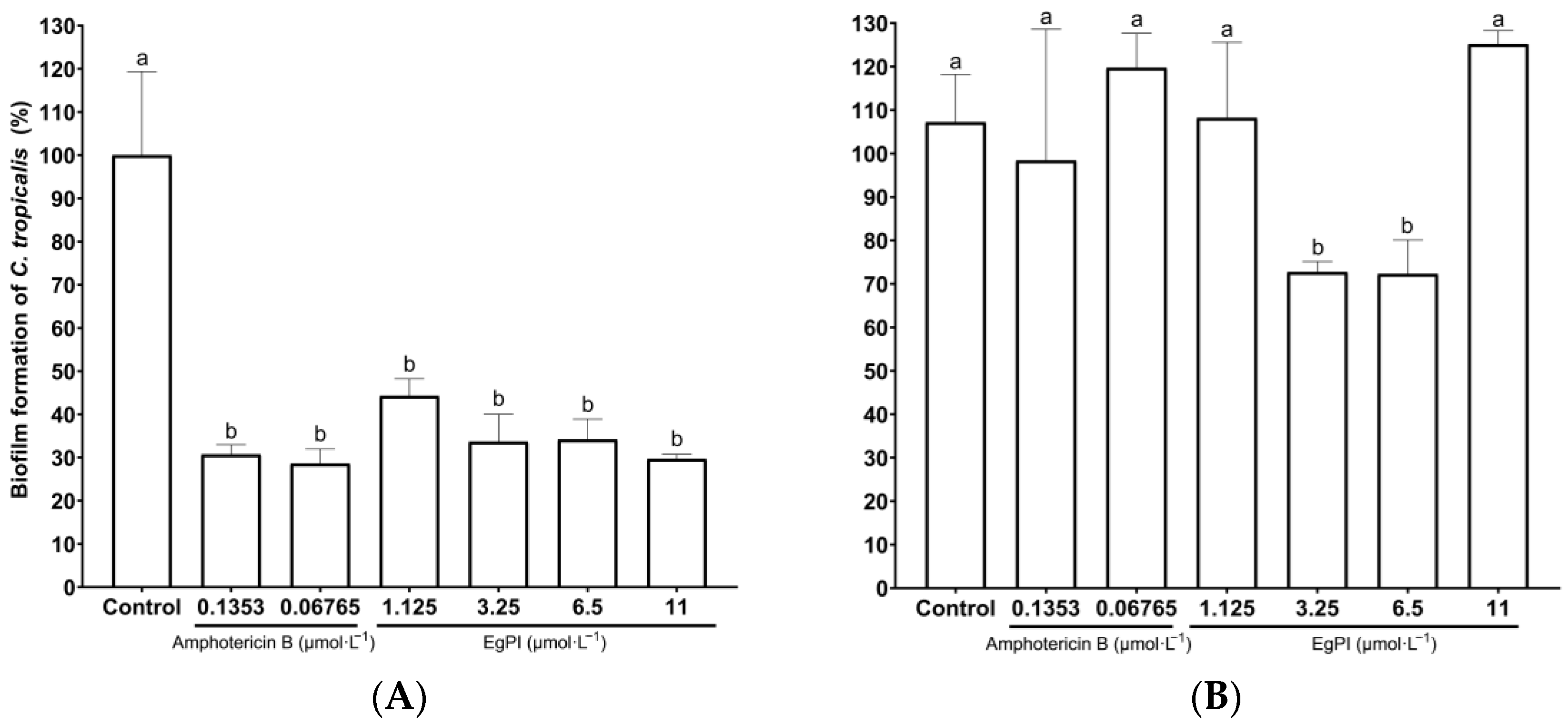
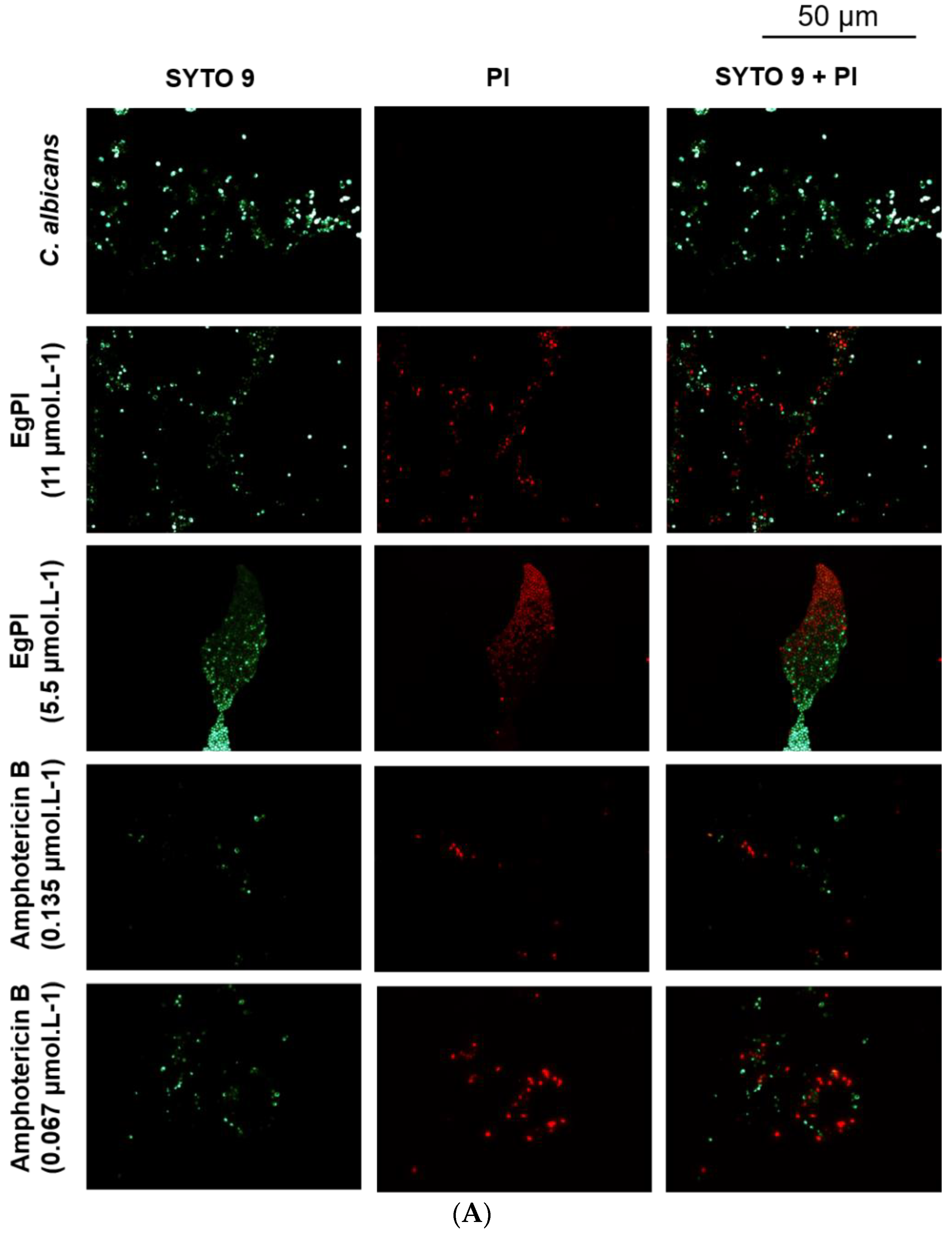
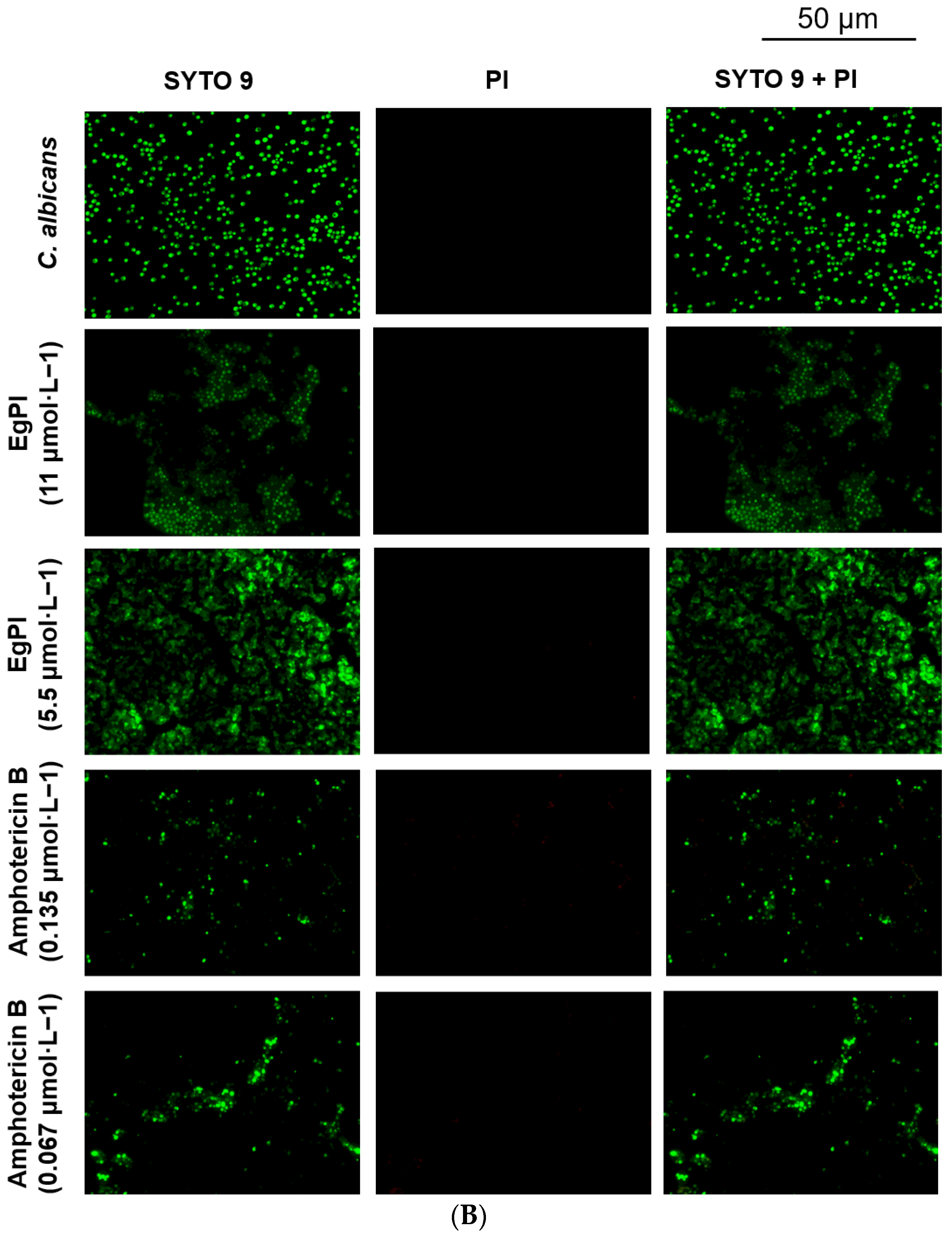
2.5. Antifungal Properties
3. Discussion
3.1. Inhibitory Activity and Ki Determination of EgPI
3.2. Inhibitory Activity and Ki Determination of EgPI
3.3. Amino-Terminal Identity
3.4. Analysis of the Secondary Structure and Stability of EgPI Using Circular Dichroism
3.5. Antifungal Activities
4. Materials and Methods
4.1. Materials
4.2. Purification of E. gummiferum Peptidase Inhibitor
4.3. Protein Quantification
4.4. Polyacrylamide Gel Electrophoresis
4.5. Inhibitory Activity
4.6. Determination of Kinetic Parameters and Dissociation Constant (Ki)
4.7. Amino-Terminal Sequencing
4.8. Circular Dichroism Spectral Analysis
4.9. Microorganisms
4.10. Antifungal Activity Assays
4.11. Evaluation of Biofilm Properties of EgPI
4.12. Analysis of Biofilm by Fluorescence Microscopy
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clemente, M.; Corigliano, M.G.; Pariani, S.A.; Sánchez-López, E.F.; Sander, V.A.; Ramos-Duarte, V.A. Plant Serine Protease Inhibitors: Biotechnology Application in Agriculture and Molecular Farming. Int. J. Mol. Sci. 2019, 20, 1345. [Google Scholar] [CrossRef] [PubMed]
- Santamaría, M.E.; Diaz-Mendoza, M.; Diaz, I.; Martinez, M. Plant Protein Peptidase Inhibitors: An Evolutionary Overview Based on Comparative Genomics. BMC Genom. 2014, 15, 812. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.K.; Atif, S.M.; Khan, R.H. Protein Proteinase Inhibitor Genes in Combat against Insects, Pests, and Pathogens: Natural and Engineered Phytoprotection. Arch. Biochem. Biophys. 2004, 431, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Hellinger, R.; Gruber, C.W. Peptide-Based Protease Inhibitors from Plants. Drug Discov. Today 2019, 24, 1877–1889. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS Database of Proteolytic Enzymes, Their Substrates and Inhibitors in 2017 and a Comparison with Peptidases in the PANTHER Database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Oliva, M.L.V.; Silva, M.C.C.; Sallai, R.C.; Brito, M.V.; Sampaio, M.U. A Novel Subclassification for Kunitz Proteinase Inhibitors from Leguminous Seeds. Biochimie 2010, 92, 1667–1673. [Google Scholar] [CrossRef]
- Bateman, K.S.; James, M.N.G. Plant Protein Proteinase Inhibitors: Structure and Mechanism of Inhibition. Curr. Protein Pept. Sci. 2011, 12, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Machado, S.W.; De Oliveira, C.F.R.; Bezerra, C.D.S.; Machado Freire, M.D.G.; Regina Kill, M.; Machado, O.L.T.; Marangoni, S.; Macedo, M.L.R. Purification of a Kunitz-Type Inhibitor from Acacia polyphylla DC Seeds: Characterization and Insecticidal Properties against Anagasta kuehniella Zeller (Lepidoptera: Pyralidae). J. Agric. Food Chem. 2013, 61, 2469–2478. [Google Scholar] [CrossRef]
- De Oliveira, C.F.R.; De Oliveira Flores, T.M.; Cardoso, M.H.; Oshiro, K.G.N.; Russi, R.; De França, A.F.J.; Dos Santos, E.A.; Franco, O.L.; De Oliveira, A.S.; Migliolo, L. Dual Insecticidal Effects of Adenanthera pavonina Kunitz-Type Inhibitor on Plodia Interpunctella Is Mediated by Digestive Enzymes Inhibition and Chitin-Binding Properties. Molecules 2019, 24, 4344. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; De Sá, C.M.; Freire, M.D.G.M.; Parra, J.R.P. A Kunitz-Type Inhibitor of Coleopteran Proteases, Isolated from Adenanthera pavonina L. Seeds and Its Effect on Callosobruchus maculatus. J. Agric. Food Chem. 2004, 52, 2533–2540. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; Freire, M.d.G.M.; Franco, O.L.; Migliolo, L.; de Oliveira, C.F.R. Practical and Theoretical Characterization of Inga laurina Kunitz Inhibitor on the Control of Homalinotus coriaceus. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2011, 158, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Dib, H.X.; de Oliveira, D.G.L.; de Oliveira, C.F.R.; Taveira, G.B.; de Oliveira Mello, E.; Verbisk, N.V.; Chang, M.R.; Corrêa Junior, D.; Gomes, V.M.; Macedo, M.L.R. Biochemical Characterization of a Kunitz Inhibitor from Inga edulis Seeds with Antifungal Activity against Candida spp. Arch. Microbiol. 2019, 201, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Araújo, N.M.S.; Dias, L.P.; Costa, H.P.S.; Sousa, D.O.B.; Vasconcelos, I.M.; de Morais, G.A.; Oliveira, J.T.A. ClTI, a Kunitz Trypsin Inhibitor Purified from Cassia leiandra Benth. Seeds, Exerts a Candidicidal Effect on Candida albicans by Inducing Oxidative Stress and Necrosis. Biochim. Biophys. Acta Biomembr. 2019, 1861, 183032. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.L.R.; Ribeiro, S.F.F.; Taveira, G.B.; Gomes, V.M.; de Barros, K.M.C.A.; Maria-Neto, S. Antimicrobial Activity of ILTI, a Kunitz-Type Trypsin Inhibitor from Inga laurina (SW.) Willd. Curr. Microbiol. 2016, 72, 538–544. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, C.F.R.; Oliveira, C.T.; Taveira, G.B.; de Oliveira Mello, E.; Gomes, V.M.; Macedo, M.L.R. Characterization of a Kunitz Trypsin Inhibitor from Enterolobium timbouva with Activity against Candida Species. Int. J. Biol. Macromol. 2018, 119, 645–653. [Google Scholar] [CrossRef] [PubMed]
- De Paula, C.A.A.; Coulson-Thomas, V.J.; Ferreira, J.G.; Maza, P.K.; Suzuki, E.; Nakahata, A.M.; Nader, H.B.; Sampaio, M.U.; Oliva, M.L.V. Enterolobium contortisiliquum Trypsin Inhibitor (EcTI), a Plant Proteinase Inhibitor, Decreases In Vitro Cell Adhesion and Invasion by Inhibition of Src Protein-Focal Adhesion Kinase (FAK) Signaling Pathways. J. Biol. Chem. 2012, 287, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, A.M.; Mayer, B.; Ries, C.; De Paula, C.A.A.; Karow, M.; Neth, P.; Sampaio, M.U.; Jochum, M.; Oliva, M.L.V. The Effects of a Plant Proteinase Inhibitor from Enterolobium contortisiliquum on Human Tumor Cell Lines. Biol. Chem. 2011, 392, 327–336. [Google Scholar] [CrossRef]
- Maqtari, M.A.; Saad, A.B.M. Screening and Purification of a Chymotrypsin Inhibitor from Entrolobium saman Seeds. Sultan Qaboos Univ. J. Sci. [SQUJS] 2010, 15, 19–29. [Google Scholar] [CrossRef]
- Morim, M.P.; Mesquita, A.L.; Bonadeu, F. Flora e Funga Do Brasil—Enterolobium gummiferum (Mart.) J.F.Macbr. Available online: https://floradobrasil.jbrj.gov.br/FB83156 (accessed on 5 June 2024).
- Batista, I.F.C.; Oliva, M.L.V.; Araujo, M.S.; Sampaio, M.U.; Richardson, M.; Fritz, H.; Sampaio, C.A.M. Primary Structure of a Kunitz-Type Trypsin Inhibitor from Enterolobium contortisiliquum Seeds. Phytochemistry 1996, 41, 1017–1022. [Google Scholar] [CrossRef]
- Wu, H.C.; Lin, J.Y. The Complete Amino Acid Sequence of a Kunitz Family Trypsin Inhibitor from Seeds of Acacia confusa. J. Biochem. 1993, 113, 258–263. [Google Scholar] [CrossRef]
- Monte Negreiros, A.N.; Carvalho, M.M.; Filho, J.X.; Blanco-Labra, A.; Shewry, P.R.; Richardson, M. The Complete Amino Acid Sequence of the Major Kunitz Trypsin Inhibitor from the Seeds of Prosopsis juliflora. Phytochemistry 1991, 30, 2829–2833. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, C.F.R.; Vasconcelos, I.M.; Aparicio, R.; Freire, M.D.G.M.H.; Baldasso, P.A.; Marangoni, S.; Macedo, M.L.R. Purification and Biochemical Properties of a Kunitz-Type Trypsin Inhibitor from Entada acaciifolia (Benth.) Seeds. Process Biochem. 2012, 47, 929–935. [Google Scholar] [CrossRef]
- Li, Q.; Huang, L.; Luo, Z.; Tamer, T.M. Stability of Trypsin Inhibitor Isolated from Potato Fruit Juice against PH and Heating Treatment and In Vitro Gastrointestinal Digestion. Food Chem. 2020, 328, 127152. [Google Scholar] [CrossRef] [PubMed]
- do Amaral, M.; Freitas, A.C.O.; Santos, A.S.; dos Santos, E.C.; Ferreira, M.M.; da Silva Gesteira, A.; Gramacho, K.P.; Marinho-Prado, J.S.; Pirovani, C.P. TcTI, a Kunitz-Type Trypsin Inhibitor from Cocoa Associated with Defense against Pathogens. Sci. Rep. 2022, 12, 698. [Google Scholar] [CrossRef] [PubMed]
- Gamal El-Din, M.; Eldahshan, O.; Singab, A.-N.; Ayoub, N.A. Genus Enterolobium: Traditional Uses, Chemistry and Biological Activities. Arch. Pharm. Sci. Ain Shams Univ. 2017, 1, 16–25. [Google Scholar] [CrossRef]
- McCue, J.T. Chapter 25 Theory and Use of Hydrophobic Interaction Chromatography in Protein Purification Applications. Methods Enzymol. 2009, 463, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Bendre, A.D.; Ramasamy, S.; Suresh, C.G. Analysis of Kunitz Inhibitors from Plants for Comprehensive Structural and Functional Insights. Int. J. Biol. Macromol. 2018, 113, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.L.R.; Durigan, R.A.; da Silva, D.S.; Marangoni, S.; Freire, M.d.G.M.; Parra, J.R.P. Adenanthera pavonina Trypsin Inhibitor Retard Growth of Anagasta kuehniella (Lepidoptera: Pyralidae). Arch. Insect Biochem. Physiol. 2010, 73, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.S.; Migliolo, L.; Aquino, R.O.; Ribeiro, J.K.C.; Macedo, L.L.P.; Andrade, L.B.S.; Bemquerer, M.P.; Santos, E.A.; Kiyota, S.; de Sales, M.P. Purification and Characterization of a Trypsin-Papain Inhibitor from Pithecelobium dumosum Seeds and Its in Vitro Effects towards Digestive Enzymes from Insect Pests. Plant Physiol. Biochem. 2007, 45, 858–865. [Google Scholar] [CrossRef]
- Zhou, D.; Lobo, Y.A.; Batista, I.F.C.; Marques-Porto, R.; Gustchina, A.; Oliva, M.L.V.; Wlodawer, A. Crystal Structures of a Plant Trypsin Inhibitor from Enterolobium contortisiliquum (EcTI) and of Its Complex with Bovine Trypsin. PLoS ONE 2013, 8, 62252. [Google Scholar] [CrossRef]
- Deshimaru, M.; Hanamoto, R.; Kusano, C.; Yoshimi, S.; Terada, S. Purification and Characterization of Proteinase Inhibitors from Wild Soja (Glycine soja) Seeds. Biosci. Biotechnol. Biochem. 2002, 66, 1897–1903. [Google Scholar] [CrossRef]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.H.; Goto, Y.; Réfrégiers, M.; Kardos, J. Accurate Secondary Structure Prediction and Fold Recognition for Circular Dichroism Spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.N.C.; Huang, L.; Noel, T.R.; Gunning, A.P.; Morris, V.J. Formation of Thermally Induced Aggregates of the Soya Globulin L-Conglycinin. Biochim. Biophys. Acta 2001, 1547, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Jiang, L.; Qi, B.; Zhou, L. Relationship between Secondary Structure and Surface Hydrophobicity of Soybean Protein Isolate Subjected to Heat Treatment. J. Chem. 2014, 2014, 475389. [Google Scholar] [CrossRef]
- Cruz, A.C.B.; Massena, F.S.; Migliolo, L.; Macedo, L.L.P.; Monteiro, N.K.V.; Oliveira, A.S.; Macedo, F.P.; Uchoa, A.F.; Grossi de Sá, M.F.; Vasconcelos, I.M.; et al. Bioinsecticidal Activity of a Novel Kunitz Trypsin Inhibitor from Catanduva (Piptadenia moniliformis) Seeds. Plant Physiol. Biochem. 2013, 70, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.L.R.; Garcia, V.A.; Freire, M.d.G.M.; Richardson, M. Characterization of a Kunitz Trypsin Inhibitor with a Single Disulfide Bridge from Seeds of Inga laurina (SW.) Willd. Phytochemistry 2007, 68, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Fang, Z.; Dicke, M.; van Loon, J.J.A.; Jongsma, M.A. The Diamondback Moth, Plutella xylostella, Specifically Inactivates Mustard Trypsin Inhibitor 2 (MTI2) to Overcome Host Plant Defence. Insect Biochem. Mol. Biol. 2009, 39, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Möhrlen, F.; Schnetter, W. Characterization of the Proteolytic Enzymes in the Midgut of the European Cockchafer, Melolontha melolontha (Coleoptera: Scarabaeidae). Insect Biochem. Mol. Biol. 2001, 32, 803–814. [Google Scholar] [CrossRef]
- Machado, S.W.; de Oliveira, C.F.R.; Zério, N.G.; Parra, J.R.P.; Macedo, M.L.R. Inga laurina Trypsin Inhibitor (ILTI) Obstructs Spodoptera frugiperda Trypsins Expressed during Adaptive Mechanisms against Plant Protease Inhibitors. Arch. Insect Biochem. Physiol. 2017, 95, e21393. [Google Scholar] [CrossRef]
- Ramos, V.D.S.; Cabrera, O.G.; Camargo, E.L.O.; Ambrósio, A.B.; Vidal, R.O.; Da Silva, D.S.; Guimarães, L.C.; Marangoni, S.; Parra, J.R.P.; Pereira, G.A.G.; et al. Molecular Cloning and Insecticidal Effect of Inga laurina Trypsin Inhibitor on Diatraea saccharalis and Heliothis virescens. Comp. Biochem. Physiol.-C Toxicol. Pharmacol. 2012, 156, 148–158. [Google Scholar] [CrossRef]
- Shen, J.J.; Jemec, G.B.E.; Arendrup, M.C.; Saunte, D.M.L. Photodynamic Therapy Treatment of Superficial Fungal Infections: A Systematic Review. Photodiagnosis Photodyn. Ther. 2020, 31, 101774. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Gongora, D.; Geddes-Mcalister, J. From Naturally-Sourced Protease Inhibitors to New Treatments for Fungal Infections. J. Fungi 2021, 7, 1016. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, F.C.; Weber, S.S.; Silva, O.N.; Jacobowski, A.C.; Ramada, M.H.S.; Macedo, M.L.R.; Franco, O.L.; Parachin, N.S. Recombinant Inga laurina Trypsin Inhibitor (ILTI) Production in Komagataella phaffii Confirms Its Potential Anti-Biofilm Effect and Reveals an Anti-Tumoral Activity. Microorganisms 2018, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Alencar de Barros, K.M.; Sardi, J.d.C.O.; Maria-Neto, S.; Macedo, A.J.; Ramalho, S.R.; Lourenço de Oliveira, D.G.; Pontes, G.S.; Weber, S.S.; Ramalho de Oliveira, C.F.; Macedo, M.L.R. A New Kunitz Trypsin Inhibitor from Erythrina poeppigiana Exhibits Antimicrobial and Antibiofilm Properties against Bacteria. Biomed. Pharmacother. 2021, 144, 112198. [Google Scholar] [CrossRef] [PubMed]
- MM, B. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- DelMar, E.G.; Largman, C.; Brodrick, J.W.; Geokas, M.C. A Sensitive New Substrate for Chymotrypsin. Anal. Biochem. 1979, 99, 316–320. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A Novel Method for Fast and Accurate Multiple Sequence Alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef]
- CLSI M07-A9; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Approved Standard—Ninth Edition; CLSI: Wayne, PA, USA, 2012.
- Filloux, A.; Ramos, J.-L. Pseudomonas Methods and Protocols; Humana: New York, NY, USA, 2014. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Steps | Total Protein (mg) | Total Activity (TIU) × 105 | Specific Activity (TIU·mg) × 104 | Yield (%) | Purification (Fold) |
|---|---|---|---|---|---|
| Crude extract (CE) | 100.00 | 12.62 | 1.26 | 100.00 | 1.00 |
| Sephadex G-100 | 21.42 | 10.27 | 4.80 | 81.38 | 3.80 |
| Phenyl HP | 5.41 | 7.13 | 13.17 | 56.46 | 10.44 |
| Inhibitors | Amino-Terminal Sequence | Identity (%) |
|---|---|---|
| EgPI | SELLDSDGDILDAGGAYYALPAVVS | |
| EcTI | KELLDSDGDILRNGGTYYILPALRG | 68 |
| AcTI | KELLDADGDILRNGGAYYILPALRG | 64 |
| PjTI | QELLDVDGEILRNGGSYYILPAFRG | 60 |
| Similarity | .**** **:** **:** ***. . |
| Conditions of Incubation | Secondary Structure Content (%) | |||
|---|---|---|---|---|
| α-Helix | β-Sheets | Random Coils | Loops | |
| EgPI | 2.9 | 21.6 | 45.7 | 29.8 |
| EgPI + DTT (1 mmol·L−1, 30 °C, 30 min) | 7.6 | 8.3 | 49.5 | 34.6 |
| EgPI at 20 °C | 6.6 | 38.3 | 32.9 | 22.2 |
| EgPI at 30 °C | 6.6 | 38.3 | 32.9 | 22.2 |
| EgPI at 40 °C | 9.9 | 14.4 | 40.7 | 35.0 |
| EgPI at 50 °C | 9.0 | 16.4 | 40.9 | 33.7 |
| EgPI at 60 °C | 9.2 | 15.1 | 42.1 | 33.5 |
| EgPI at 70 °C | 25.6 | 8.4 | 33.7 | 32.3 |
| EgPI at 80 °C | 36.1 | 8.0 | 24.6 | 31.2 |
| EgPI at 90 °C | 38.0 | 5.4 | 31.9 | 24.7 |
| EgPI at pH 2.0 | 5.4 | 13.3 | 46.2 | 35.0 |
| EgPI at pH 4.0 | 10.5 | 10.9 | 39.7 | 39.0 |
| EgPI at pH 6.0 | 8.6 | 13.5 | 44.1 | 33.8 |
| EgPI at pH 8.0 | 5.2 | 17.1 | 44.6 | 33.2 |
| EgPI at pH 10.0 | 7.6 | 9.7 | 48.2 | 34.5 |
| Strains | Planktonic | Biofilm | ||
|---|---|---|---|---|
| MIC (µmol·L−1) | MFC (µmol·L−1) | MBIC (µmol·L−1) | MBEC (µmol·L−1) | |
| C. albicans ATCC 5314 | >25 | >25 | 1.125 | 5.5 |
| C. albicans ATCC 90028 | >25 | >25 | ---- | ---- |
| C. parapsilosis ATCC 22019 | >25 | >25 | ---- | ---- |
| C. guillermonds ATCC 6260 | >25 | >25 | ---- | ---- |
| C. tropicalis ATCC 750 | >25 | >25 | 1.125 | >11 |
| C. glabrata ATCC 90030 | >25 | >25 | ---- | ---- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, M.M.; de Oliveira, C.F.R.; Almeida, C.V.; Sobrinho, I.A.S.; Macedo, M.L.R. A Novel Kunitz Trypsin Inhibitor from Enterolobium gummiferum Seeds Exhibits Antibiofilm Properties against Pathogenic Yeasts. Molecules 2024, 29, 3777. https://doi.org/10.3390/molecules29163777
da Silva MM, de Oliveira CFR, Almeida CV, Sobrinho IAS, Macedo MLR. A Novel Kunitz Trypsin Inhibitor from Enterolobium gummiferum Seeds Exhibits Antibiofilm Properties against Pathogenic Yeasts. Molecules. 2024; 29(16):3777. https://doi.org/10.3390/molecules29163777
Chicago/Turabian Styleda Silva, Matheus M., Caio F. R. de Oliveira, Claudiane V. Almeida, Ismaell A. S. Sobrinho, and Maria L. R. Macedo. 2024. "A Novel Kunitz Trypsin Inhibitor from Enterolobium gummiferum Seeds Exhibits Antibiofilm Properties against Pathogenic Yeasts" Molecules 29, no. 16: 3777. https://doi.org/10.3390/molecules29163777