1. Introduction
Triterpene saponins represent a vast group of plant metabolites with a multitude of biological activities. They can be classified into numerous groups and subgroups based on their aglycone type and the structure of the sugar moiety. Given the structural diversity of saponins, they are the subject of intensive investigation with regard to their biological activities, which include anti-inflammatory, adaptogenic, antimicrobial, antidiabetic, antiviral, and hepatoprotective effects [
1,
2,
3,
4,
5]. One of the most promising areas of research in the field of triterpene saponin bioactivity is the investigation of their potential to induce cytotoxicity in cancer cell lines in vitro, as well as their anti-tumor effects [
2,
6]. Triterpene saponins are present in a wide range of plant families, including the Amaranthaceae family [
7]. These are primarily oleanolic acid-type monodesmosidic or bidesmosidic derivatives, which have received attention due to their anticancer activity in both in vitro and in vivo testing [
6,
7,
8].
In the pursuit of novel, sustainable sources of these bioactive saponins, our research has focused on an as yet uninvestigated species:
Chenopodium strictum Roth (syn. C
henopodium betaceum Andrz.) [
9].
Chenopodium strictum Roth is an annual plant belonging to the genus
Chenopodium, which is widely distributed in North America, Europe, and Asia. It is commonly known as late-flowering goosefoot or striped goosefoot. The only published reports on the phytochemical composition of this plant species demonstrated the presence of flavonoids and proteins [
10,
11,
12]. However, no previous data refer to saponins.
This study reports the first isolation of oleanolic acid (OA)-type saponins from
Ch. strictum: calenduloside E (CE) and chikusetsusaponin IVa (ChIVa). Both compounds possess a glucuronic acid moiety at the C-3 position of the sapogenin. However, ChIVa differs from CE in that it contains an additional ester-linked glucose residue via the C-28 carboxyl group of OA. It may be significant to examine the impact of these compounds on cancer cells given that some authors have proposed that they may serve as metabolites of the more complex OA saponins and contribute to the observed effects in vivo [
8]. Moreover, the distinctions between these two saponins are noteworthy in the context of the structure–activity relationship (SAR).
Even though both compounds have previously been shown to exert cytotoxic effects in several cancer cell lines (calenduloside E against HeLA, MCF-7, A549, Lovo, LN229, WiDr, B16F10, and HepG2 cells [
8,
13,
14,
15,
16], whereas chikusetsusaponin IVa against HL-60, HCT-116, A2780, A2780, HEY, SK -Hep-1, HT-29, PC-3, LNCaP, Du145, HEC1B, and A549 cells [
8,
14,
17,
18,
19,
20,
21]), only two comparative analyses on the anticancer potential of these saponins have been carried out so far. One study investigated the effect of the compounds on WiDr and MCF-7 cancer cells [
14], while another examined the inhibitory effect on melanoma growth in 16F10 tumor-bearing mice [
8]. It is noteworthy that the cytotoxic activity of isolated saponins has not yet been compared with other compounds with a related structure, such as oleanolic acid, or saponins with an analogous structure and documented cytotoxic activity, such as momordin Ic (MIc).
Momordin Ic is the predominant saponin present in the seeds of
Bassia scoparia (
Kochiae fructus), a plant belonging to the Amaranthaceae family [
22]. MIc differs from calenduloside E solely in the presence of an additional xylose residue situated at the C-3′ position of the GlcA. The activity of this compound was primarily evaluated in models of prostate and gastrointestinal cancer. A number of published studies indicate that MIc may possess anticancer properties and exerts its effects through a range of mechanisms, including the PI3K/Akt-mediated pathways or MAPK-dependent PPARγ activation [
23,
24,
25]. Recent studies have demonstrated that MIc functions as an inhibitor of an SUMO protease isoform, SENP1. The overexpression of SENP1 has been observed in certain types of cancer, including prostate, lung, blood, and colon cancers [
26].
A substantial body of evidence suggests that the structural characteristics of a compound can influence its cytotoxic potency against cancer cells [
6,
27,
28]. Nevertheless, the research on OA-type sapogenins containing GlcA in the sugar moiety remains scarce. Furthermore, the impact of the structural characteristics on the activity of these compounds towards normal cells is rarely explored. The primary objective of this study, in addition to the isolation and structural elucidation of saponins from
Ch. strictum Roth, was to examine their cytotoxicity and selectivity in a wide range of human cell lines (cancerous and normal). Furthermore, the study sought to examine the relationship between the cytotoxic activity and compound structure. In addition to the isolated saponins, namely calenduloside E and chikusetsusaponin IVa, momordin Ic was also subjected to analysis, as was oleanolic acid, which represents the aglycone of these saponins. The study therefore encompasses a series of analogues with the potential to exert cytotoxic effects. To the best of our knowledge, comparative studies on the cytotoxic effects and selectivity of these compounds have not been conducted previously.
3. Materials and Methods
3.1. Chemicals and Reagents
Methanol, chloroform, ethyl acetate, butanol, 2-propanol, and acetone were obtained from CHEMPUR (Gliwice, Poland). Hydrochloric acid was obtained from Avantor Performance Materials Poland S.A. (Gliwice, Poland). All reagents were of analytical grade. Momordin Ic (phyproof® Reference Substance, purity ≥98.0% (HPLC), oleanolic acid (≥97%), D-glucose, D-galactose, L-arabinose, D-xylose, glucuronic acid, HPLC grade formic acid, lactic acid, and monosodium phosphate were purchased from Sigma-Aldrich (St. Louis, MO, USA). HPLC-grade methanol was purchased from Merck (Darmstadt, Germany). Water was prepared using a Milli-Q system (Millipore Corp., Bedford, MA, USA).
3.2. General Experimental Procedures
Medium-pressure liquid chromatography (MPLC) was performed on a Sepacore apparatus equipped with a C-615 Pump Manager (BÜCHI Labortechnik AG, Flawil, Switzerland). MPLC was carried out on NP: silica gel 230–400 mesh (Sigma-Aldrich, Darmstadt, Germany) and RP: reverse phase silica gel (LiChroprep, RP-18 (40–63 μm), Merck, Darmstadt, Germany).
Preparative high-pressure liquid chromatography (HPLC) was performed on a KNAUER Azura system (Knauer GmbH, Berlin, Germany): P 4.1S pump and a UV/VIS K-2600 detector (detection at 210 nm). Chromatographic separations were carried out using the Gemini® 5 µm NX-C18 110 Å LC column 250 × 10 mm (Phenomenex, Torrance, CA, USA) equipped with SecurityGuard SemiPrep cartridges Gemini-NX C18 10 × 10 mm precolumn.
Open-column chromatography (CC) was carried out on NP: silica gel (230–400 mesh; Sigma-Aldrich, Germany) using a glass column (350 × 25 mm).
Thin-layer chromatography (TLC) was performed on NP: silica gel 60 plates (Merck, Germany) and RP-18 F254S silica gel 60 plates (Supelco, Darmstadt, Germany). Detection: 25% solution of H2SO4 in methanol was used as spraying reagent, and visualization occurred after heating the plate at 120 °C for 4 min on a TLC plate heater (CAMAG, Muttenz, Switzerland).
LC–MS analysis was performed on UPLC/MS Waters ACQUITY TQD (Waters Corporation, Milford, MA, USA) apparatus operated in the negative and positive electrospray ionization modes. The Acquity UPLC bridged ethyl hybrid -BEH C18 column (100 × 2.1 mm, and particle size of 1.7 μm), equipped with Acquity UPLC BEH C18 VanGuard precolumn (2.1 × 5 mm, and particle size of 1.7 μm) was used for chromatographic separations under conditions described previously [
55]. MS detection settings of Waters TQD mass spectrometer were also described previously [
55].
NMR spectra (1D (1H NMR—500 MHz and DEPTQ—125 MHz) and 2D (HSQC, F2-coupled HSQC, H2BC, HMBC, COSY, TOCSY, and T-ROESY) were recorded in the mixture of pyridine-d5/D2O (250/10) with 0.2% TFA on a BrukerAvance III HD Ascend-500 spectrometer (Bruker BioSpin, Rheinstetten, Germany), equipped with 5 mm 1H{109Ag-31P} broad-band inverse (BBI) probe; coupling constants are reported in Hz. Chemical shifts (δ) are provided in ppm. Spectra were analyzed using ACD/Labs 1D NMR Processor 12.0 (Academic Edition)(Advanced Chemistry Development, Inc, Toronto, Ontario, Canada) and CARA 1.9.1.4. - Computer Aided Resonance Assignment (CARA Definition Team (CDT), free software licensed under the GNU General Public Licence (GPL))
The cytotoxicity study was conducted using a Microplate Reader (BioTek Instruments Inc., Winooski, VT, USA) with Gen 5 software.
3.3. Plant Material
Chenopodium strictum Roth was collected from natural habitat in July 2019, near Cracow, Poland (50°00′42.7″ N 19°59′42.6″ E). Species identity was confirmed by a botanist from the Department of Pharmacognosy UJCM, Cracow, Poland. The voucher specimen (No. Ch_str/2019_B) has been deposited at the Department of Pharmacognosy, Pharmaceutical Faculty, Medical College, Jagiellonian University, Cracow, Poland.
Plant material was divided into parts (roots, leaves, stems, and fruits) and dried under steady, controlled conditions (at 23 °C in air-conditioned room, in the dark) to a constant weight. The plant material was cut and ground to a fine powder using a mechanical laboratory mill (BOSCHMKM6003, BSHGmbH, Munich, Germany), and then kept in airtight containers.
3.4. Extraction and Isolation
The samples of powdered plant material (roots—2 g, leaves—2 g, fruits—2 g, and stems—2 g) were placed in a round bottom flasks and extracted sequentially with chloroform (plant material/solvent ratio—DSR 1:10, 2 times for 1 h) and methanol (DSR 1:10, 2 times for 2 h) by heat reflux extraction (under cooler with the use of a heating mantle). The combined MeOH extracts were concentrated under reduced pressure on a rotary evaporator to yield residues from roots (215 mg), leaves (427 mg), stems (317 mg), and fruits (147 mg). The dried crude extracts were subjected to thin-layer chromatography (TLC) using the conditions developed and described previously: silica gel, CHCl
3-CH
3OH-H
2O (20:12:2
v/
v), 25% methanolic H
2SO
4 + heating (120 °C/4 min) [
32]. Preliminary TLC investigation showed a presence of saponins in the roots of
Ch. strictum (
Figure S1). To isolate the saponins, extract from a larger sample of
Ch. strictum roots (250 g) was prepared. Extraction was performed using the conditions described above for a small sample of the plant material. The methanol extract was concentrated under reduced pressure using a rotary evaporator, and 22.7 g of a brown viscous residue was obtained, which was suspended in water and extracted twice with n-BuOH (100 mL). The n-butanol extract was evaporated to dryness under reduced pressure on a rotary evaporator (temp. 50–60 °C) to yield 15.5 g. Portions of the n-BuOH extract (2.0 g) were fractionated by column chromatography (CC, column 25 × 350 mm; silica gel 230–400 mesh) using the isocratic solvent system (CHCl
3-CH
3OH-H
2O (20:12:2
v/
v)). Fractions were collected and combined based on TLC examination (silica gel 60 plates, CHCl
3-CH
3OH-H
2O (20:12:2
v/
v).
CC chromatography yielded 14 pooled fractions (Fr/1-Fr/14), of which fractions F/4 and F/8 were further subjected to medium-pressure liquid chromatography (MPLC) on silica gel (230–400 mesh). The following conditions were used to fractionate Fr/4: MPLC, column 12 × 150 mm, flow rate 3 mL/min); isocratic solvent system: CHCl
3-CH
3OH-H
2O (23:12:2
v/
v). Fractions were combined based on TLC analysis (silica gel, CHCl
3-CH
3OH-H
2O (23:12:2
v/
v), 25% methanolic H
2SO
4 + heating, to yield 6 major fractions (Fr4/A-Fr4/F). The fraction Fr4/C rich in compound
1 (R
f = 0.51) was further separated by RP-MPLC chromatography (using solvent system: CH
3OH-H
2O (7:1.5
v/
v) in conditions previously described [
32] to yield seven fractions (Fr4C/1-Fr4C/7), of which fraction Fr4C/4 constituted compound
1 (21 mg), while fractions Fr4C/5 and Fr4C/6 were combined (31 mg) and further purified using a preparative RP-HPLC technique. The following chromatographic conditions were used: Phenomenex Gemini
® 5 µm NX-C18 110 Å LC column 250 × 10 mm (Phenomenex, Torrance, CA, USA); solvent system: CH
3OH-H
2O-HCOOH (8:2:0.01
v/
v/
v); flow rate: 2 mL/min; UV detection at 210 nm), which led to isolation of 10 mg of compound
1. Fractions containing compound
1, obtained by both MPLC and preparative HPLC, were controlled using UPLC-ESI-MS method under conditions described previously [
55] and pooled based on similarity and purity. As a result, the separation process yielded 36 mg of compound
1.
Fraction Fr/8 was chromatographed by MPLC (silica gel 60, column 12 × 150 mm, flow rate 2.5 mL/min) using the following solvent system: CHCl3-CH3OH-H2O (30:25:5 v/v) to yield 7 fractions (Fr8/A-Fr8/G).
The Fr8/C and Fr8/D were combined based on TLC similarity (silica gel, CHCl
3-CH
3OH-H
2O (30:25:5
v/
v); 25% methanolic H
2SO
4 + heating (120 °C/4 min) and further purified by RP-MPLC (using solvent system: CH
3OH-H
2O (7:3) in conditions previously described [
32]. The separation process using following conditions yielded 24 mg of compound
2.
3.5. Structure Elucidation
NMR spectra were recorded on a BrukerAvance III HD Ascend-500 spectrometer (Bruker BioSpin, Rheinstetten, Germany) (see
Section 3.2, General Experimental Procedures). Spectra were analyzed using ACD/Labs 1D NMR Processor 12.0 (Academic Edition) and CARA 1.9.1.4. (Computer Aided Resonance Assignment).
LC–MS analysis was performed on UPLC/MS Waters ACQUITY TQD (Waters Corporation, Milford, MA, USA) apparatus (see
Section 3.2, General Experimental Procedures).
Acid hydrolysis of isolated saponins (compound
1 and
2) was performed under the conditions described previously [
56]. The following sugar standards were used for analysis: D-galactose R
f = 0.26, D-glucose R
f = 0.38; L-arabinose R
f = 0.45; D-xylose R
f = 0.6, glucuronic acid lacton R
f = 0.80.
3.5.1. Compound 1 (CE): Oleanolic Acid-3-O-β-d-Glucuronopyranoside (Caleduloside E)
White powder: ESI MS (negative and positive ion modes)—data previously described [
32]. Acidic hydrolysis: sugar spot R
f = 0.81. For
1H and
13C NMR spectroscopic data, see
Table S1 (Supplementary Materials).
3.5.2. Compound 2 (ChIVa): 3-O-β-d-Glucuronopyranosyl Oleanolic Acid 28-O-β-d-Glucopyranosyl Ester (Chikusetsusaponin IVa)
White powder: ESI MS (negative and positive ion modes)—data previously described [
32]. Acidic hydrolysis: sugar spots: R
f = 0.38, R
f = 0.80. For
1H and
13C NMR spectroscopic data, see
Table S1 (Supplementary Materials).
3.6. Cell Culture
Cell lines used in the study were grouped as follows: skin panel (melanomas: HTB140, derived from metastatic site: lymph node, ATCC Hs 294 T; malignant A375, ATCC CRL-1619; stage I primary WM793, RRID:CVCL 8787; skin keratinocytes HaCaT); prostate panel (prostate carcinomas: Du145, derived from metastatic site: brain, ATCC HTB-81; grade IV PC3, derived from metastatic site: bone, ATCC CRL-1435, LNCaP, derived from the metastatic site: lymph node, ATCC CRL-1740; prostate epithelial, PNT2, ECACC 95012613); gastrointestinal panel (colorectal adenocarcinomas: Caco2, ATCC HTB-37, HT29, ATCC HTB-38, DLD-1, ATCC CCL-221, HCT116, ATCC CCL-247, and colon epithelial cells CCD 841 CoN, ATCC CRL-1790; hepatocellular carcinoma HepG2, ATCC HB-8065); thyroid panel (follicular thyroid carcinoma FTC133, ECACC 9406090, undifferentiated thyroid carcinoma 8505C, ECACC 94090184, papillary thyroid carcinoma TPC-1, SCC147, normal thyroid follicular epithelial Nthy-ori 3–1, ECACC 90011609); and breast panel (ER-positive breast adenocarcinoma MCF7, ATCC HTB-22; ER-negative breast adenocarcinoma MDA-MB-231, ATCC HTB-26; breast epithelial MCF10A, ATCC CRL-10317). Cells were grown under standard conditions (37 °C, 5% CO
2, relative humidity), and culture media (DMEM/F12 for PNT2, WM 793, HT29, PC3, FTC133, 8505 C, MDA-MB-231, TPC-1; DMEM Low Glucose for DU145; DMEM High Glucose for HTB140, A375, HaCaT; MEM with NEAA for Caco2 and MCF7; RPMI1640 with sodium pyruvate for LNCaP; DMEM/F12 with 20 ng/mL epidermal growth factor (EGF), 10 μg/mL insulin, 0.5 μg/mL hydrocortisone, 100 ng/mL cholera toxin) for MCF10A) supplemented with 10% fetal bovine serum (FBS) or 5% donor horse serum for MCF10A, and 1% antibiotics solution (10,000 U penicillin and 10 mg streptomycin/mL) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics solution (10,000 U penicillin and 10 mg streptomycin/mL), as previously described [
57,
58]. All cell lines, culture media, and supplements were from Sigma-Aldrich (Germany).
3.7. Cell Viability Assay
The cells were seeded in 96-well plates (1.5 × 10
4 cells/well). After 24 h, fresh medium was added, containing the tested compounds (0.5–100 μg/mL) and incubated for 24 or 48 h. Cell viability was determined by MTT assay, according to the manufacturer’s instructions, as described previously [
58]. Each experiment was performed in triplicate. The absorbance was measured at 570 nm using a Biotek Synergy microplate reader (BioTek Instruments Inc., Winooski, VT, USA). Cell viability was expressed as a percent of control (untreated cells). Doxorubicin was used as a reference drug.
3.8. Selectivity Index
The selectivity index (SI) was calculated from the following formula:
IC
50, half maximal inhibitory concentration, is a concentration required to kill 50% of the cells.
3.9. Chemometric Analysis
For each tested compound, a simple principal component analysis (PCA) model was constructed, in which cell lines were the objects and different concentrations of the test compound were the parameters. All parameters in all PCA models were mean-centered and scaled to unit variance. The parameters with large absolute values of their coordinates (>0.3) on the first two principal components in the PCA model were assumed to determine the axes of the new coordinate system in PCA to the greatest extent.
To express the strength of bivariate associations between such parameters, the cosines of the corresponding angles (i.e., correlation coefficients) were calculated. The “corresponding angle” means the angle determined by the two lines connecting the origin with coordinates of both parameters on the PCA loadings plot. The parameters were considered negatively correlated if their loadings within the PCA model showed the opposite signs; otherwise, they were considered positively correlated. Such association indicates only a similar effect of different concentrations of the tested compound on the cell lines used.
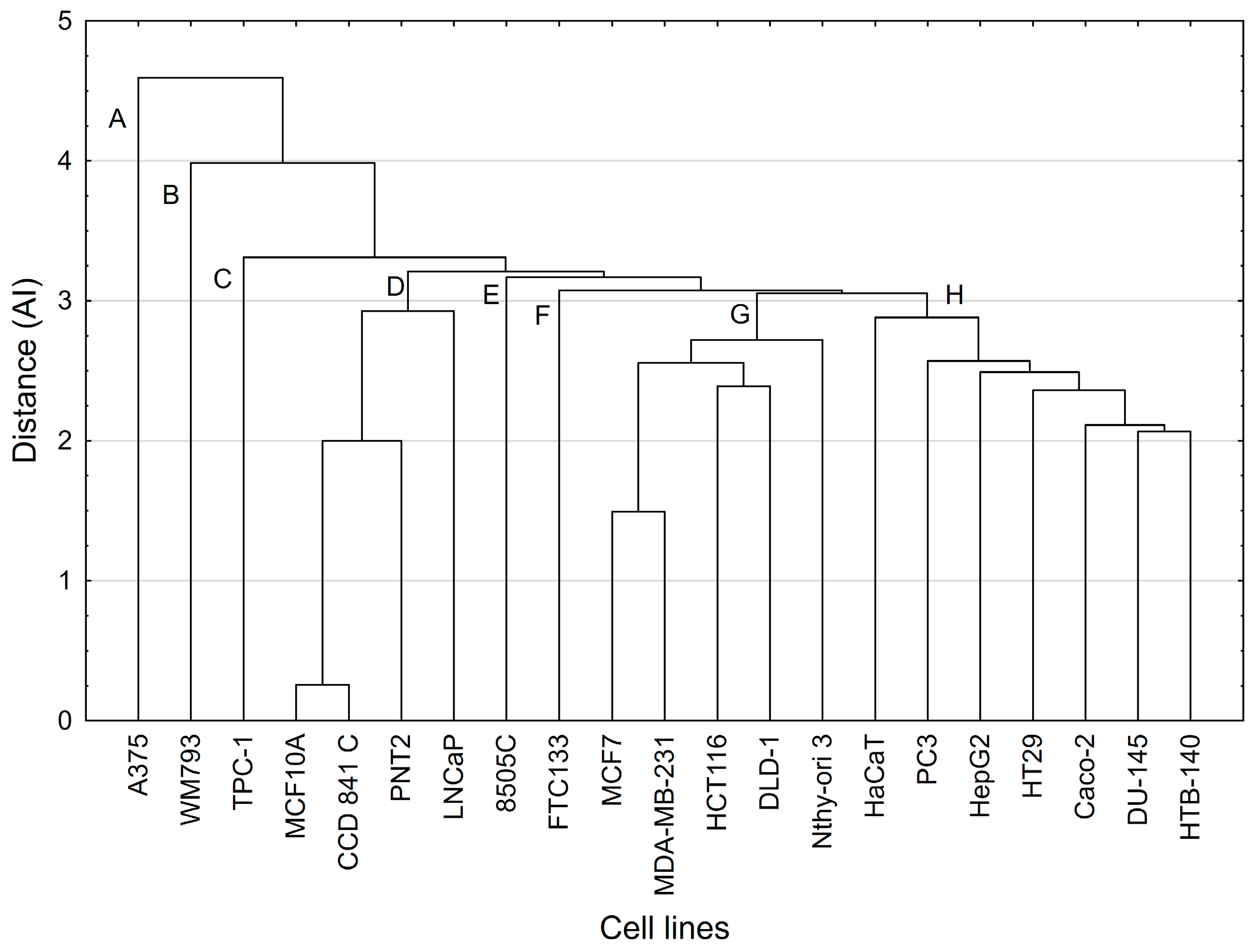
In the superior hierarchical PCA (hPCA) model, the first two principal components of each simple PCA model (previously constructed for the four tested compounds) were used as independent parameters.
A cluster analysis (CA) approach was used to test whether clusters of cell lines that appeared in the hPCA scores plot would be confirmed by another multivariate statistical method and, if so, what the possible reason was for this. Cluster analysis of results of cell lines (i.e., t-scores) obtained on the first three principal components of each of the above-mentioned four simple PCA models used single linkage procedure as a method of grouping and Euclidean distance as a function of the distance.
3.10. Data Analysis
The results were expressed as mean (±SD). Data were analyzed using Statistica v.13.3 (StatSoft, Tulsa, OK, USA). One-way analysis of variance (ANOVA) and the post hoc Tukey multiple comparison test were used. The probability level of p < 0.05 was considered statistically significant. Statistical analysis (PCA, CA) was conducted using packages STATISTICA v.13 (TIBCO Software Inc., Palo Alto, CA, USA) and SIMCA-P v.9 (Umetrics, Umeå, Sweden). The IC50 values in the cytotoxic assay were determined using Origin Pro 2020b (OriginLab Corporation, Northampton, MA, USA). Chemical structures of the compounds were illustrated with ChemDraw 19.0 software. Graphs were created with Excel 365 (Microsoft Office), and illustrations were designed using CorelDraw 2021.5.
4. Conclusions
Two known saponins, calenduloside E (CE) and chikusetsusaponin IVa (ChIVa), have been isolated from the roots of Chenopodium strictum Roth. Thus, our study revealed that Chenopodium strictum should be considered as a novel source of these oleanolic acid (OA)-type saponins. To the best of our knowledge, this is the first report on saponins in this plant species and constitutes a contribution to the general knowledge of the chemical composition of the Amaranthaceae family.
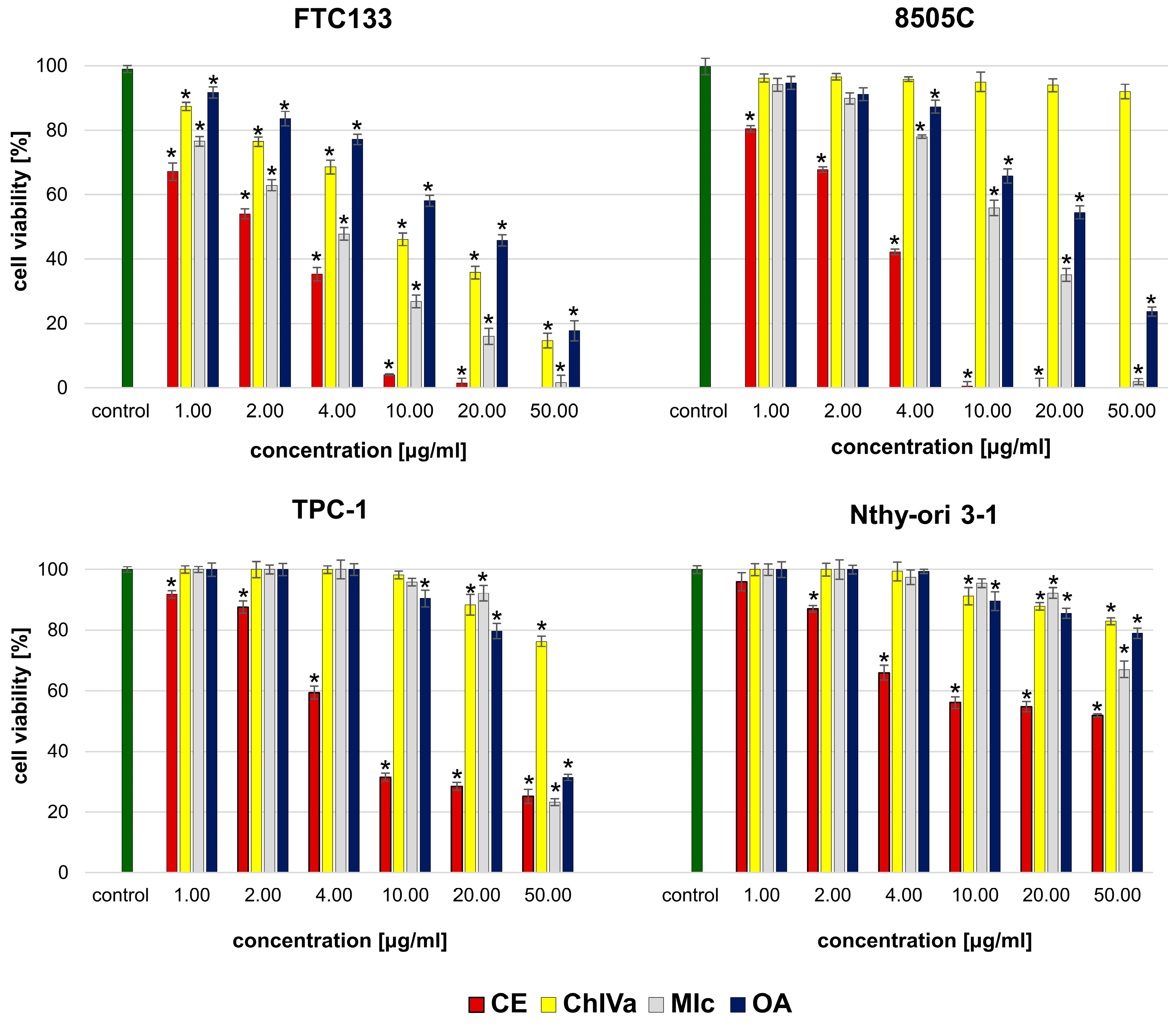
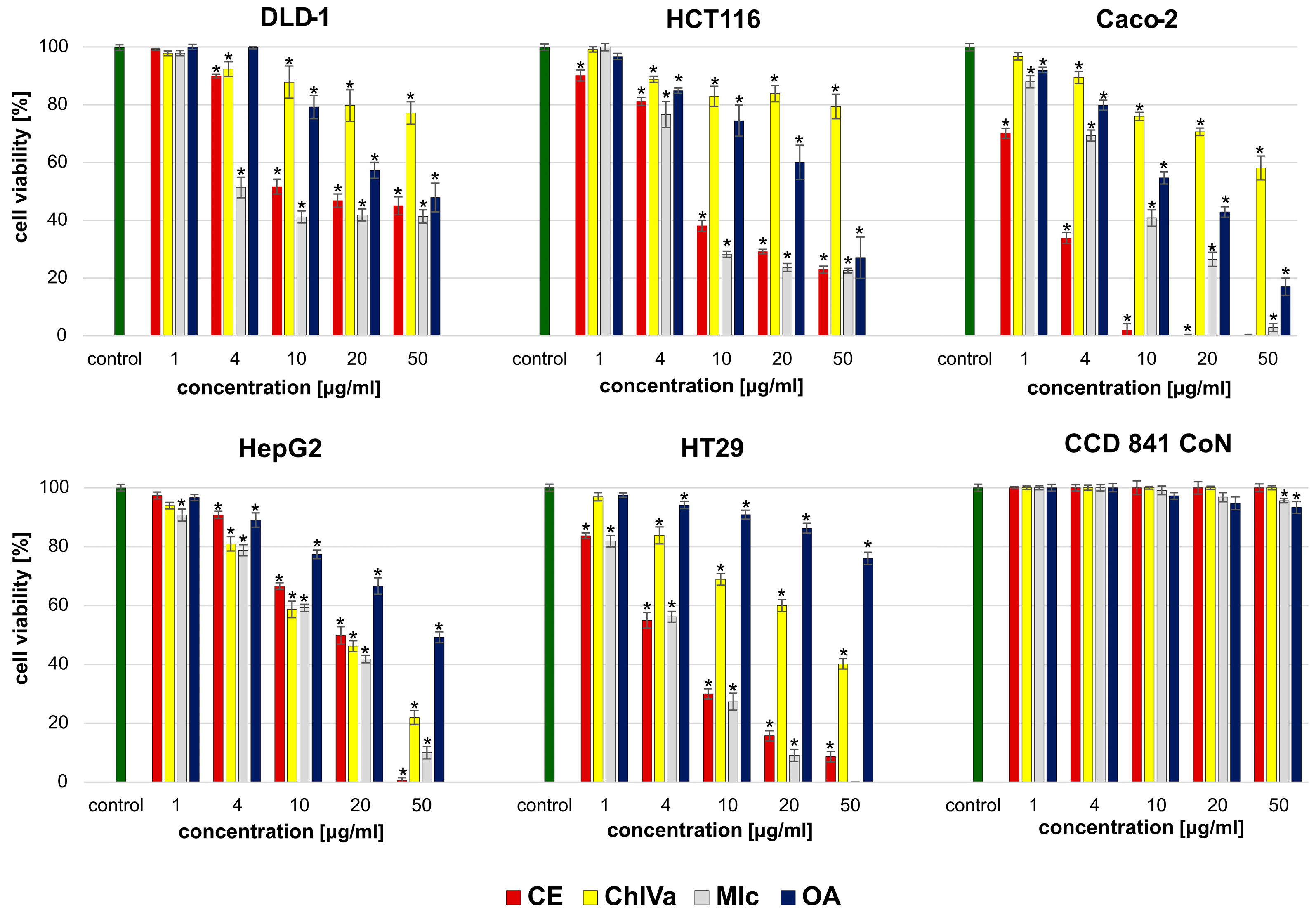
Both isolated saponins and the structurally similar momordin Ic (MIc), as well as oleanolic acid (OA), were found active in a cytotoxic assay, but the observed effect depended on the structure of the compound and the susceptibility of the human cancer cell line.
It appears that the key structural elements responsible for the strong cytotoxic activity against most cancer cell lines are the free carboxyl group (28COOH) in the structure of sapogenin (OA) and the presence of a sugar moiety in the molecule. Although the activity depends on the susceptibility of the specific cell line, monodesmosides containing a sugar moiety with glucuronic acid at the C3 position of OA have a stronger effect than bidesmoside or OA alone.
The results of our study enable suggesting that the presence of an additional xylose in the sugar chain affects the activity towards cancer cells, but this effect strictly depends on the specific cell line. The presence of an additional xylose residue at the C’3 position of the glucuronic acid in the sugar moiety suppressed the cytotoxicity against HTB-140, FTC133, and 8505C,Caco-2 cells while increasing the activity against DLD-1, HTC116, MDA-MB-231, and HepG2 cells compared to the effect of the compound without an extended sugar moiety. The obtained data indicate the importance of SAR screening against a large number of different cancer cells as incomplete conclusions regarding the overall activity may be drawn based on analyses of only one or a few cell lines.
The findings of this study revealed that OA-saponins bearing GlcA, especially CE and MIc, due to their cytotoxic potential and selectivity, are good candidates for further research on thyroid, breast, prostate, and gastrointestinal cancers. Nevertheless, further experiments in in vivo models are needed to confirm these in vitro observations. On the other hand, although the tested oleanolic acid derivatives, particularly CE, revealed high cytotoxicity towards melanoma cells, they did not act selectively. Therefore, while their potential against skin cancers is promising, our research indicates the need to explore ways to reduce their toxicity towards normal skin cells.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}