

Cultivated Winter-Type Lunaria annua L. Seed: Deciphering the Glucosinolate Profile Integrating HPLC, LC-MS and GC-MS Analyses, and Determination of Fatty Acid Composition

 ,
,  , , and
, , and 
Abstract

1. Introduction
2. Results and Discussion
2.1. Whole Seed Quality
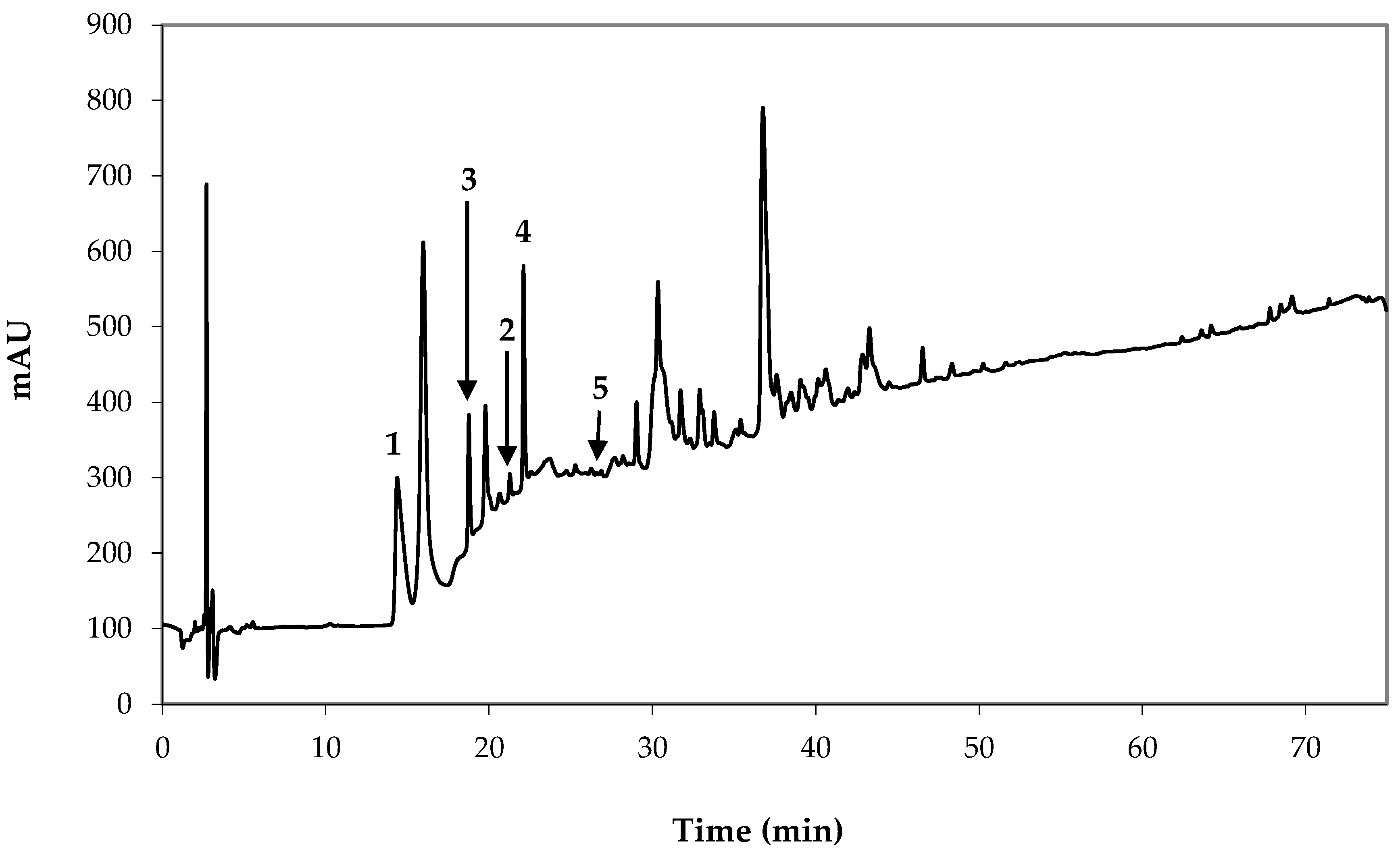
2.1.1. Intact Glucosinolate LC-MS Analysis
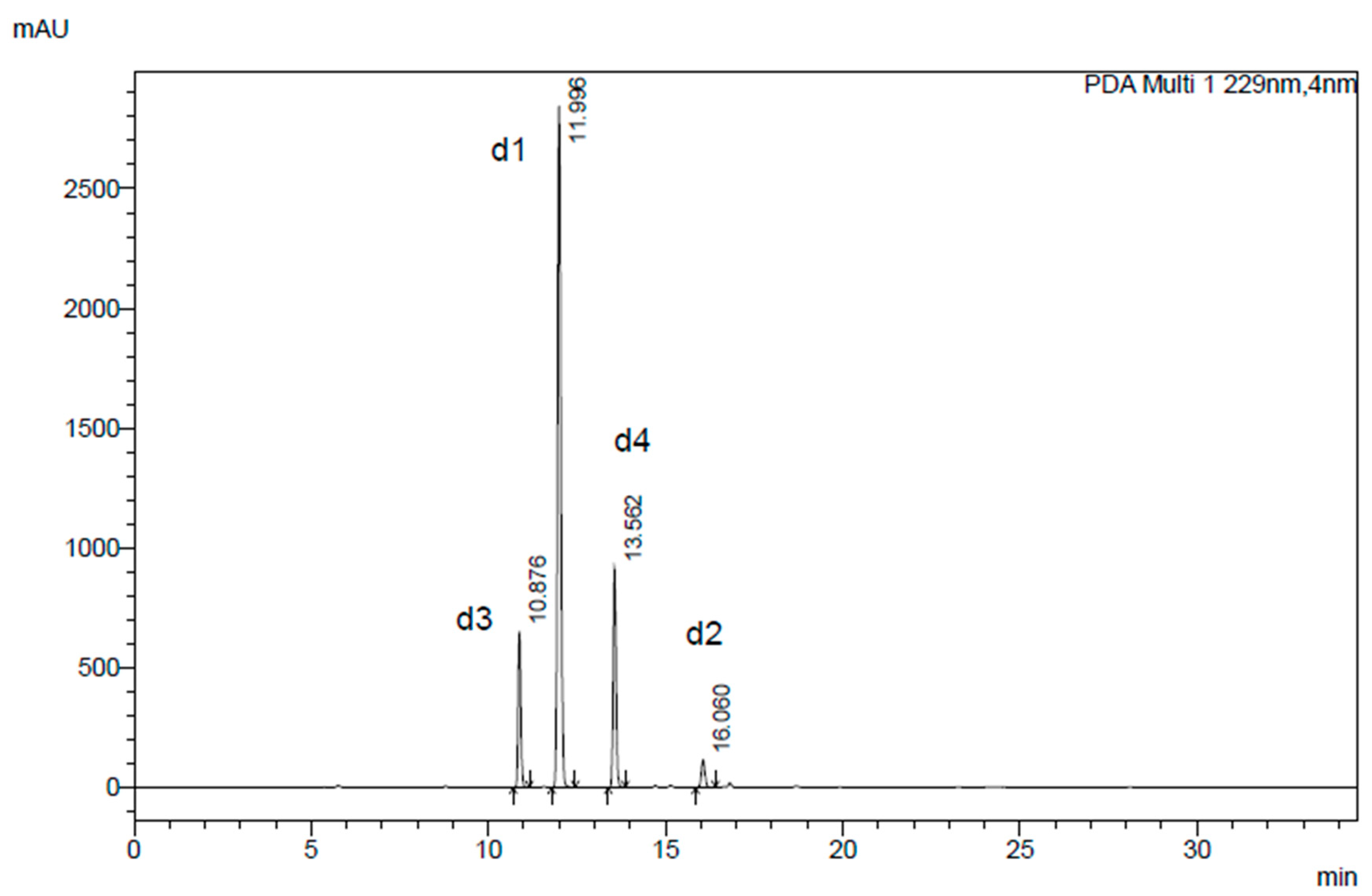
2.1.2. Desulfoglucosinolate HPLC Profiling and Quantitative Analysis by Laboratory A (CREA Research Centre for Vegetable and Ornamental Crops, Pescia, Italy)
2.1.3. Isothiocyanate GC-MS Analysis
2.1.4. Presscake Glucosinolate Content
2.1.5. Lunaria annua Presscake Prospects
2.1.6. Lunaria annua Seed Fatty Acid Composition
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Whole Seed Analyses
3.3.1. Moisture and Oil Content
3.3.2. LC-MS Analysis of Intact Glucosinolates
3.3.3. HPLC-PDA Analysis and Quantification of Desulfoglucosinolates by Laboratory A (CREA Research Centre for Vegetable and Ornamental Crops, Pescia, Italy)
3.3.4. Isothiocyanate Production through Myrosinase-Catalyzed Hydrolysis of Glucosinolates
3.3.5. GC-MS Analysis of Isothiocyanates
3.3.6. HPLC-DAD Analysis and Quantification of Desulfoglucosinolates in Whole Seed and Presscake by Laboratory B (Terres Inovia, Ardon, France)
3.3.7. Fatty Acid Extraction and Determination
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, F.U.; Khan, F.A.; Azki, I.; Mir, M.M. Effect of growing seasons, nitrogen and phosphorus on growth, flowering and spike production of lunaria (Lunaria biennis Linn.). J. Ornam. Hortic. 2005, 8, 290–292. [Google Scholar]
- Leins, P.; Fligge, K.; Erbar, C. Silique valves as sails in anemochory of Lunaria (Brassicaceae). Plant Biol. 2018, 20, 238–243. [Google Scholar] [CrossRef]
- Guo, X.; Mandáková, T.; Trachtová, K.; Özüdoğru, B.; Liu, J.; Lysak, M.A. Linked by ancestral bonds: Multiple whole-genome duplications and reticulate evolution in a Brassicaceae tribe. Mol. Biol. Evol. 2021, 38, 1695–1714. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, F.; Monti, A.; Berti, M.T. Challenges and opportunities for new industrial oilseed crops in EU-27: A review. Ind. Crops Prod. 2013, 50, 580–595. [Google Scholar] [CrossRef]
- Dodos, G.S.; Karonis, D.; Zannikos, F.; Lois, E. Renewable fuels and lubricants from Lunaria annua L. Ind. Crops Prod. 2015, 75, 43–50. [Google Scholar] [CrossRef]
- Wang, P.; Xiong, X.; Zhang, X.; Wu, G.; Liu, F. A review of erucic acid production in Brassicaceae oilseeds: Progress and prospects for the genetic engineering of high and low-erucic acid rapeseeds (Brassica napus). Front. Plant Sci. 2022, 13, 899076. [Google Scholar] [CrossRef]
- Liu, F.; Wang, P.; Xiong, X.; Zeng, X.; Zhang, X.; Wu, G. A review of nervonic acid production in plants: Prospects for the genetic engineering of high nervonic acid cultivars plants. Front. Plant Sci. 2021, 12, 626625. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sun, H.; Zhou, Q.; Yu, G.; Qin, D.; Ma, Q. Nervonic acid regulates the oxidative imbalance in experimental allergic encephalomyelitis. Food Sci. Technol. Res. 2021, 27, 269–280. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, e112100. [Google Scholar] [CrossRef]
- Galanakis, C.M. Recovery techniques, stability, and applications of glucosinolates. In Glucosinolates: Properties, Recovery, and Applications; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 251–280. [Google Scholar] [CrossRef]
- Mastebroek, H.D.; Marvin, H.J.P. Breeding prospects of Lunaria annua L. Ind. Crops Prod. 2000, 11, 139–143. [Google Scholar] [CrossRef]
- Walker, R.L.; Walker, K.C.; Booth, E.J. Adaptation potential of novel oilseed crop, Honesty (Lunaria annua), to the Scottish climate. Ind. Crops Prod. 2003, 18, 7–15. [Google Scholar] [CrossRef]
- Martin, R.J.; Porter, N.G.; Deo, B. Initial studies on seed oil composition of Calendula and Lunaria. Agronomy N.Z. 2005, 35, 129–137. Available online: https://www.agronomysociety.org.nz/files/2005_14._Seed_oil_composition_Calendula_Lunaria.pdf (accessed on 24 June 2024).
- Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Screening crucifer seeds as sources of specific intact glucosinolates using ion-pair high-performance liquid chromatography negative ion electrospray mass spectrometry. J. Agric. Food Chem. 2004, 52, 428–438. [Google Scholar] [CrossRef]
- Vaughn, S.; Palmquist, D.; Duval, S.; Berhow, M. Herbicidal activity of glucosinolate-containing seedmeals. Weed Sci. 2006, 54, 743–748. [Google Scholar] [CrossRef]
- Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Popović, M.; Guillot, X.; Burčul, F.; Rollin, P. Microwave-assisted versus conventional isolation of glucosinolate degradation products from Lunaria annua L. and their cytotoxic activity. Biomolecules 2020, 10, 215. [Google Scholar] [CrossRef]
- ISO 9167:2019; Rapeseed and Rapeseed Meals—Determination of Glucosinolates Content—Method Using High-Performance Liquid Chromatography. ISO: Geneva, Switzerland, 2019. Available online: https://www.iso.org/standard/72207.html (accessed on 24 June 2024).
- ISO 12966-4:2015; Animal and Vegetable Fats and Oils—Gas Chromatography of Fatty Acid Methyl esters—Part 4: Determination by Capillary Gas Chromatography. ISO: Geneva, Switzerland, 2015. Available online: https://www.iso.org/standard/63503.html (accessed on 24 June 2024).
- Montaut, S.; Grandbois, J.; Righetti, L.; Barillari, J.; Iori, R.; Rollin, P. Updated glucosinolate profile of Dithyrea wislizenii. J. Nat. Prod. 2009, 72, 889–893. [Google Scholar] [CrossRef] [PubMed]
- Montaut, S.; Bleeker, R.S.; Jacques, C. Phytochemical constituents of Cardamine diphylla. Can. J. Chem. 2010, 88, 50–55. [Google Scholar] [CrossRef]
- Blažević, I.; De Nicola, G.R.; Montaut, S.; Rollin, P. Glucosinolates in two endemic plants of the Aurinia genus and their chemotaxonomic significance. Nat. Prod. Commun. 2013, 8, 1463–1466. [Google Scholar] [CrossRef] [PubMed]
- Botting, C.H.; Davidson, N.E.; Griffiths, D.W.; Bennett, R.N.; Botting, N.P. Analysis of intact glucosinolates by MALDI-TOF mass spectrometry. J. Agric. Food Chem. 2002, 50, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D.W.; Bain, H.; Deighton, N.; Botting, N.P.; Robertson, A.A. Evaluation of liquid chromatography–atmospheric pressure chemical ionisation–mass spectrometry for the identification and quantification of desulphoglucosinolates. Phytochem. Anal. 2000, 11, 216–225. [Google Scholar] [CrossRef]
- Wathelet, J.-P.; Iori, R.; Leoni, O.; Rollin, P.; Quinsac, A.; Palmieri, S. Guidelines for glucosinolate analysis in green tissues used for biofumigation. Agroindustria 2004, 3, 257–266. [Google Scholar]
- Clarke, D. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310–325. [Google Scholar] [CrossRef]
- Kjær, A.; Ohashi, M.; Wilson, J.M.; Djerassi, C. Mass spectra of isothiocyanates. Acta Chem. Scand. 1963, 17, 2143–2154. [Google Scholar] [CrossRef]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Compendium of Food Additive Specifications. 69th Meeting June 2008. In FAO JECFA Monographs, 5th ed.; FAO: Rome, Italy, 2008; 141p, Available online: https://www.fao.org/publications/card/en/c/93ba3fb3-fe00-54f4-b9e6-91176f88e8ce (accessed on 24 June 2024).
- NIST National Institute of Standards and Technology Chemistry WebBook, Standard Reference Database Number 69, 2023. Available online: https://webbook.nist.gov/chemistry/ (accessed on 24 June 2024). [CrossRef]
- Han, N.; Ku, K.M.; Kim, J. Postharvest variation of major glucosinolate and their hydrolytic products in Brassicoraphanus ‘BB1’. Postharvest Biol. Technol. 2019, 154, 70–78. [Google Scholar] [CrossRef]
- Spencer, G.F.; Daxenbichler, M.E. Gas chromatography-mass spectrometry of nitriles, isothiocyanates and oxazolidinethiones derived from cruciferous glucosinolates. J. Sci. Food Agric. 1980, 31, 359–367. [Google Scholar] [CrossRef]
- Vaughn, S.; Berhow, M. Glucosinolate hydrolysis products from various plant sources: pH effects, isolation, and purification. Ind. Crops Prod. 2005, 21, 193–202. [Google Scholar] [CrossRef]
- Chiang, W.C.K.; Pusateri, D.J.; Leitz, R.E.A. Gas chromatography/mass spectrometry method for the determination of sulforaphane and sulforaphane nitrile in broccoli. J. Agric. Food Chem. 1998, 46, 1018–1021. [Google Scholar] [CrossRef]
- Cedrowski, J.; Dąbrowa, K.; Przybylski, P.; Krogul-Sobczak, A.; Litwinienko, G. Antioxidant activity of two edible isothiocyanates: Sulforaphane and erucin is due to their thermal decomposition to sulfenic acids and methylsulfinyl radicals. Food Chem. 2021, 353, 129213. [Google Scholar] [CrossRef]
- Borgonovo, G.; Zimbaldi, N.; Guarise, M.; De Nisi, P.; De Petrocellis, L.; Schiano Moriello, A.; Bassoli, A. Isothiocyanates and glucosinolates from Sisymbrium officinale (L.) Scop. (“the singers’ plant”): Isolation and in vitro assays on the somatosensory and pain receptor TRPA1 channel. Molecules 2019, 24, 949. [Google Scholar] [CrossRef]
- Borgonovo, G.; Zimbaldi, N.; Guarise, M.; Bedussi, F.; Winnig, M.; Vennegeerts, T.; Bassoli, A. Glucosinolates in Sisymbrium officinale (L.) Scop.: Comparative analysis in cultivated and wild plants and in vitro assays with T2Rs bitter taste receptors. Molecules 2019, 24, 4572. [Google Scholar] [CrossRef]
- Di Sotto, A.; Di Giacomo, S.; Vitalone, A.; Nicoletti, M.; Mazzanti, G. Antimutagenic thio compounds from Sisymbrium officinale. J. Nat. Prod. 2012, 75, 2062–2068. [Google Scholar] [CrossRef]
- De Nicola, G.R.; Rollin, P.; Mazzon, E.; Iori, R. Novel gram-scale production of enantiopure R-sulforaphane from Tuscan black kale seeds. Molecules 2014, 19, 6975–6986. [Google Scholar] [CrossRef] [PubMed]
- Mangla, B.; Javed, S.; Sultan, M.H.; Kumar, P.; Kohli, K.; Najmi, A.; Alhazmi, H.A.; Al Bratty, M.; Ahsan, W. Sulforaphane: A review of its therapeutic potentials, advances in its nanodelivery, recent patents, and clinical trials. Phytother. Res. 2021, 35, 5440–5458. [Google Scholar] [CrossRef] [PubMed]
- Kinae, N.; Masuda, H.; Shin, I.S.; Furugori, M.; Shimoi, K. Functional properties of wasabi and horseradish. BioFactors 2000, 13, 265–269. [Google Scholar] [CrossRef]
- Miklavčič Višnjevec, A.; Tamayo Tenorio, A.; Steenkjær Hastrup, A.C.; Hansen, N.M.L.; Peeters, K.; Schwarzkopf, M. Glucosinolates and isothiocyanates in processed rapeseed determined by HPLC-DAD-qTOF. Plants 2021, 10, 2548. [Google Scholar] [CrossRef]
- Hebert, M.; Mhemdi, H.; Vorobiev, E. Selective and eco-friendly recovery of glucosinolates from mustard seeds (Brassica juncea) using process optimization and innovative pretreatment (high voltage electrical discharges). Food Bioprod. Process. 2020, 124, 11–23. [Google Scholar] [CrossRef]
- Meza, S.; Zhou, Y.; Chastain, J.; Yang, Y.; Cheng, H.H.; Iassonova, D.; Rivest, J.; You, H. Eco-Efficient quantification of glucosinolates in Camelina seed, oil, and defatted meal: Optimization, development, and validation of a UPLC-DAD method. Antioxidants 2022, 11, 2441. [Google Scholar] [CrossRef] [PubMed]
- Drabińska, N.; Siger, A.; Jeleń, H.H. Unravelling the importance of seed roasting for oil quality by the non-targeted volatilomics and targeted metabolomics of cold-pressed false flax (Camelina sativa L.) oil and press cakes. Food Chem. 2024, 458, 140207. [Google Scholar] [CrossRef]
- Singh, B.K.; Bala, M.; Rai, P.K. Fatty acid composition and seed meal characteristics of Brassica and allied genera. Natl. Acad. Sci. Lett. 2014, 37, 219–226. [Google Scholar] [CrossRef]
- Lazzeri, L.; Malaguti, L.; Bagatta, M.; D’Avino, L.; Ugolini, L.; De Nicola, G.R.; Casadei, N.; Cinti, S.; Matteo, R.; Iori, R. Characterization of the main glucosinolate content and fatty acid composition in non-food Brassicaceae seeds. Acta Hortic. 2013, 1005, 331–338. [Google Scholar] [CrossRef]
- Todorović, Z.B.; Mitrović, P.M.; Zlatković, V.; Grahovac, N.L.; Banković-Ilić, I.B.; Troter, D.Z.; Marjanović-Jeromela, A.M.; Veljković, V.B. Optimization of oil recovery from oilseed rape by cold pressing using statistical modeling. Food Meas. 2024, 18, 474–488. [Google Scholar] [CrossRef]
- NF V03-909:2021. NF V03-909:2021; Graines Oléagineuses—Détermination de la Teneur en eau et en Matières Volatiles—Méthode simplifiée. Afnor Editions: La Plaine Saint-Denis, France, 2021; Available online: https://norminfo.afnor.org/norme/nf-v03-909/graines-oleagineuses-determination-de-la-teneur-en-eau-et-en-matieres-volatiles-methode-simplifiee/194342 (accessed on 24 June 2024).
- NF V03-908:2009; Graines Oléagineuses—Détermination de la Teneur en Huile—Méthode Alternative. Afnor Editions: La Plaine Saint-Denis, France, 2009. Available online: https://norminfo.afnor.org/consultation/nf-v03-908/graines-oleagineuses-determination-de-la-teneur-en-huile-methode-alternative/56924 (accessed on 24 June 2024).
- Zrybko, C.L.; Fukuda, E.K.; Rosen, R.T. Determination of glucosinolates in domestic and wild mustard by high-performance liquid chromatography with confirmation by electrospray mass spectrometry and photodiode-array detection. J. Chromatogr. A. 1997, 767, 43–52. [Google Scholar] [CrossRef]
- Montaut, S.; Zhang, W.D.; Nuzillard, J.M.; De Nicola, G.R.; Rollin, P. Glucosinolate Diversity in Bretschneidera sinensis of Chinese Origin. J. Nat. Prod. 2015, 78, 2001–2006. [Google Scholar] [CrossRef] [PubMed]
- ISO 12966-2:2017; Animal and Vegetable Fats and Oils—Gas Chromatography of Fatty Acid Methyl Esters—Part 2: Preparation of Methyl Esters of Fatty Acids. ISO: Geneva, Switzerland, 2017. Available online: https://www.iso.org/standard/72142.html (accessed on 24 June 2024).


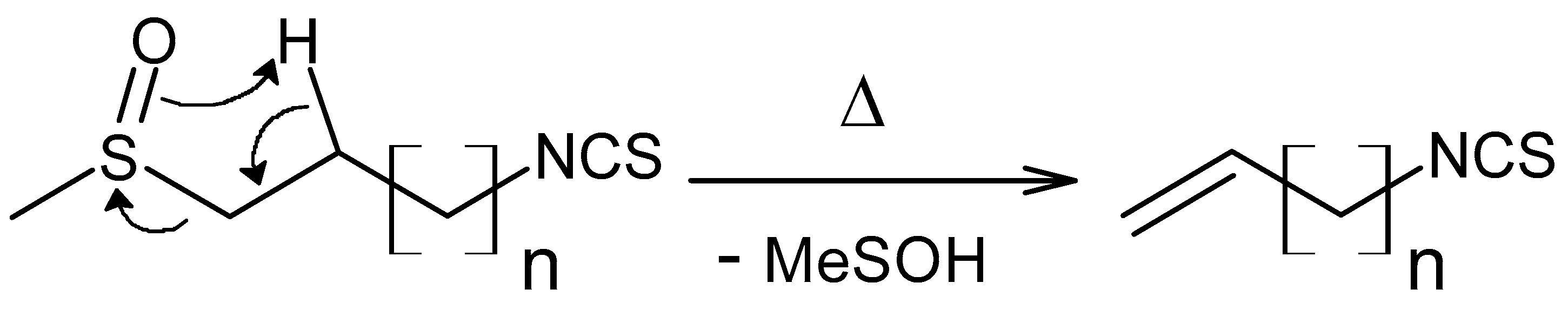

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucosinolate (GSL) | Whole Seed Content | ||||||
|---|---|---|---|---|---|---|---|
| # | Class | Semisystematic Name (Trivial Name) | [M − H] 1 m/z (%) | µmol g−1 | mg g−1 | % on Total GSLs | % (w/w) |
| 1 | Val | 1-Methylethyl GSL (glucoputranjivin) | 359.8 (100) | 44.58 ± 1.10 2,3 | 16.11 ± 0.40 | 64.75 | 1.61 |
| 2 | Ile | (1S)-1-Methylpropyl GSL (glucocochlearin) | 374.0 (100) | 1.76 ± 0.05 | 0.66 ± 0.02 | 2.56 | 0.07 |
| 3 | Met | (Rs)-5-(Methylsulfinyl)pentyl GSL (glucoalyssin) | 449.8 (100) | 8.87 ± 0.20 | 4.00 ± 0.09 | 12.88 | 0.40 |
| 4 | Met | (Rs)-6-(Methylsulfinyl)hexyl GSL (glucohesperin) | 464.0 (100) | 13.64 ± 0.74 | 6.35 ± 0.35 | 19.81 | 0.63 |
| 5 | Met | (2S)-2-Hydroxy-4-pentenyl GSL (gluconapoleiferin) | 402.0 (100) | - | - | - | - |
| Total | 68.85 ± 2.09 | 27.12 ± 0.86 | 100 | 2.71 | |||
| Isothiocyanates (ITCs) | ||||
|---|---|---|---|---|
| Peak # | Semisystematic Name (Trivial Name) | Precursor Compound | Injector Port T (°C) a | MS Spectral Data (m/z) b |
| 1 | 1-Methylethyl ITC (putranjivin) | 1-Methylethyl GSL (glucoputranjivin) | 180, 250 | 101, 86, 72, 60, 59, 58, 43, 42, 41, 39 Lit. c,d,e |
| 2 | (1S)-1-Methylpropyl ITC | (1S)-1-Methylpropyl GSL (glucocochlearin) | 180, 250 | 115, 86, 72, 57, 56, 41, 39 Lit. c,d,e |
| 3 | 4-Pentenyl ITC (brassicanapin) | (Rs)-5-(Methylsulfinyl)pentyl ITC (R-alyssin) | 250 | 127, 126, 112, 99, 85, 72, 70, 67, 60, 59, 58, 55, 53, 41, 39 Lit. c,d,f |
| 4 | 5-Hexenyl ITC | (Rs)-6-(Methylsulfinyl)hexyl ITC (R-hesperin) | 250 | 141, 140, 126, 113, 108, 97, 84,72, 67, 55, 54, 41, 39 Lit. d |
| 5 | (Rs)-5-(Methylsulfinyl)pentyl ITC (R-alyssin) | (Rs)-5-(Methylsulfinyl)pentyl GSL (glucoalyssin) | 180, 250 | 174, 128, 72, 69, 41, 39 Lit. g |
| 6 | (Rs)-6-(Methylsulfinyl)hexyl ITC (R-hesperin) b | (Rs)-6-(Methylsulfinyl)hexyl GSL (glucohesperin) | 180, 250 | 188, 142, 126, 72, 55, 41, 39 Lit. g,h |
| Humidity (%) | Oil (%) | Glucosinolate Content | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (µmol g−1) | % (w/w) | |||||||||||||||||
| d1 | d2 | d3 | d4 | d5 | d6 | d7 | Total | 1 | 2 | 3 | 4 | 5 | 6 | 7 | Total | |||
| Whole seed | 8.7 | 36.8 | 47.01 ± 1.51 | 1.90 ± 0.04 | 8.10 ± 0.10 | 9.01 ± 0.06 | n.d. | 0.20 ± 0.01 | n.d. | 66.22 ± 1.72 | 1.7 | 0.1 | 0.4 | 0.4 | n.d. | - | n.d | 2.6 |
| Cold pressure extraction | ||||||||||||||||||
| Presscake | 80.10 ± 1.47 | 3.01 ± 0.03 | 14.00 ± 0.19 | 14.71 ± 0.23 | 0.19 ± 0.04 | 0.31 ± 0.01 | 0.12 ± 0.01 | 112.44 ± 1.98 | 2.9 | 0.1 | 0.6 | 0.7 | - | - | - | 4.3 | ||
| Fatty Acid (FA) | Content (% on Total FAs) | |
|---|---|---|
| Semisystematic Name (Trivial Name) | ||
| Hexadecanoic acid (palmitic acid) | C16:0 | 1.3 ± 0.1 1,2 |
| Octadecanoic acid (stearic acid) | C18:0 | 0.1 ± 0.0 |
| Total saturated fatty acids (SFAs) | 1.4 ± 0.1 | |
| (Z)-Octadec-9-enoic acid (oleic acid) | C18:1 ω9 | 24.2 ± 1.1 |
| (Z)-Icos-11-enoic acid (gondoic acid) | C20:1 ω9 | 0.9 ± 0.4 |
| (Z)-Docos-13-enoic acid (erucic acid) | C22:1 ω9 | 44.6 ± 0.1 |
| (Z)-tetracos-15-enoic acid (nervonic acid) | C24:1 ω9 | 21.8 ± 0.8 |
| Total monounsaturated fatty acids (MUFAs) | 91.5 ± 2.4 | |
| (9Z,12Z)-octadeca-9,12-dienoic acid (linoleic acid) | C18:2 ω6 | 6.4 ± 0.2 |
| (9Z,12Z,15Z)-octadeca-9,12,15-trienoic acid (linolenic acid) | C18:3 ω3 | 1.0 ± 0.1 |
| Total polyunsaturated fatty acids (PUFAs) | 7.4 ± 0.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Nicola, G.R.; Montaut, S.; Leclair, K.; Garrioux, J.; Guillot, X.; Rollin, P. Cultivated Winter-Type Lunaria annua L. Seed: Deciphering the Glucosinolate Profile Integrating HPLC, LC-MS and GC-MS Analyses, and Determination of Fatty Acid Composition. Molecules 2024, 29, 3803. https://doi.org/10.3390/molecules29163803
De Nicola GR, Montaut S, Leclair K, Garrioux J, Guillot X, Rollin P. Cultivated Winter-Type Lunaria annua L. Seed: Deciphering the Glucosinolate Profile Integrating HPLC, LC-MS and GC-MS Analyses, and Determination of Fatty Acid Composition. Molecules. 2024; 29(16):3803. https://doi.org/10.3390/molecules29163803
Chicago/Turabian StyleDe Nicola, Gina Rosalinda, Sabine Montaut, Kayla Leclair, Joëlle Garrioux, Xavier Guillot, and Patrick Rollin. 2024. "Cultivated Winter-Type Lunaria annua L. Seed: Deciphering the Glucosinolate Profile Integrating HPLC, LC-MS and GC-MS Analyses, and Determination of Fatty Acid Composition" Molecules 29, no. 16: 3803. https://doi.org/10.3390/molecules29163803
APA StyleDe Nicola, G. R., Montaut, S., Leclair, K., Garrioux, J., Guillot, X., & Rollin, P. (2024). Cultivated Winter-Type Lunaria annua L. Seed: Deciphering the Glucosinolate Profile Integrating HPLC, LC-MS and GC-MS Analyses, and Determination of Fatty Acid Composition. Molecules, 29(16), 3803. https://doi.org/10.3390/molecules29163803