
Dehydroevodiamine Alleviates Ulcerative Colitis by Inhibiting the PI3K/AKT/NF-κB Signaling Pathway via Targeting AKT1 and Regulating Gut Microbes and Serum Metabolism


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
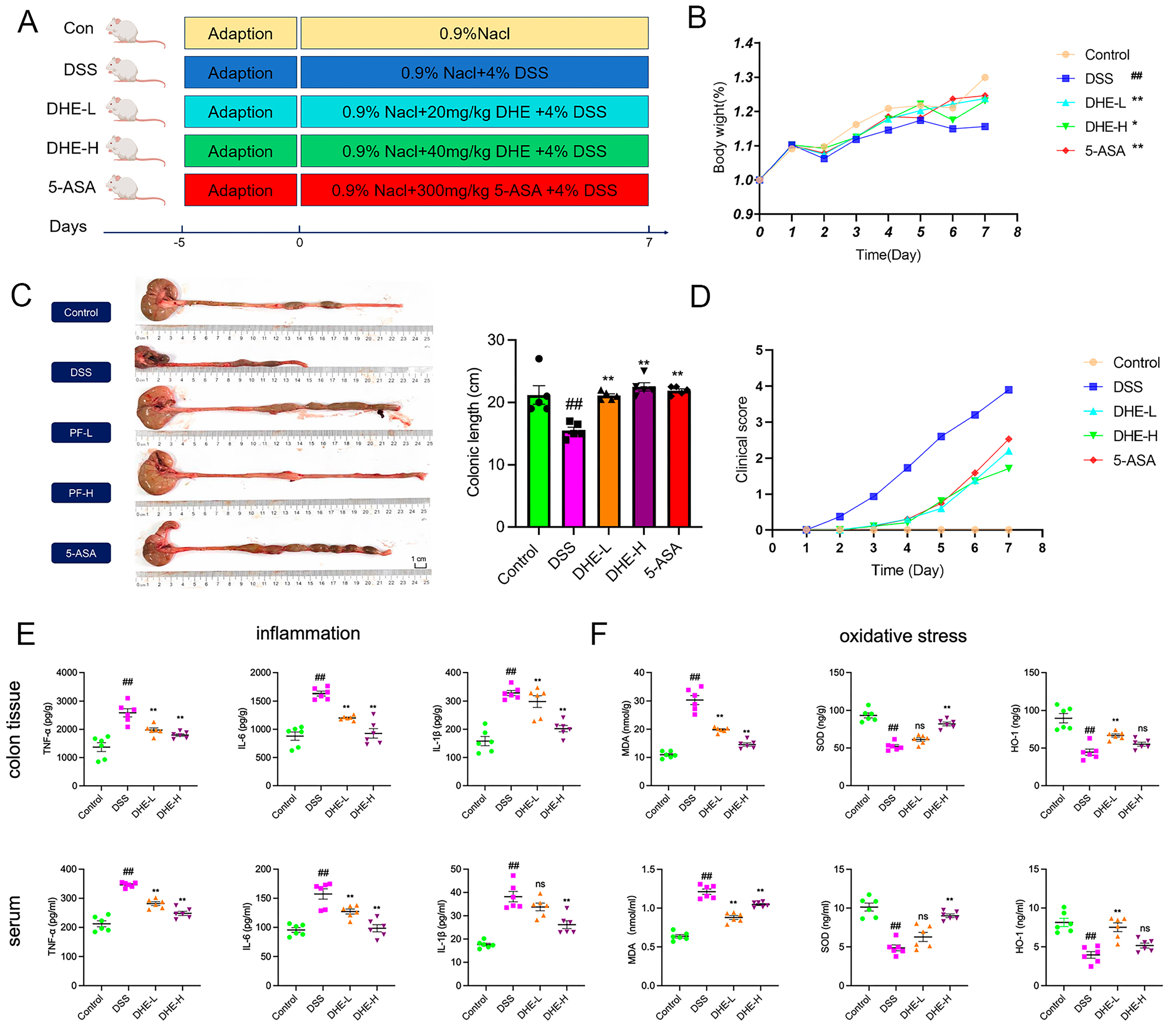
2.1. DHE Relieved Symptoms of Colitis in UC Rats
2.2. DHE Reducing Inflammation and Oxidative Stress in UC Rats
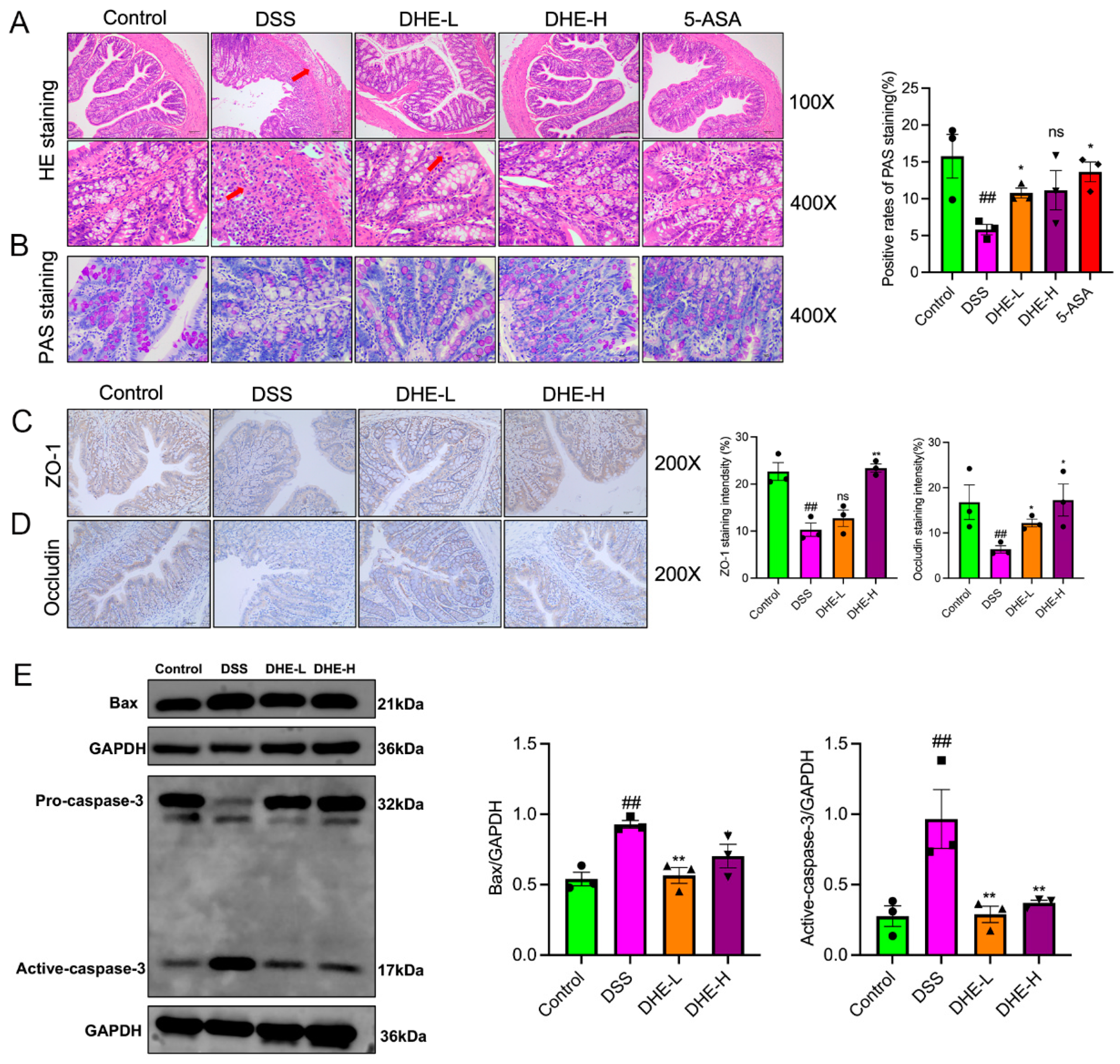
2.3. DHE Repaired the Intestinal Mucosal Barrier and Inhibited Apoptosis in UC Rats
2.4. DHE Mitigated Deregulation of Gut Microbes Induced by DSS in UC Rats
2.5. DHE Modulated DSS-Induced Alterations in Serum Metabolites of Rats
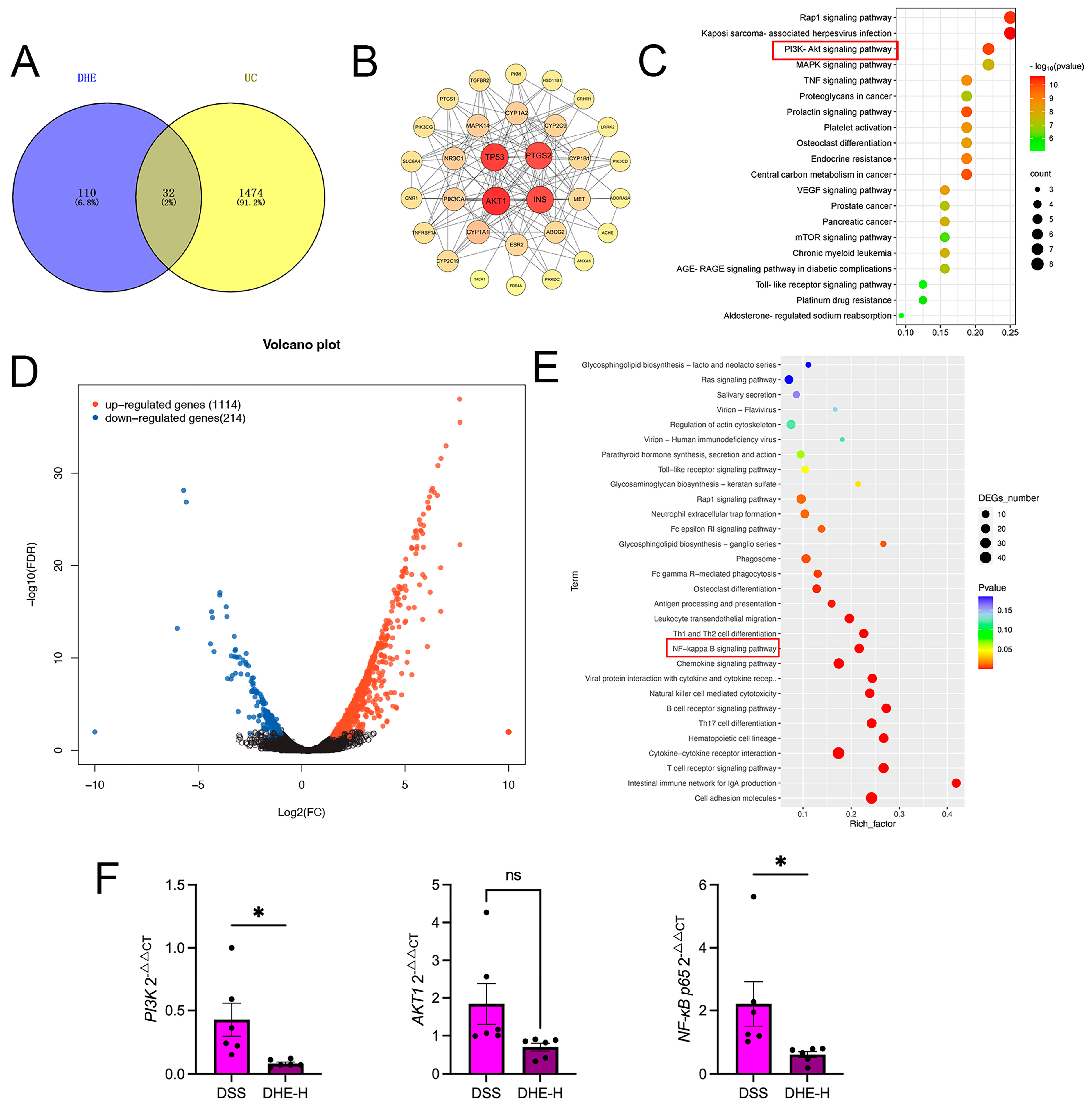
2.6. Integrated Network Pharmacology with RNA Sequencing Revealed the Mechanisms of DHE on UC
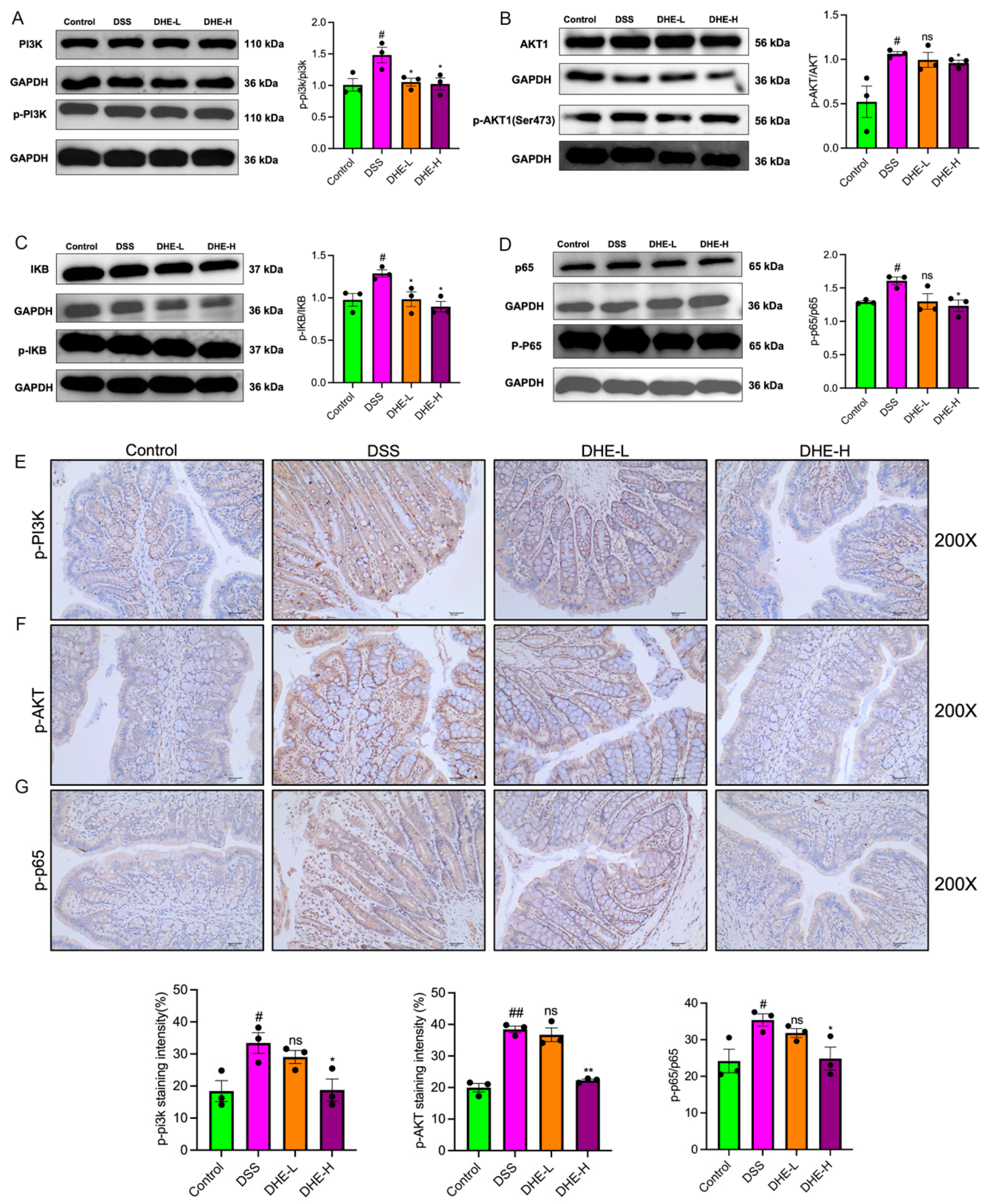
2.7. DHE-Suppressed DSS Mediated the Activation of PI3K/AKT/NF-κB Signaling Pathway
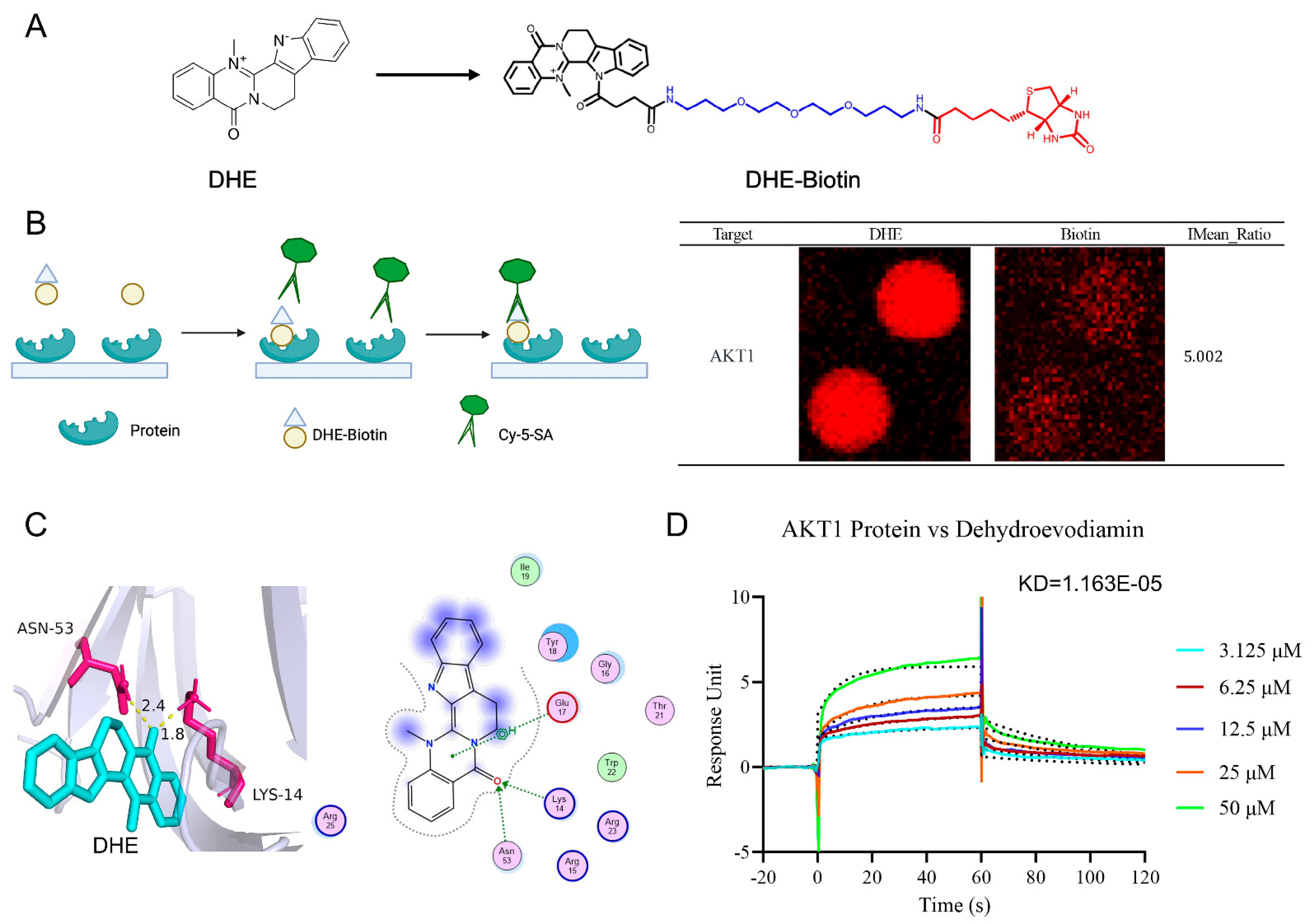
2.8. HuProtTM20K Proteomics Revealed Direct Binding of DHE to AKT1
3. Discussion
4. Methodology and Experimental Materials
4.1. Chemicals
4.2. Establishment of UC Rat Model and Drug Administration Process
4.3. Measurement of Inflammatory and Oxidative Stress Factors by Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Hematoxylin–Eosin (HE) Staining
4.5. AB-PAS Staining
4.6. Immunohistochemistry (IHC) Assay
4.7. RT-qPCR Assay
4.8. Western Blotting (WB) Assay
4.9. 16S rRNA Sequencing
4.10. Metabolomics Assay
4.11. Network Pharmacology
4.12. RNA Sequencing
4.13. HuProtTM20K Proteomics
4.14. Molecular Docking
4.15. Surface Plasmon Resonance (SPR) Assay
4.16. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Du, L.; Ha, C. Epidemiology and Pathogenesis of Ulcerative Colitis. Gastroenterol. Clin. N. Am. 2020, 49, 643–654. [Google Scholar] [CrossRef]
- Segal, J.P.; LeBlanc, J.-F.; Hart, A.L. Ulcerative Colitis: An Update. Clin. Med. 2021, 21, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Le Berre, C.; Honap, S.; Peyrin-Biroulet, L. Ulcerative Colitis. Lancet 2023, 402, 571–584. [Google Scholar] [CrossRef]
- Nakase, H.; Sato, N.; Mizuno, N.; Ikawa, Y. The Influence of Cytokines on the Complex Pathology of Ulcerative Colitis. Autoimmun. Rev. 2022, 21, 103017. [Google Scholar] [CrossRef]
- Deng, B.; Liao, F.; Liu, Y.; He, P.; Wei, S.; Liu, C.; Dong, W. Comprehensive Analysis of Endoplasmic Reticulum Stress-Associated Genes Signature of Ulcerative Colitis. Front. Immunol. 2023, 14, 1158648. [Google Scholar] [CrossRef]
- Li, C.; Wu, Y.; Xie, Y.; Zhang, Y.; Jiang, S.; Wang, J.; Luo, X.; Chen, Q. Oral Manifestations Serve as Potential Signs of Ulcerative Colitis: A Review. Front. Immunol. 2022, 13, 1013900. [Google Scholar] [CrossRef]
- Calméjane, L.; Laharie, D.; Kirchgesner, J.; Uzzan, M. Review Article: Updated Management of Acute Severe Ulcerative Colitis: From Steroids to Novel Medical Strategies. United Eur. Gastroenterol. J. 2023, 11, 722–732. [Google Scholar] [CrossRef]
- Ochoa, B.; McMahon, L. Surgery for Ulcerative Colitis. Semin. Pediatr. Surg. 2024, 33, 151404. [Google Scholar] [CrossRef] [PubMed]
- Gros, B.; Kaplan, G.G. Ulcerative Colitis in Adults. JAMA 2023, 330, 951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, X.; Zhang, Q.; Yang, J.; Liu, G. Roles of Macrophages on Ulcerative Colitis and Colitis-Associated Colorectal Cancer. Front. Immunol. 2023, 14, 1103617. [Google Scholar] [CrossRef] [PubMed]
- Krugliak Cleveland, N.; Torres, J.; Rubin, D.T. What Does Disease Progression Look Like in Ulcerative Colitis, and How Might It Be Prevented? Gastroenterology 2022, 162, 1396–1408. [Google Scholar] [CrossRef]
- Kucharzik, T.; Koletzko, S.; Kannengießer, K.; Dignaß, A. Ulcerative Colitis—Diagnostic and Therapeutic Algorithms. Dtsch. Arztebl. Int. 2020, 117, 564–573. [Google Scholar] [CrossRef]
- Cottone, M.; Renna, S.; Modesto, I.; Orlando, A. Is 5-ASA Still the Treatment of Choice for Ulcerative Colitis? Curr. Drug Targets 2011, 12, 1396–1405. [Google Scholar] [CrossRef] [PubMed]
- Noh, E.J.; Ahn, K.S.; Shin, E.M.; Jung, S.H.; Kim, Y.S. Inhibition of Lipopolysaccharide-Induced INOS and COX-2 Expression by Dehydroevodiamine through Suppression of NF-ΚB Activation in RAW 264.7 Macrophages. Life Sci. 2006, 79, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhao, Y.; Gong, M.; He, Z.; Wang, W.; Li, Y.; Zhai, W.; Yu, Z. Dehydroevodiamine Ameliorates Neurological Dysfunction after Traumatic Brain Injury in Mice via Regulating the SIRT1/FOXO3a/Bim Pathway. Phytomedicine 2024, 125, 155321. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.D.; Han, Y.-H.; Mun, J.-G.; Yoon, D.H.; Lee, Y.G.; Kee, J.-Y.; Hong, S.-H. Dehydroevodiamine Inhibits Lung Metastasis by Suppressing Survival and Metastatic Abilities of Colorectal Cancer Cells. Phytomedicine 2022, 96, 153809. [Google Scholar] [CrossRef]
- Zhu, S.-L.; Qi, M.; Chen, M.-T.; Lin, J.-P.; Huang, H.-F.; Deng, L.-J.; Zhou, X.-W. A Novel DDIT3 Activator Dehydroevodiamine Effectively Inhibits Tumor Growth and Tumor Cell Stemness in Pancreatic Cancer. Phytomedicine 2024, 128, 155377. [Google Scholar] [CrossRef]
- Wen, J.; Tong, Y.; Ma, X.; Wang, R.; Li, R.; Song, H.; Zhao, Y. Therapeutic Effects and Potential Mechanism of Dehydroevodiamine on N-Methyl-N′-Nitro-N-Nitrosoguanidine-Induced Chronic Atrophic Gastritis. Phytomedicine 2021, 91, 153619. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Chen, L.; He, Y.; He, T.; Wang, R.; Wei, S.; Jing, M.; Zhou, X.; Li, H.; et al. Integrated Network Pharmacology and Metabolomics to Investigate the Effects and Possible Mechanisms of Dehydroevodiamine against Ethanol-Induced Gastric Ulcers. J. Ethnopharmacol. 2024, 319, 117340. [Google Scholar] [CrossRef]
- Gajendran, M.; Loganathan, P.; Jimenez, G.; Catinella, A.P.; Ng, N.; Umapathy, C.; Ziade, N.; Hashash, J.G. A Comprehensive Review and Update on Ulcerative Colitis. Disease-a-Month 2019, 65, 100851. [Google Scholar] [CrossRef]
- Guo, X.Y.; Liu, X.J.; Hao, J.Y. Gut Microbiota in Ulcerative Colitis: Insights on Pathogenesis and Treatment. J. Dig. Dis. 2020, 21, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, D.; Mao, M.; Liu, W.; Wang, Y.; Liang, Y.; Cao, W.; Zhong, X. Betaine Ameliorates Acute Sever Ulcerative Colitis by Inhibiting Oxidative Stress Induced Inflammatory Pyroptosis. Mol. Nutr. Food Res. 2022, 66, 2200341. [Google Scholar] [CrossRef] [PubMed]
- van Muijlwijk, G.H.; Rice, T.A.; Flavell, R.A.; Palm, N.W.; de Zoete, M.R. Allobaculum Mucilyticum Sp. Nov. and Allobaculum Fili Sp. Nov., Isolated from the Human Intestinal Tract. Int. J. Syst. Evol. Microbiol. 2023, 73, 005635. [Google Scholar] [CrossRef] [PubMed]
- Alhobayb, T.; Ciorba, M.A. Clostridium Difficile in Inflammatory Bowel Disease. Curr. Opin. Gastroenterol. 2023, 39, 257–262. [Google Scholar] [CrossRef]
- Seishima, J.; Iida, N.; Kitamura, K.; Yutani, M.; Wang, Z.; Seki, A.; Yamashita, T.; Sakai, Y.; Honda, M.; Yamashita, T.; et al. Gut-Derived Enterococcus Faecium from Ulcerative Colitis Patients Promotes Colitis in a Genetically Susceptible Mouse Host. Genome Biol. 2019, 20, 252. [Google Scholar] [CrossRef]
- Jia, D.-J.-C.; Wang, Q.-W.; Hu, Y.-Y.; He, J.-M.; Ge, Q.-W.; Qi, Y.-D.; Chen, L.-Y.; Zhang, Y.; Fan, L.-N.; Lin, Y.-F.; et al. Lactobacillus Johnsonii Alleviates Colitis by TLR1/2-STAT3 Mediated CD206 + Macrophages IL−10 Activation. Gut Microbes 2022, 14, 2145843. [Google Scholar] [CrossRef]
- Yao, S.; Zhao, Z.; Wang, W.; Liu, X. Bifidobacterium Longum: Protection against Inflammatory Bowel Disease. J. Immunol. Res. 2021, 2021, 8030297. [Google Scholar] [CrossRef]
- Han, T.; Hu, X.; Li, K.; Zhang, D.; Zhang, Y.; Li, J. Bifidobacterium Infantis Maintains Genome Stability in Ulcerative Colitis via Regulating Anaphase-Promoting Complex Subunit 7. Front. Microbiol. 2021, 12, 761113. [Google Scholar] [CrossRef]
- Dou, W.; Xu, H.; Feng, S.; Liu, T.; Xiao, L.; Wu, Y.; Chen, S.; Pan, Y.; Wang, X. The Alleviating Effects and Mechanisms of Betaine on Dextran Sulfate Sodium-Induced Colitis in Mice. Mol. Nutr. Food Res. 2023, 67, 2300376. [Google Scholar] [CrossRef]
- Ye, Z.; Li, Y.; She, Y.; Wu, M.; Hu, Y.; Qin, K.; Li, L.; Yu, H.; Zhao, Q.; Jin, Z.; et al. Renshen Baidu Powder Protects Ulcerative Colitis via Inhibiting the PI3K/Akt/NF-ΚB Signaling Pathway. Front. Pharmacol. 2022, 13, 880589. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, C.; Cao, J.; Bei, G.; Wang, X.; Miao, Z. Yiyi Fuzi Baijiang Decoction Alleviates Ulcerative Colitis Partly by Regulating TLR4-Mediated PI3K/Akt and NF-ΚB Pathways. Evid.-Based Complement. Altern. Med. 2022, 2022, 8780514. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gong, L.; Jiang, Y.; Huo, X.; Huang, L.; Lei, H.; Gu, Y.; Wang, D.; Guo, D.; Deng, Y. Sanguisorba Officinalis Ethyl Acetate Extract Attenuates Ulcerative Colitis through Inhibiting PI3K-AKT/NF-ΚB/STAT3 Pathway Uncovered by Single-Cell RNA Sequencing. Phytomedicine 2023, 120, 155052. [Google Scholar] [CrossRef] [PubMed]
- Siddika, T.; Balasuriya, N.; Frederick, M.I.; Rozik, P.; Heinemann, I.U.; O’Donoghue, P. Delivery of Active AKT1 to Human Cells. Cells 2022, 11, 3834. [Google Scholar] [CrossRef] [PubMed]
- George, B.; Gui, B.; Raguraman, R.; Paul, A.M.; Nakshatri, H.; Pillai, M.R.; Kumar, R. AKT1 Transcriptomic Landscape in Breast Cancer Cells. Cells 2022, 11, 2290. [Google Scholar] [CrossRef]
- Guo, H.-X.; Wang, B.-B.; Wu, H.-Y.; Feng, H.-Y.; Zhang, H.-Y.; Gao, W.; Yuan, B. Turtle Peptide and Its Derivative Peptide Ameliorated DSS-Induced Ulcerative Colitis by Inhibiting Inflammation and Modulating the Composition of the Gut Microbiota. Int. Immunopharmacol. 2024, 132, 112024. [Google Scholar] [CrossRef]
- Yang, H.-B.; Song, J.-Y.; Xu, C.; Li, J.; Zhang, C.; Xie, S.; Teng, C. Interventional Effects of Pueraria Oral Liquid on T2DM Rats and Metabolomics Analysis. Biomed. Pharmacother. 2024, 175, 116780. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Hu, Q.; Jiang, T.; Chen, Y.; Zhang, W.; Gao, P.; Zeng, J.; Efferth, T. Dehydroevodiamine Alleviates Ulcerative Colitis by Inhibiting the PI3K/AKT/NF-κB Signaling Pathway via Targeting AKT1 and Regulating Gut Microbes and Serum Metabolism. Molecules 2024, 29, 4031. https://doi.org/10.3390/molecules29174031
Ma X, Hu Q, Jiang T, Chen Y, Zhang W, Gao P, Zeng J, Efferth T. Dehydroevodiamine Alleviates Ulcerative Colitis by Inhibiting the PI3K/AKT/NF-κB Signaling Pathway via Targeting AKT1 and Regulating Gut Microbes and Serum Metabolism. Molecules. 2024; 29(17):4031. https://doi.org/10.3390/molecules29174031
Chicago/Turabian StyleMa, Xiao, Qichao Hu, Tao Jiang, Yuan Chen, Wenwen Zhang, Pan Gao, Jinhao Zeng, and Thomas Efferth. 2024. "Dehydroevodiamine Alleviates Ulcerative Colitis by Inhibiting the PI3K/AKT/NF-κB Signaling Pathway via Targeting AKT1 and Regulating Gut Microbes and Serum Metabolism" Molecules 29, no. 17: 4031. https://doi.org/10.3390/molecules29174031