

In Situ Antimicrobial Properties of Sabinene Hydrate, a Secondary Plant Metabolite

Abstract
:1. Introduction
- (i)
- blue-stain fungi: Ceratocystis polonica, Ophiostoma bicolor and O. penicillatum (syn. Grosmannia penicillata)), frequently originating from bark beetle galleries;
- (ii)
- other fungi associated with Ips. typographus, i.e., three fungal strains: Musicillium theobromae, Plectosphaerella cucumerina and Trichoderma sp., isolated from sapwood underneath bark beetle galleries (Ips typographus) on spruce (Picea abies) stems;
- (iii)
- Verticillium fungicola, isolated from diseased I. typographus larvae;
- (iv)
- and to compare SH activity against:
- (v)
- two Gram-positive bacteria (Bacillus subtilis and Staphylococcus aureus),
- (vi)
- two Gram-negative bacteria (Escherichia coli and Pseudomonas aeruginosa);
- (vii)
- five yeasts (Candida albicans, C. krusei, C. parapsilosis, Saccharomyces cerevisiae, and Rhodotorula muscilaginosa),
- (viii)
- two saprophytic fungi (Aspergillus niger and Penicillium notatum).
2. Results
2.1. Agar Disc Diffusion Method
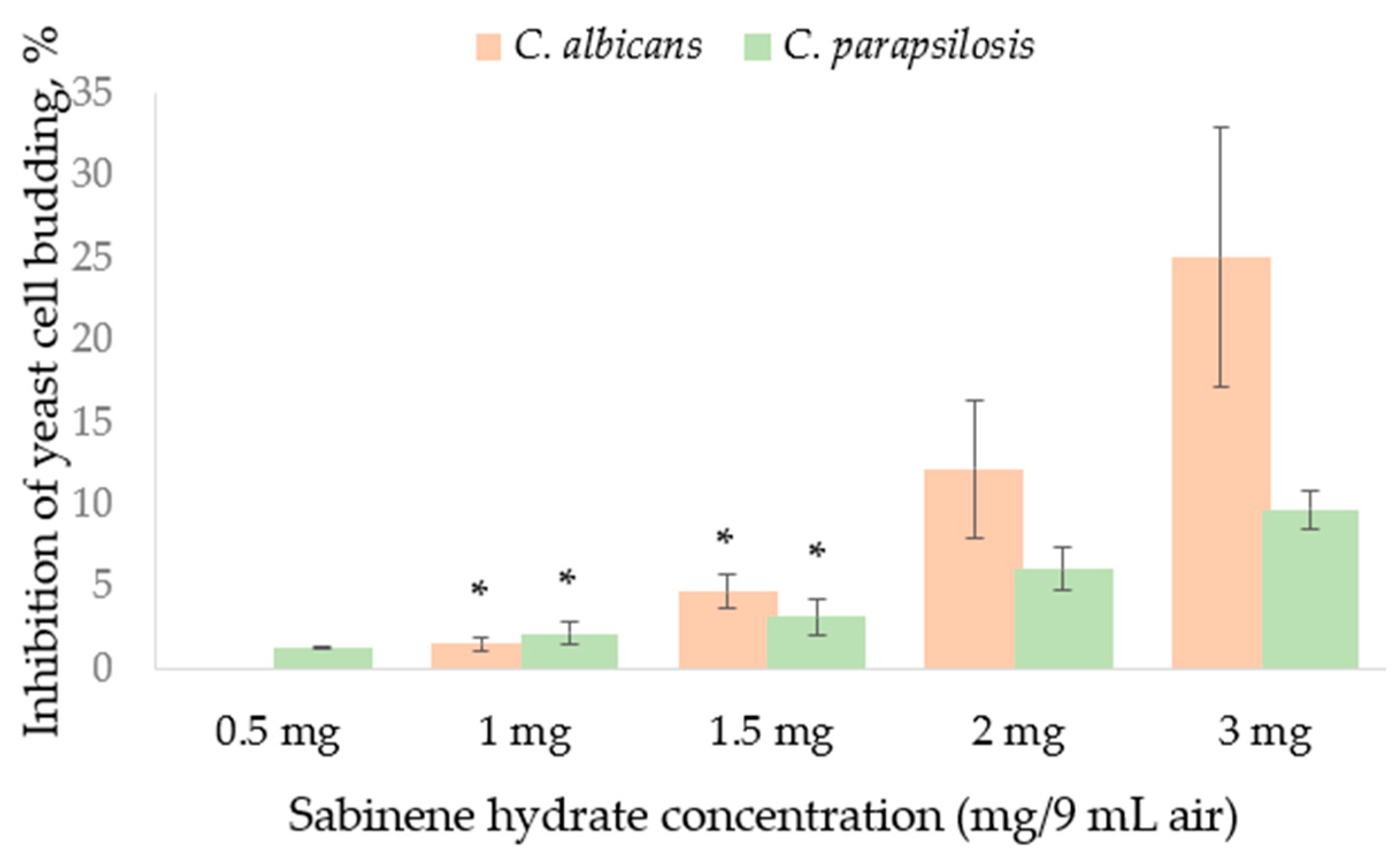
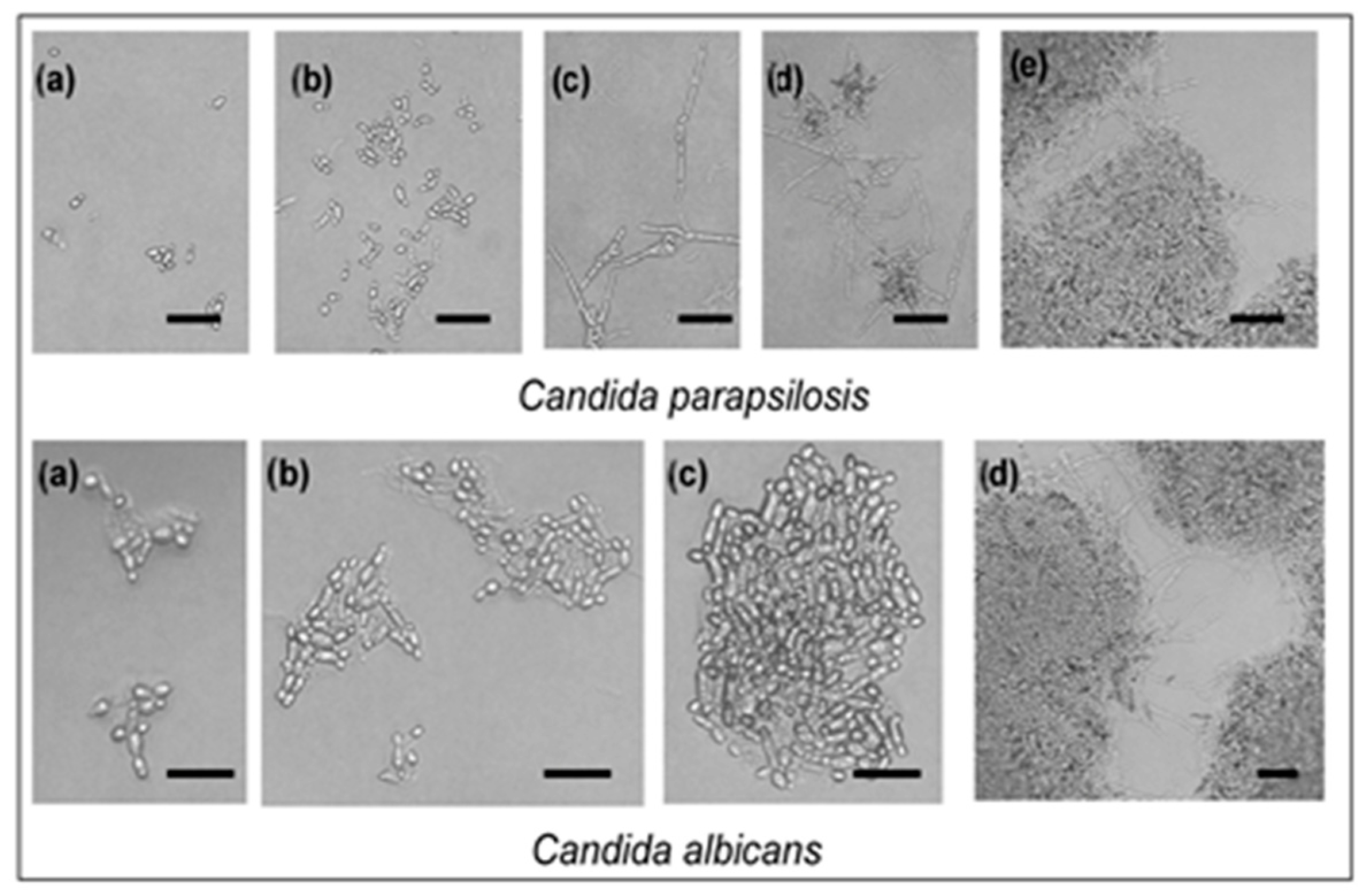
2.2. Broth Macro-Dilution Method
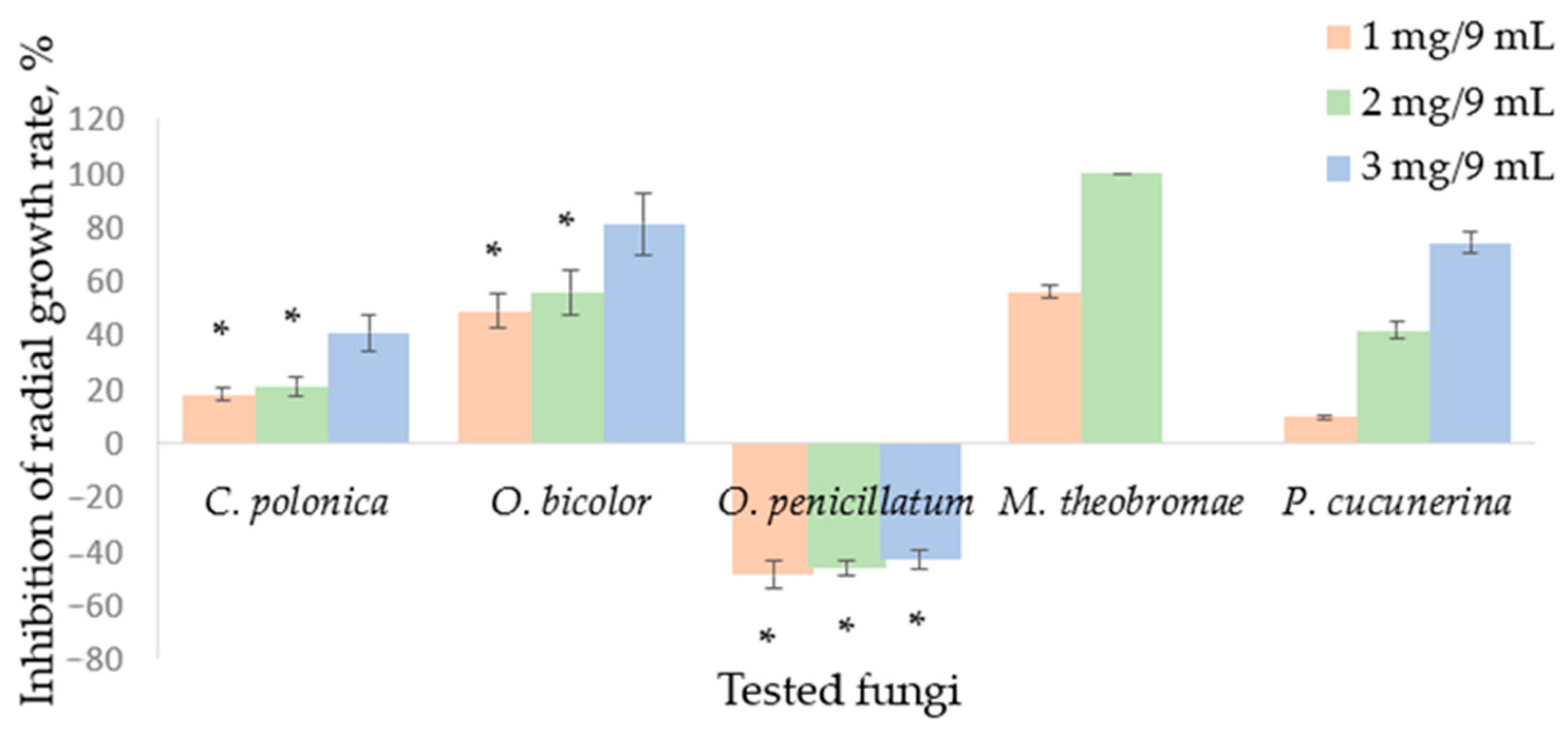
2.3. Sabinene Hydrate Volatilization from Paper Disc (Vapour Phase Activity Test)
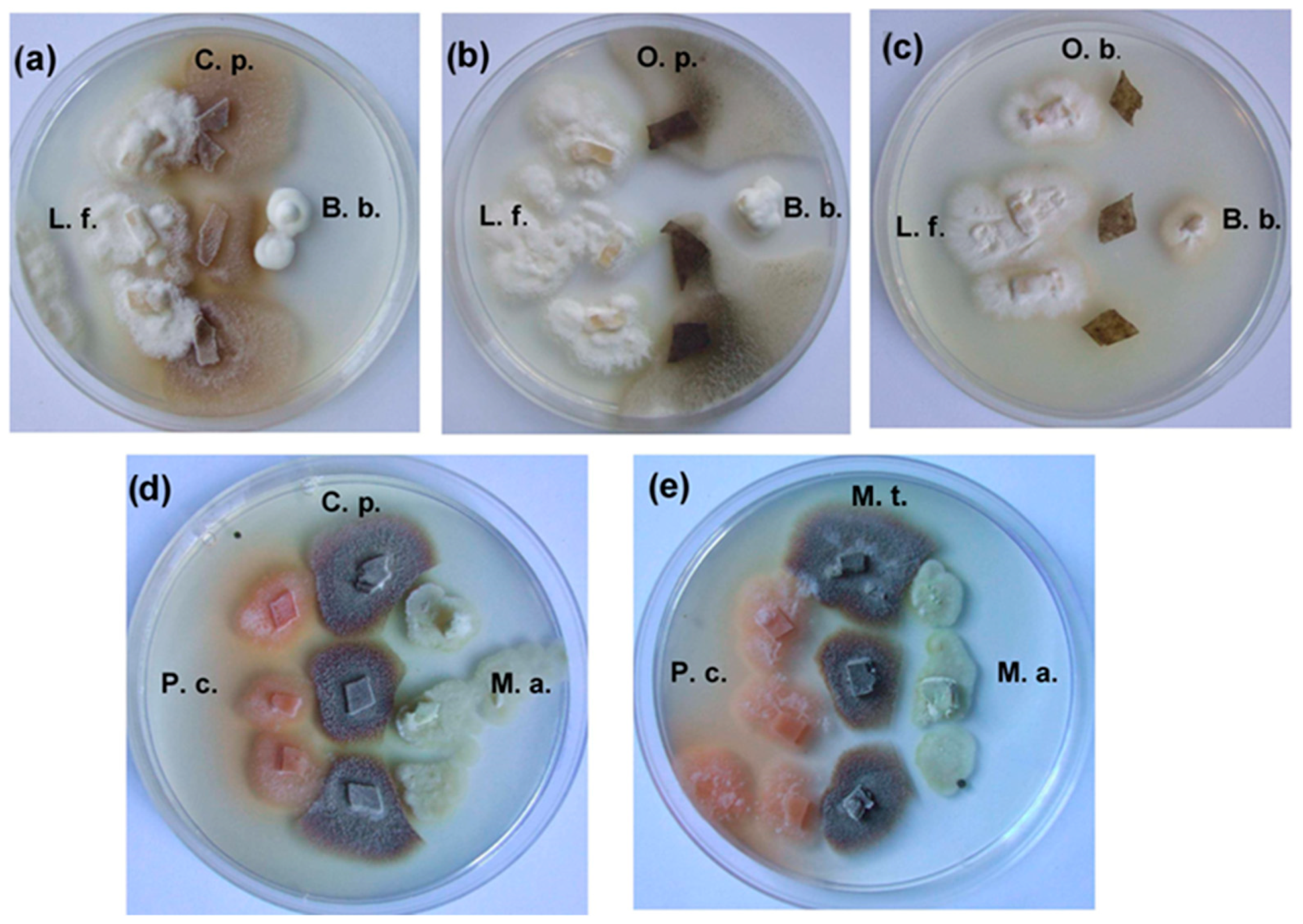
2.4. Results of Dual Culture Tests
3. Discussion
4. Materials and Methods
4.1. Sabinene Hydrate (Dissolution and Preparation for Bioassays)
4.2. Test Organisms
- (a)
- blue-stain fungi: Ceratocystis polonica (Siem.) C. Moreau 1994-169/113, C. polonica BIGTC-2133, Ophiostoma bicolor Davidson & Wells BIGTC-2133, and O. penicillatum (Grosmann.) Siemaszko (syn. Grosmannia penicillata (Grosmann) Goid) 2006-209/44/2;
- (b)
- other mycelial fungi, which were isolated from Ips typographus Linnaeous (Coleoptera, Curculionidae, Scolytinae) galleries in Picea abies L. stemps and from its discolorated sapwood, such as Musicillium theobromae (Turconi) Zare & W. Gams (syn. Stachylidium theobromae Turconi, Verticillium theobromae E.W. Mason & S. Hughes BIGTC-20132 and Plectosphaerella cucumerina (Lindt.) W. Gams BIGTC-136, Aspergillus niger Tiegh. BIGTC-9823, Penicillium notatum Wehmer BIGTC-8914, and Trichoderma sp. BIGTC-2131;
- (c)
- yeasts Candida albicans BIGTC-MK2, C. parapsilosis BIGTC-MK9, C. krusei BIGTC-MK3 (Blastomycetes, Cryptococcales, Cryptococcaceae) and Saccharomyces cerevisiae BIGTC-MK11 (Saccharomycetes, Saccharomycetales, Saccharomycetaceae) phylogenetically related to the Ascomycota, as well as Rhodotorula muscilaginosa BIGTC-056 (Urediomycetes, Sporidiales) phylogenetically related to Basidiomycota;
- (d)
- Gram-positive bacteria Staphylococcus aureus BIGTC-BK06 and Bacillus subtilis BIGTC-BK09;
- (e)
- Gram-negative bacteria Escherichia coli BIGTC-BK08 and Pseudomonas aeruginosa BIGTC-BK19.
4.3. Preparation of Test Organisms
4.4. In Vitro Antifungal and Antibacterial Activity Testing
4.4.1. Agar Disk Diffusion Method
4.4.2. Broth Macro-Dilution Tests
4.4.3. Disc Volatilization Method (Vapor Phase Activity)
4.4.4. Dual Culture Tests
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rehman, R.; Hanif, M.A.; Mushtaq, Z.; Al-Sadi, A.M. Biosynthesis of essential oils in aromatic plants: A review. Food Rev. Int. 2016, 32, 117–160. [Google Scholar] [CrossRef]
- Hallachan, T.W.; Croteau, R. Monoterpene biosynthesis: Demonstration of a geranyl pyrophosphate: Sabinene hydrate cyclase in soluble enzyme preparations from sweet marjoram (Majorana hortensis). Arch. Biochem. Biophys. 1988, 264, 618–631. [Google Scholar] [CrossRef]
- Arsenijević, J.; Marković, J.; Šoštarić, I.; Ražić, S. A chemometrics as a powerful tool in the elucidation of the role of metals in the biosynthesis of volatile organic compounds in Hungarian thyme samples. Plant Physiol. Biochem. 2013, 71, 298–306. [Google Scholar] [CrossRef]
- Krause, S.T.; Köllner, T.G.; Asbach, J.; Degenhardt, J. Stereochemical mechanism of two sabinene hydrate synthases forming antipodal monoterpenes in thyme (Thymus vulgaris). Arch. Biochem. Biophys. 2013, 529, 112–121. [Google Scholar] [CrossRef]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile compounds in Citrus essential oils: A comprehensive review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Hernández, M.G.; Sánchez-Bravo, P.; Hernández, F.; Carbonell-Barrachina, Á.A.; Pastor-Pérez, J.J.; Legua, P. Determination of the volatile profile of lemon peel oils as affected by rootstock. Foods 2020, 9, 241. [Google Scholar] [CrossRef]
- Flamini, G.; Najar, B.; Leonardi, M.; Ambryszewska, K.E.; Cioni, P.L.; Parri, F.; Melai, B.; Pistelli, L. Essential oil composition of Salvia rosmarinus Spenn. wild samples collected from six sites and different seasonal periods in Elba Island (Tuscan Archipelago, Italy). Nat. Prod. Res. 2020, 36, 1919–1925. [Google Scholar] [CrossRef]
- Matulyte, I.; Marksa, M.; Ivanauskas, L.; Kalvėnienė, Z.; Lazauskas, R.; Bernatonienė, J. GC-MS analysis of the composition of the extracts and essential oil from Myristica fragrans seeds using magnesium aluminometasilicate as excipient. Molecules 2019, 24, 1062. [Google Scholar] [CrossRef]
- Adiani, V.; Gupta, S.; Chatterjee, S.; Variyar, P.S.; Sharma, A. Activity guided characterization of antioxidant components from essential oil of Nutmeg (Myristica fragrans). J. Food Sci. Technol. 2013, 52, 221–230. [Google Scholar] [CrossRef]
- Kapoor, I.P.S.; Singh, B.; Singh, G.; De Heluani, C.S.; De Lampasona, M.P.; Catalan, C.A.N. Chemical composition and antioxidant activity of essential oil and oleoresins of nutmeg (Myristica fragrans Houtt.) fruits. Int. J. Food Prop. 2013, 16, 1059–1070. [Google Scholar] [CrossRef]
- Danin, A.; Ravid, U.; Umano, K.; Shibamoto, T. Essential oil composition of Origanum ramonense Danin leaves from Israel. J. Essent. Oil Res. 1997, 9, 411–417. [Google Scholar] [CrossRef]
- Ghazal, T.S.A.; Schelz, Z.; Vidács, L.; Szemerédi, N.; Veres, K.; Spengler, G.; Hohmann, J. Antimicrobial, multidrug resistance reversal and biofilm formation inhibitory effect of Origanum majorana extracts, essential oil and monoterpenes. Plants 2022, 11, 1432. [Google Scholar] [CrossRef] [PubMed]
- Arafat, K.; Al-Azawi, A.M.; Sulaiman, S.; Attoub, S. Exploring the anticancer potential of Origanum majorana essential oil monoterpenes alone and in combination against non-small cell lung cancer. Nutrients 2023, 15, 5010. [Google Scholar] [CrossRef] [PubMed]
- Dudai, N.; Larkov, O.; Chaimovitsh, D.; Lewinsohn, E.; Freiman, L.; Ravid, U. Essential oil compounds of Origanum dayi Post. Flavour. Fragr. J. 2003, 18, 334–337. [Google Scholar] [CrossRef]
- Tabanca, N.; Özek, T.; Baser, K.H.C. Comparison of the essential oils of Origanum majorana L. and Origanum x majoricum Cambess. J. Essent. Oil Res. 2004, 16, 248–252. [Google Scholar] [CrossRef]
- Raina, A.P.; Negi, K.S. Essential oil composition of Origanum majorana and Origanum vulgare ssp. hirtum growing in India. Chem. Nat. Compd. 2012, 47, 1015–1017. [Google Scholar] [CrossRef]
- Lukas, B.; Schmiderer, C.; Novak, J. Essential oil diversity of European Origanum vulgare L. (Lamiaceae). Phytochemistry 2015, 119, 32–40. [Google Scholar] [CrossRef]
- Mohamed, A.A.; El-Hefny, M.; El-Shanhorey, N.A.; Ali, H.M. Foliar application of bio-stimulants enhancing the production and the toxicity of Origanum majorana essential oils against four rice seed-borne fungi. Molecules 2020, 25, 2363. [Google Scholar] [CrossRef] [PubMed]
- Simirgiotis, M.J.; Burton, D.; Parra, F.; López, J.; Muñoz, P.; Escobar, H.; Parra, C. Antioxidant and antibacterial capacities of Origanum vulgare L. essential oil from the Arid Andean region of Chile and its chemical characterization by GC-MS. Metabolites 2020, 10, 414. [Google Scholar] [CrossRef]
- Piasecki, B.; Balázs, V.L.; Kieltyka-Dadasiewicz, A.; Szabó, P.; Kocsis, B.; Horváth, G.; Ludwiczuk, A. Microbiological studies on the influence of essential oils from several Origanum species on respiratory pathogens. Molecules 2023, 28, 3044. [Google Scholar] [CrossRef]
- Kandoudi, W.; Radácsi, P.; Gosztola, B.; Zámboriné Németh, É. Elicitation of medicinal plants in vivo—Is it a realistic tool? The effect of methyl jasmonate and salicylic acid on Lamiaceae species. Horticulturae 2022, 8, 5. [Google Scholar] [CrossRef]
- Farouk, A.; Mohsen, M.; Ali, H.; Shaaban, H.; Albaridi, N. Antioxidant activity and molecular docking study of volatile constituents from different aromatic Lamiaceous plants cultivated in Madinah Monawara, Saudi Arabia. Molecules 2021, 26, 4145. [Google Scholar] [CrossRef]
- Satyal, P.; Murray, B.L.; McFeeters, R.L.; Setzer, W.N. Essential oil characterization of Thymus vulgaris from various geographical locations. Foods 2016, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Bumbulytė, G.; Būdienė, J.; Būda, V. Essential oils and their components control behaviour of yellow mealworm (Tenebrio molitor) larvae. Insects 2023, 14, 636. [Google Scholar] [CrossRef]
- Groendahl, E.; Ehlers, B.K.; Keefover-Ring, E. A new cis-sabinene hydrate chemotype detected in large thyme Thymus pulegioides L. growing wild in Denmark. J. Essent. Oil Res. 2008, 20, 40–41. [Google Scholar] [CrossRef]
- Kim, S.-W.; Lee, H.-R.; Jang, M.-J.; Jung, C.-S.; Park, I.-K. Fumigant toxicity of Lamiaceae plant essential oils and blends of their constituents against adult rice weevil Sitophilus oryzae. Molecules 2016, 21, 361. [Google Scholar] [CrossRef]
- Trendafilova, A.; Todorova, M.; Ivanova, V.; Zhelev, P.; Aneva, I. Essential oil composition of ten species from sect. Serpyllum of genus Thymus growing in Bulgaria. Diversity 2023, 15, 759. [Google Scholar] [CrossRef]
- Wiese, N.; Fischer, J.; Heidler, J.; Lewkowski, O.; Degenhardt, J.; Erler, S. The terpenes of leaves, pollen, and nectar of thyme (Thymus vulgaris) inhibit growth of bee disease-associated microbes. Sci. Rep. 2018, 8, 14634. [Google Scholar] [CrossRef]
- Bayan, Y.; Küsek, M. Chemical composition and antifungal and antibacterial activity of Mentha spicata L. volatile oil. Cien. Inv. Agr. 2018, 45, 64–69. [Google Scholar] [CrossRef]
- Kumar, A.; Yadav, L.B.S.; Ahmad, J.; Dubey, N.; Puri, S. Chemical composition of commercial Juniperus communis L. leaf oil. J. Essent. Oil Bear. Plants 2013, 10, 310–313. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Astatkie, T.; Jeliazkova, E.A.; Tatman, A.O.; Schlegel, V. Distillation time alters essential oil yield, composition and antioxidant activity of female Juniperus scopulorum trees. J. Essent. Oil Res. 2013, 25, 62–69. [Google Scholar] [CrossRef]
- Cornwell, C.P.; Leach, D.N.; Wyllie, S.W. The origin of terpinen-4-ol in the steam distillates of Melaleuca argentea, M. dissitiflora and M. linariifolia. J. Essent. Oil Res. 1999, 11, 49–53. [Google Scholar] [CrossRef]
- Valdivieso-Ugarte, M.; Gomez-Llorente, C.; Plaza-Díaz, J.; Gil, Á. Antimicrobial, antioxidant, and immunomodulatory properties of essential oils: A systematic review. Nutrients 2019, 11, 2786. [Google Scholar] [CrossRef]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial activity of some essential oils—Present status and future perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Alternat. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef]
- Rathore, S.; Mukhia, S.; Kumar, R.; Kumar, R. Essential oil composition and antimicrobial potential of aromatic plants grown in the mid-hill conditions of the Western Himalayas. Sci. Rep. 2023, 13, 4878. [Google Scholar] [CrossRef]
- Santoyo, S.; Cavero, S.; Jaime, L.; Ibañez, E.; Señoráns, F.J.; Reglero, G. Supercritical carbon dioxide extraction of compounds with antimicrobial activity from Origanum vulgare L.: Determination of optimal extraction parameters. J. Food Prot. 2006, 69, 369–375. [Google Scholar] [CrossRef]
- Radaelli, M.; da Silvaa, B.P.; Weidlich, L.; Hoehne, L.; Flach, A.; Mendonc, L.A.; da Costa, A.; Ethur, E.M. Antimicrobial activities of six essential oils commonly used as condiments in Brazil against Clostridium perfringens. Braz. J. Microbiol. 2016, 47, 424–430. [Google Scholar] [CrossRef]
- Riahi, L.; Ghazghazi, H.; Ayari, B.; Aouadhi, C.; Klay, I.; Chograni, H.; Cherif, A.; Zoghlami, N. Effect of environmental conditions on chemical polymorphism and biological activities among Artemisia absinthium L. essential oil provenances grown in Tunisia. Ind. Crops Prod. 2015, 66, 96–102. [Google Scholar] [CrossRef]
- Paudel, P.N.; Satyal, P.; Satyal, R.; Setzer, W.N.; Gyawali, R. Chemical composition, enantiomeric distribution, antimicrobial and antioxidant activities of Origanum majorana L. essential oil from Nepal. Molecules 2022, 27, 6136. [Google Scholar] [CrossRef]
- Semenzato, G.; Del Duca, S.; Vassallo, A.; Zaccaroni, M.; Mucci, N.; Greco, C.; Padula, A.; Castronovo, L.M.; Chioccioli, S.; Pistelli, L.; et al. Exploring the nexus between the composition of essential oil and the bacterial phytobiome associated with different compartments of the medicinal plants Origanum vulgare ssp. vulgare, O. vulgare ssp. hirtum, and O. heracleot. Ind. Crops Prod. 2023, 191, 115997. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Ishangulyyeva, G.; Erbilgin, N.; Bonello, P. Terpenoids are involved in the expression of systemic-induced resistance in Austrian pine. Plant Cell Environ. 2024, 47, 2206–2227. [Google Scholar] [CrossRef]
- Rahmani, R.; Hedenström, E.; Schroeder, M. SPME collection and GC-MS analysis of volatiles emitted during the attack of male Polygraphus poligraphus (Coleoptera, Curcolionidae) on Norway spruce. Z. Für Naturforschung C 2015, 70, 265–273. [Google Scholar] [CrossRef]
- Toffolatti, L.; Maddalena, G.; Passera, A.; Casati, P.; Bianco, P.A.; Quaglino, F. 16-Role of terpenes in plant defense to biotic stress. In Biocontrol Agents and Secondary Metabolites; Applications and Immunization for Plant Growth and Protection; Woodhead publishing: Sawston, UK, 2021; pp. 401–417. [Google Scholar] [CrossRef]
- Zhao, T.; Krokene, P.; Björklund, N.; Långström, B.O.; Solheim, H.; Christiansen, E.; Borg-Karlson, A.-K. The influence of Ceratocystis polonica inoculation and methyl jasmonate application on terpene chemistry of Norway spruce, Picea abies. Phytochemistry 2010, 71, 1332–1341. [Google Scholar] [CrossRef]
- Wadke, N.; Kandasamy, D.; Vogel, H.; Lah, L.; Wingfield, B.D.; Paetz, C.; Wright, L.P.; Gershenzon, J.; Hammerbacher, A. The bark-beetle-associated fungus, Endoconidiophora polonica, utilizes the phenolic defence compounds of its host as a carbon source. Plant Physiol. 2016, 171, 914–931. [Google Scholar] [CrossRef] [PubMed]
- Hammerbacher, A.; Schmidt, A.; Wadke, N.; Wright, L.P.; Schneider, B.; Joerg Bohlmann, J.; Brand, W.A.; Fenning, T.M.; Gershenzon, J.; Paetz, C. A common fungal associate of the spruce bark beetle metabolizes the stilbene defenses of Norway spruce. Plant Physiol. 2013, 162, 1324–1336. [Google Scholar] [CrossRef]
- Zhao, T.; Kandasamy, D.; Krokene, P.; Chen, J.; Gershenzon, J.; Hammerbacher, A. Fungal associates of the tree-killing bark beetle, Ips typographus, vary in virulence, ability to degrade conifer phenolics and influence bark beetle tunneling behavior. Fungal Ecol. 2019, 38, 71–79. [Google Scholar] [CrossRef]
- Kandasamy, D.; Zaman, R.; Nakamura, Y.; Zhao, T.; Hartmann, H.; Andersson, M.N.; Hammerbacher, A.; Gershenzon, J. Conifer-killing bark beetles locate fungal symbionts by detecting volatile fungal metabolites of host tree resin monoterpenes. PLoS Biol. 2023, 21, e3001887. [Google Scholar] [CrossRef]
- Lehmanski, L.M.A.; Kandasamy, D.; Andersson, M.N.; Netherer, S.; Alves, E.G.; Huang, J.; Hartmann, H. Addressing a century-old hypothesis–do pioneer beetles of Ips typographus use volatile cues to find suitable host trees? New Phytol. 2023, 238, 1762–1770. [Google Scholar] [CrossRef]
- Blažytė-Čereškienė, L.; Apšegaitė, V.; Radžiutė, S.; Mozūraitis, R.; Būda, V.; Pečiulytė, D. Electrophysiological and behavioural responses of Ips typographus (L.) to trans-4-thujanol—A host tree volatile compound. Ann. For. Sci. 2016, 73, 247–256. [Google Scholar] [CrossRef]
- Schiebe, C.; Unelius, C.R.; Ganji, S.; Binyameen, M.; Birgersson, G.; Schlyter, F. Styrene, (+)-trans-(1R,4S,5S)-4-thujanol and oxygenated monoterpenes related to host stress elicit strong electrophysiological responses in the bark beetle Ips typographus. J. Chem. Ecol. 2019, 45, 474–489. [Google Scholar] [CrossRef] [PubMed]
- Jirošová, A.; Kalinová, B.; Modlinger, R.; Jakuš, R.; Unelius, C.R.; Blaženec, M.; Schlyter, F. Anti-attractant activity of (+)-trans-4-thujanol for Eurasian spruce bark beetle typographus: Novel potency for females. Pest Manag. Sci. 2022, 78, 1992–1999. [Google Scholar] [CrossRef] [PubMed]
- Trubin, A.; Mezei, P.; Afratakhti, K.Z.; Surovy, P.; Jakuš, R. Northernmost European spruce bark beetle Ips typographus outbreak: Modelling tree mortality using remote sensing and climate data. For. Ecol. Manag. 2022, 505, 119829. [Google Scholar] [CrossRef]
- Baños-Quintana, A.P.; Gershenzon, J.; Kaltenpoth, M. The Eurasian spruce bark beetle Ips typographus shapes the microbial communities of its offspring and the gallery environment. Front. Microbiol. 2024, 15, 1367127. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Wingfield, M.J.; Takahashi, I.; Solheim, H. Ophiostomatoid fungi associated with the spruce bark beetle Ips typographus f. aponicus in Japan. Mycol. Res. 1997, 1001, 1215–1227. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Takahashi, I.; Iguchi, K. Virulence of ophiostomatoid fungi associated with the spruce bark beetle Ips typographus f. japonicus in Yezo spruce. J. For. Res. 2000, 5, 87–94. [Google Scholar] [CrossRef]
- Kirisits, T. Fungal associates of European bark beetles with special emphasis on the opiostomatoid fungi. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Gregoire, J.-C., Evans, H.F., Eds.; Kluwer, Acad: Dordrecht, The Netherlands, 2004; pp. 181–236. [Google Scholar] [CrossRef]
- Bidart-Bouzat, M.G.; Imeh-Nathaniel, A. Global change effects on plant chemical dfenses against insect herbivores. J. Integr. Plant Biol. 2008, 50, 1339–1354. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Krokene, P. Chapter 5—Conifer defense and resistance to bark beetles. In Bark Beetles; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2015; pp. 177–207. [Google Scholar] [CrossRef]
- Fang, J.X.; Zhang, S.F.; Liu, F.; Zhang, X.; Zhang, F.B.; Guo, X.B.; Zhang, Z.; Zhang, Q.H.; Kong, X.B. Differences in gut bacterial communities of Ips typographus (Coleoptera: Curculionidae) induced by enantiomer-specific alpha-pinene. Environ. Entomol. 2020, 49, 1198–1205. [Google Scholar] [CrossRef]
- Chakraborty, A.; Purohit, A.; Khara, A.; Modlinger, R.; Roy, A. Life-stage and geographic location determine the microbial assemblage in Eurasian spruce bark beetle, Ips typographus L. (Coleoptera: Curculionidae). Front. Forest. Glob. Chang. 2023, 6, 1176160. [Google Scholar] [CrossRef]
- Moussa, A.; Nones, S.; Vannucchi, P.E.; Shahzad, G.I.R.; Dittmer, J.; Corretto, E.; Schebeck, M.; Faccoli, M.; Battisti, A.; Stauffer, C.; et al. The bacterial community of the European spruce bark beetle in space and time. bioRxiv 2023. [Google Scholar] [CrossRef]
- Veselská, T.; Švec, K.; Kostovčík, M.; Peral-Aranega, E.; Garcia-Fraile, P.; Křížková, B.; Havlíček, V.; Saati-Santamaría, Z.; Kolařík, M. Proportions of taxa belonging to the gut core microbiome change throughout the life cycle and season of the bark beetle Ips typographus. FEMS Microbiol. Ecol. 2023, 99, fiad072. [Google Scholar] [CrossRef] [PubMed]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus–a review of recent research. For. Ecol. Manag. 2004, 2002, 67–82. [Google Scholar] [CrossRef]
- Netherer, S.; Kandasamy, D.; Jirosová, A.; Kalinová, B.; Schebeck, M.; Schlyter, F. Interactions among Norway spruce, the bark beetle Ips typographus and its fungal symbionts in times of drought. J. Pest Sci. 2021, 94, 591–614. [Google Scholar] [CrossRef]
- Tanin, S.M.; Kandasamy, D.; Krokene, P. Fungal interactions and host tree preferences in the spruce bark beetle Ips typographus. Front. Microbiol. 2021, 12, 695167. [Google Scholar] [CrossRef]
- Johns, L.E.; Goldman, G.H.; Ries, L.N.A.; Brown, N.A. Nutrient sensing and acquisition in fungi: Mechanisms promoting pathogenesis in plant and human hosts. Fung. Biol. Rev. 2021, 36, 1–14. [Google Scholar] [CrossRef]
- Fei, W.; Liu, Y. Biotrophic fungal pathogens: A critical overview. Appl. Biochem. Biotechnol. 2023, 195, 1–16. [Google Scholar] [CrossRef]
- Liao, C.-J.; Hailemariam, S.; Sharon, A.; Mengiste, T. Pathogenic strategies and immune mechanisms to necrotrophs: Differences and similarities to biotrophs and hemibiotrophs. Curr. Opin. Plant Biol. 2022, 69, 102291. [Google Scholar] [CrossRef]
- Zeneli, G.; Krokene, P.; Chistiansen, E.; Krekling, T.; Gershenzon, J. Methyl jasmonate treatment of mature Norway spruce (Picea abies) trees increases the accumulation of terpenoid resin components and protects against infection by Ceratocystis polonica, a bark beetle-associated fungus. Tree Physiol. 2006, 26, 977–988. [Google Scholar] [CrossRef]
- Novak, M.; Urbanek Krajnc, A.; Lah, L.; Zupanec, N.; Kraševec, N.; Križman, M.; Bohlmann, J.; Komel, R. Low-density Ceratocystis polonica inoculation of Norway spruce (Picea abies) triggers accumulation of monoterpenes with antifungal properties. Eur. J. Forest Res. 2014, 133, 573–583. [Google Scholar] [CrossRef]
- Förster-Fromme, K.; Höschle, B.; Mack, C.; Bott, M.; Armbruster, W.; Jendrossek, D. Identification of genes and proteins necessary for catabolism of acyclic terpenes and leucine/isovalerate in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2006, 72, 4819–4828. [Google Scholar] [CrossRef] [PubMed]
- Van, L.T.; Hagiu, I.; Popovici, A.; Marinescu, F.; Gheorghe, I.; Curutiu, C.; Ditu, L.M.; Holban, A.-M.; Sesan, T.E.; Lazar, V. Antimicrobial efficiency of some essential oils in antibiotic-resistant Pseudomonas aeruginosa isolates. Plants 2022, 11, 2003. [Google Scholar] [CrossRef]
- Iseppi, R.; Mariani, M.; Condò, C.; Sabia, C.; Messi, P. Essential Oils: A natural weapon against antibiotic-resistant bacteria responsible for nosocomial infections. Antibiotics 2021, 10, 417. [Google Scholar] [CrossRef]
- Deka, B.; Baruah, C.; Babu, A. Entomopathogenic microorganisms: Their role in insect pest management. Egypt. J. Biol. Pest Control 2021, 31, 121. [Google Scholar] [CrossRef]
- Altinok, H.H.; Altinok, M.A.; Koca, A.S. Modes of action of entomopathogenic fungi. Curr. Trends Nat. Sci. 2019, 8, 117–124. Available online: http://natsci.upit.ro (accessed on 18 June 2024).
- Bugti, G.A.; Bin, W.; Memon, S.A.; Khaliq, G.; Jaffar, M.A. Entomopathogenic fungi: Factors involved in successful microbial control of insect pests. J. Entomol. 2020, 17, 74–83. Available online: https://scialert.net/abstract/?doi=je.2020.74.83 (accessed on 8 July 2024). [CrossRef]
- Qu, S.; Wang, S. Interaction of entomopathogenic fungi with the host immune system. Dev. Comp. Immunol. 2018, 83, 96–103. [Google Scholar] [CrossRef]
- Davis, T.S.; Stewart, J.E.; Mann, A.; Bradley, C.; Hofstetter, R.W. Evidence for multiple ecological roles of Leptographium abietinum, a symbiotic fungus associated with the north American spruce beetle. Fungal Ecol. 2019, 38, 62–70. [Google Scholar] [CrossRef]
- Elliot, S.L.; Sabelis, M.W.; Janssen, A.; Van Der Geest, L.P.S.; Beerling, E.A.M.; Fransen, J. Can plants use entomopathogens as bodyguards? Ecol. Lett. 2000, 3, 228–235. [Google Scholar] [CrossRef]
- Rostás, M.; Simon, M.; Hilker, M. Ecological cross-effects of induced plant responses towards herbivores and phytopathogenic fungi. Basic Appl. Ecol. 2003, 4, 43–62. [Google Scholar] [CrossRef]
- Zgoda, J.P.; Porter, J.R. A convenient microdilution method for screening natural products against bacteria and fungi. Pharm. Biol. 2001, 39, 221–225. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Samples | ||||||
|---|---|---|---|---|---|---|---|
| Sabinene Hydrate, mg/mL (μg/disc) | ETA 1 | Reference 2 | |||||
| 12.5 (125) | 25.0 (250) | 50.0 (500) | 75.0 (750) | 100 (1000) | |||
| Mycelial fungi | |||||||
| Ceratocystis polonica | 0.0 3 | 0.0 a | 6.0 ± 0.2 a | 13.4 ± 0.5 b, c | 15.5 ± 0.5 b | 11.5 ± 1.5 c | 30.0 ± 3.4 |
| Ophiostoma bicolor | 7.2 ± 0.3 a, b, e | 7.6 ± 0.6 a, f | 9.2 ± 0.8 b, c, d, g | 12.3 ± 2.1 c | 17.2 ± 3.8 d | 9.0 ± 0.9 e, f, g | 41.9 ± 2.7 |
| Ophiostoma penicillatum 4 | 0.0 a | 0.0 a | 6.0 ± 0.1 | 7.8 ± 0.6 b | 13.1 ± 0.7 | 10.5 ± 1.5 b | 30.5 ± 3.1 |
| Musicillium theobromae | 0.0 | 6.0 ± 0.1 a, b, d | 6.0 ± 0.1 a, c, e | 6.0 ± 0.3 b, c, f | 15.4 ± 0.2 | 6.8 ± 1.3 d, e, f | 36.8 ± 2.9 |
| Plectosphaerella cucumerina | 0.0 a | 0.0 a | 6.0 ± 0.0 | 7.4 ± 0.5 b | 11.8 ± 0.6 c | 7.0 ± 0.3 b | 12.6 ± 1.8 c |
| Aspergillus niger | 0.0 a, b | 0.0 a, c | 0.0 ± 0.0 b, c | 6.0 ± 0.1 d, e | 6.0 ± 0.0 d, f | 6.0 ± 0.0 e, f | 30.0 ± 1.0 |
| Penicillium notatum | 0.0 a | 0.0 a | 6.0 ± 0.0 b, c | 6.0 ± 0.0 b, d | 7.9 ± 0.6 | 6.0 ± 0.2 c, d | 28.2 ± 2.3 |
| Trichoderma sp. | 0.0 a | 0.0 a | 6.0 ± 0.1 b, c, e | 6.0 ± 0.0 b, d, f | 6.0 ± 0.2 c, d, g | 6.5 ± 0.3 e, f, g | 30.3 ± 3.5 |
| Yeasts | |||||||
| Candida albicans | 6.0 ± 0.1 a | 8.2 ± 0.3 a | 11.7 ± 1.5 | 16.1 ± 0.4 b | 21.4 ± 0.8 c | 10.2 ± 0.9 | 26.4 ± 6.1 b, c |
| Candida krueii | 6.0 ± 0.2 a, b, e | 6.0 ± 0.1 a, c, f | 8.5 ± 1.8 b, d, g | 8.6 ± 0.9 d, h | 15.5 ± 0.7 | 7.1 ± 0.8 e, f, g, h | 29.0 ± 2.3 |
| Candida parapsilosis | 6.0 ± 0.1 a, b, c, d | 6.0 ± 0.2 a, e | 6.0 ± 0.3 b, c, d, f | 7.0 ± 0.0 d | 18.4 ± 0.9 g | 6.6 ± 0.0 d, e, f | 16.8 ± 1.8 g |
| Saccharomyces cerevisiae 5 | 6.0 ± 0.2 a, d | 6.0 ± 0.1 a, b, c, e | 6.0 ± 0.1 b, c, f | 8.1 ± 0.3 | 18.3 ± 0.6 | 6.5 ± 0.2 d, e, f | 36.9 ± 4.2 |
| Rhodotorula muscilaginae 4 | 6.0 ± 0.1 a, d | 6.0 ± 0.1 a, b, e | 7.0 ± 0.2 b, c, f | 9.2 ± 1.3 c | 14.4 ± 0.6 | 8.2 ± 0.4 d, e, f | 33.7 ± 3.2 |
| Bacteria | |||||||
| Bacillus subtilis | 7.0 ± 0.4 a, b | 7.9 ± 0.8 a | 14.8 ± 2.3 c | 18.9 ± 2.1 d | 33.5 ± 2.7 e | 6.0 ± 0.3 b | 22.4 ± 6.1 c, d, e |
| Staphylococcus aureus | 6.0 ± 0.1 b | 8.0 ± 0.3 a | 9.1 ± 0.7 a | 13.4 ± 0.4 | 28.8 ± 1.6 c | 6.0 ± 0.2 b | 36.5 ± 4.8 c |
| Escherichia coli | 6.0 ± 0.0 a | 7.8 ± 0.5 a, b, d, e, f | 7.6 ± 0.2 c | 8.4 ± 0.9 b, c | 23.8 ± 1.2 | 7.2 ± 0.4 d, e, f | 31.7 ± 1.6 |
| Pseudomonas aeruginosa | 6.0 ± 0.1 a, b, d, k | 6.0 ± 0.1 a, e, f, g, l | 6.0 ± 0.2 b, e, h, i, m | 6.0 ± 0.3 c, f, h, j | 6.0 ± 0.4 d, g, i, j, n | 7.5 ± 0.7 k, l, m, n | 45.7 ± 8.6 |
| Organisms | Sabinene Hydrate, mg/mL (μg/mL) | Nystatin-Dihydrate, μg/mL | Chloramphenicol, μg/mL |
|---|---|---|---|
| Yeasts | |||
| Candida albicans | 0.125 (125) | 9.375 | – |
| Candida kruei | 0.25 (250) | 37.5 | – |
| Candida parapsilosis | 0.75 (750) | 150.0 | – |
| Saccharomyces cerevisiae | 0.5 (500) | 18.75 | – |
| Rhodotorula muscilaginosa | 0.5 (500) | 9.375 | – |
| Gram-positive bacteria | |||
| Bacillus subtilis | 0.0312 (30) | – | 62.50 |
| Staphylococcus aureus | 0.0625 (62.5) | – | <15.625 |
| Gram-negative bacteria | |||
| Escherichia coli | 0.125 (125) | – | 31.25 |
| Pseudomonas aeruginosa | >1.0 mg/mL * | – | 62.50 |
| Fungi | Sabinene Hydrate Concentration, mg/mL (μg/mL) | |||||
|---|---|---|---|---|---|---|
| 0.0625 (62.5) | 0.125 (125) | 0.25 (2500) | 0.5 (500) | 0.75 (750) | 1.0 (1000) | |
| Ceratocystis polonica | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 8.62 ± 4.80 b | 17.11 ± 4.35 b, c | 31.19 ± 5.17 | 29.64 ± 7.07 c |
| Ophiostoma bicolor | 0.52 ± 0.21 a | 1.59 ± 1.43 a | 21.21 ± 9.01 | 47.98 ± 10.09 b, c, d | 59.93 ± 19.08 b, c, d | 67.35 ± 9.93 d |
| O. penicillatum | 0.0 ± 0.0 a | 0.0 ± 0.01 a | 18.93 ± 7.17 | 32.14 ± 5.74 | 49.46 ± 12.38 b | 49.21 ± 3.67 b |
| Plectosphaerella cucumerina | 0.0 ± 0.0 a, b | 0.0 ± 0.01 a | 0.0 ± 0.01 b | 7.35 ± 2.51 c | 7.68 ± 5.22 c | 19.89 ± 6.47 |
| Aspergillus niger | 0.0 ± 0.0 a, b | 0.0 ± 0.00 a, c | 0.0 ± 0.01 b | 0.0 ± 0.00 c | 0.82 ± 0.31 d | 1.45 ± 0.82 d |
| Musicillium theobromae | 0.0 ± 0.0 a | 0.0 ± 0.00 a | 6.73 ± 2.98 b | 13.25 ± 4.91 c, d | 8.97 ± 6.14 b, c | 24.00 ± 9.45 d |
| Nystatin-Dihydrate (N-D) Concentration, mg/mL (μg/mL) | ||||||
| 0.009375 (9.375) | 0.01875 (18.75) | 0.0375 (37.5) | 0.075 (75) | 0.15 (150) | 0.3 (300) | |
| C. polonica | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 26.71 ± 11.14 | 63.28 ± 24.41 | 98.55 ± 1.23 a | 100 a |
| O. bicolor | 0.92 ± 0.55 | 13.07 ± 4.46 | 48.92 ± 20.02 | 80.27 ± 19.22 a | 100 a, b | 100 b |
| O. penicillatum | 0.08 ± 0.03 | 0.88 ± 0.22 | 37.52 ± 11.55 | 96.27 ± 3.71 a, b | 100 a, b, c | 100 c |
| P. cucumerina | 0.19 ± 0.08 | 14.38 ± 9.71 a | 33.61 ± 22.04 a | 87.23 ± 10.71 b | 99.01 ± 0.04 b, c | 100 c |
| A. niger | 0.04 ± 0.01 | 3.56 ± 1.88 | 22.83 ± 6.47 | 86.87 ± 5.23 a | 86.17 ± 13.64 a, b | 100 b |
| M. theobromae | 0.62 ± 0.17 | 31.05 ± 27.91 a | 66.37 ± 33.06 a, b, c, d | 90.87 ± 6.61 b, c, d | 96.14 ± 2.87 b, c, d | 100 b, c, d |
| Inhibiting Fungus | Medium | Inhibition of Radial Growth Rate (%) | ||
|---|---|---|---|---|
| C. polonica | O. bicolor | O. penicillatum | ||
| Ceratocystis polonica | MEA | 12.08 ± 1.88 | 78.91 ± 11.47 | |
| MEA + SPE | 49.31 ± 10.31 a | 46.77 ± 20.28 a | ||
| Oprhiostoma bicolor | MEA | 98.89 ± 1.11 | 43.28 ± 10.99 | |
| MEA + SPE | 24.68 ± 9.55 a | 41.63 ± 4.75 a | ||
| Ophiostoma penicillatum | MEA | 12.43 ± 4.88 | 65.14 ± 22.07 | |
| MEA + SPE | 49.27 ± 10.87 a | 66.89 ± 15.68 a | ||
| Beauveria bassiana | MEA | 50.77 ± 12.32 | 22.16 ± 3.84 | 94.11 ± 5.33 |
| MEA + SPE | 45.54 ± 6.27 a | 24.02 ± 8.33 a | 91.58 ± 8.03 | |
| Lecanicillium fungicola | MEA | 82.42 ± 9.45 a | 38.34 ± 11.66 | 100 a |
| MEA + SPE | 86.82 ± 13.15 a | 56.72 ± 15.21 | 100 a | |
| Metarhizium anissopliae | MEA | 58.62 ± 15.27 a | 17.89 ± 5.16 | 62.19 ± 13.04 a |
| MEA + SPE | 57.44 ± 20.33 a | 21.14 ± 8.87 | 60.73 ± 21.04 a | |
| Musicillium theobromae | MEA | 59.46 ± 10.19 a, b | 78.45 ± 14.37 a, c | 46.28 ± 11.17 b, c |
| MEA + SPE | 32.29 ± 14.03 a, b | 53.67 ± 22.02 a, c | 36.66 ± 6.04 b, c | |
| Plectosphaerella cucumerina | MEA | 46.55 ± 5.09 | 14.56 ± 4.08 | 90.01 ± 4.17 |
| MEA + SPE | 44.10 ± 16.99 a | 18.23 ± 2.78 a | 89.65 ± 10.20 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Judžentienė, A.; Pečiulytė, D.; Nedveckytė, I. In Situ Antimicrobial Properties of Sabinene Hydrate, a Secondary Plant Metabolite. Molecules 2024, 29, 4252. https://doi.org/10.3390/molecules29174252
Judžentienė A, Pečiulytė D, Nedveckytė I. In Situ Antimicrobial Properties of Sabinene Hydrate, a Secondary Plant Metabolite. Molecules. 2024; 29(17):4252. https://doi.org/10.3390/molecules29174252
Chicago/Turabian StyleJudžentienė, Asta, Dalė Pečiulytė, and Irena Nedveckytė. 2024. "In Situ Antimicrobial Properties of Sabinene Hydrate, a Secondary Plant Metabolite" Molecules 29, no. 17: 4252. https://doi.org/10.3390/molecules29174252