
Neurotoxicity, Neuroprotection, In Vitro MAOA/MAOB Inhibitory Activity Assessment, Molecular Docking, and Permeability Assay Studies of Newly Synthesized Hydrazones Containing a Pyrrole Ring

 ,
,  , , , ,
, , , ,
Abstract
1. Introduction
2. Results
2.1. Chemistry
2.2. In Vitro Neurotoxicity Evaluation
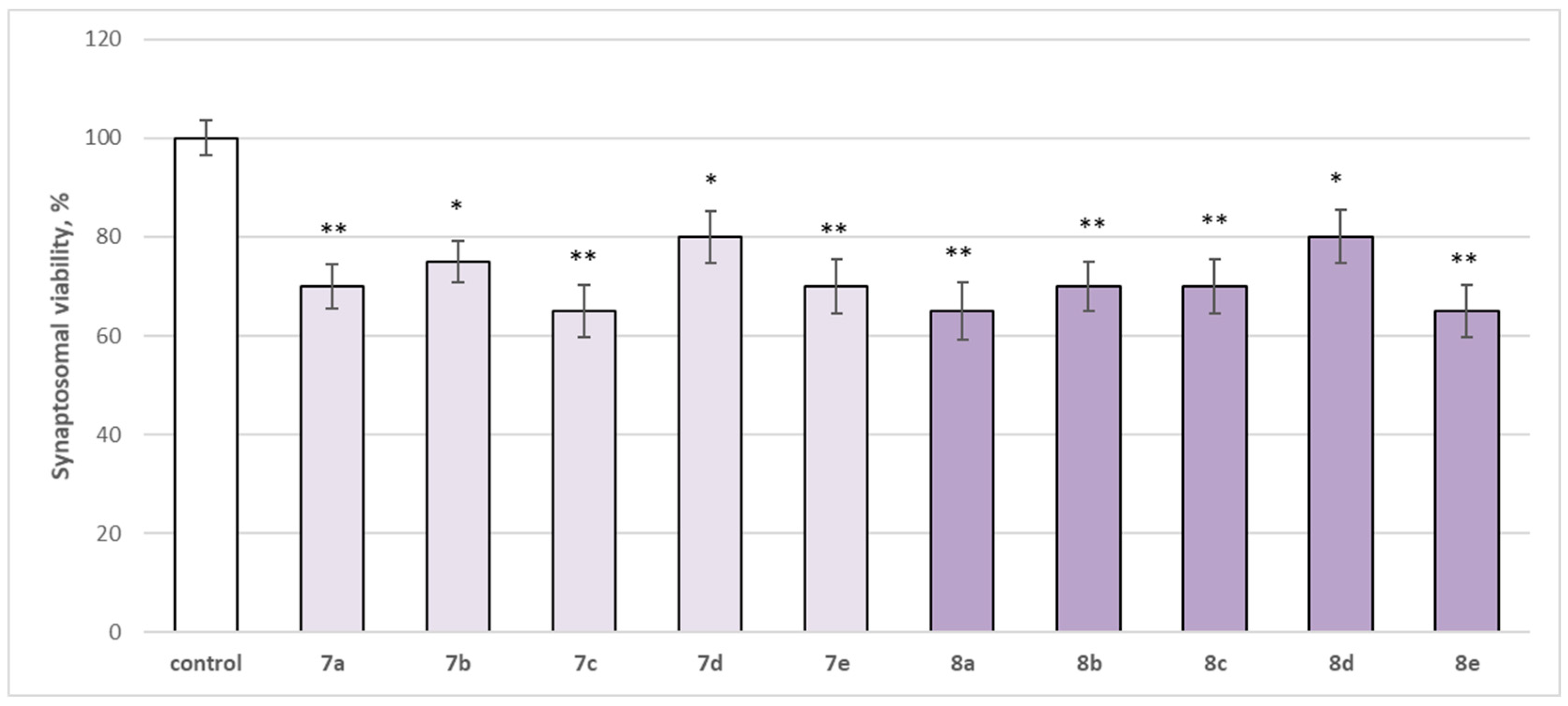
2.2.1. Effects of Newly Synthesized Hydrazones on Isolated Rat Brain Synaptosomes
2.2.2. Effects of Newly Synthesized Hydrazones on Isolated Rat Brain Mitochondria
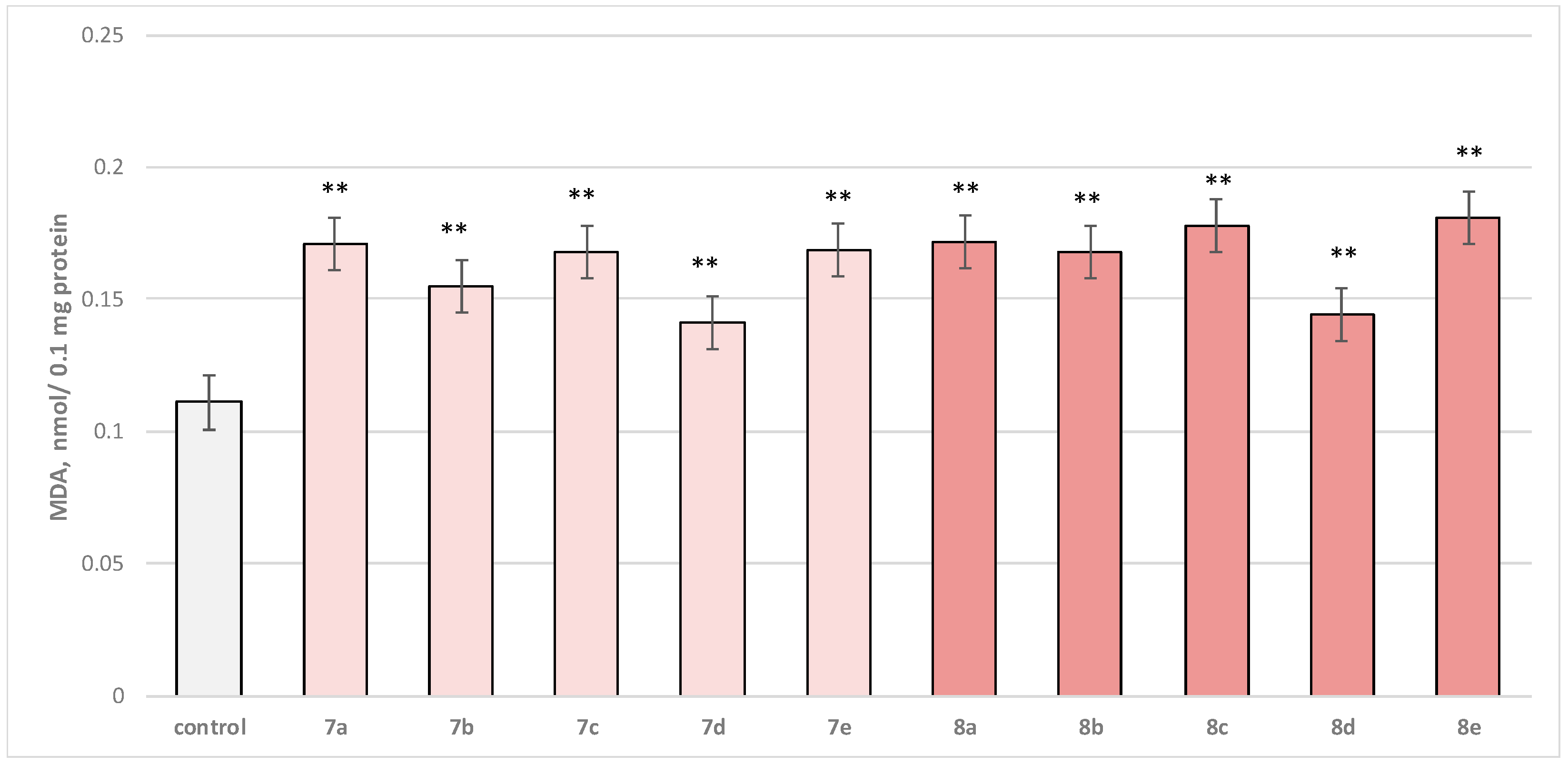
2.2.3. Effects of Newly Synthesized Hydrazones on Isolated Rat Brain Microsomes
2.3. In Vitro Evaluation of Neuroprotective Effects of the Target Compounds
2.3.1. Effect of the Target Substances in a Model of 6-OHDA-Induced Neurotoxicity on Isolated Rat Brain Synaptosomes
2.3.2. Effect of the Target Substances in a Model of t-BuOOH-Induced Oxidative Stress on Isolated Rat Brain Mitochondria
2.3.3. Effect of the Target Substances in a Model of Fe2+/AA-Induced Oxidative Stress on Isolated Rat Brain Microsomes
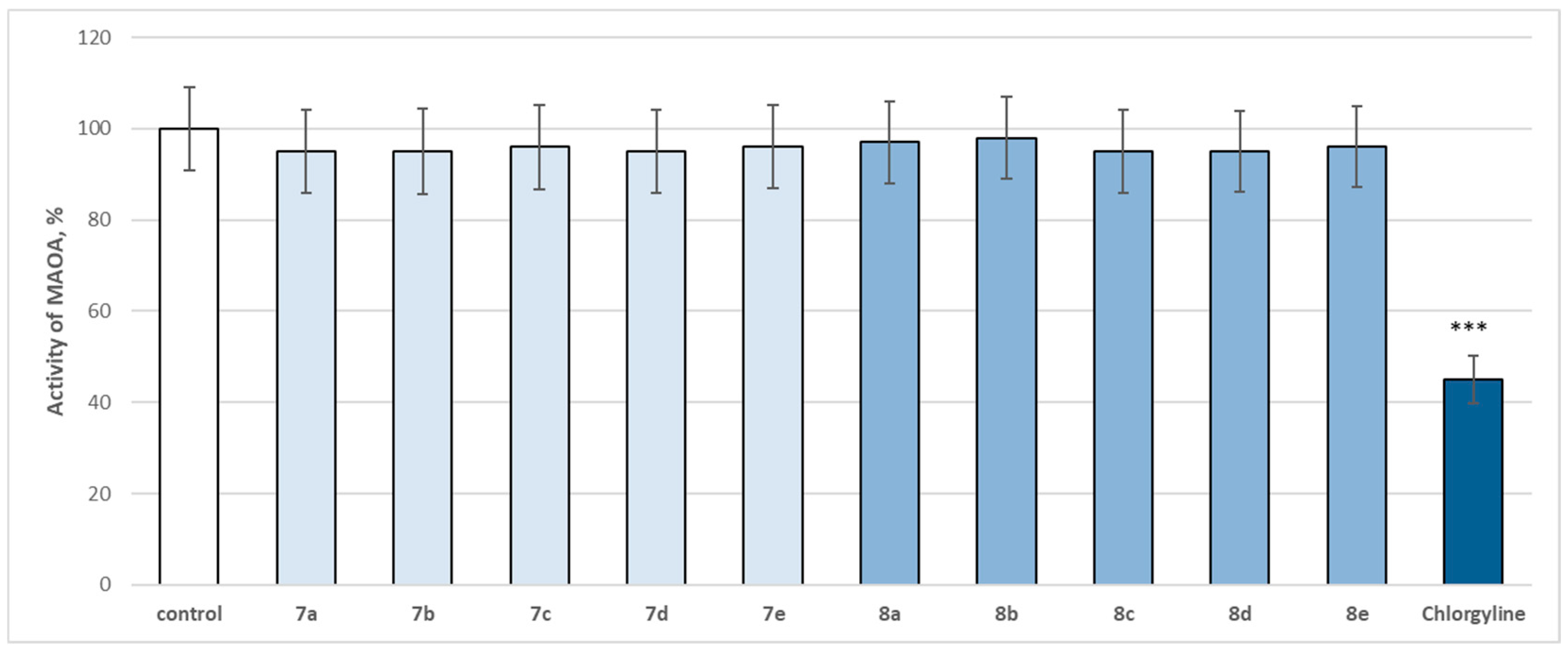
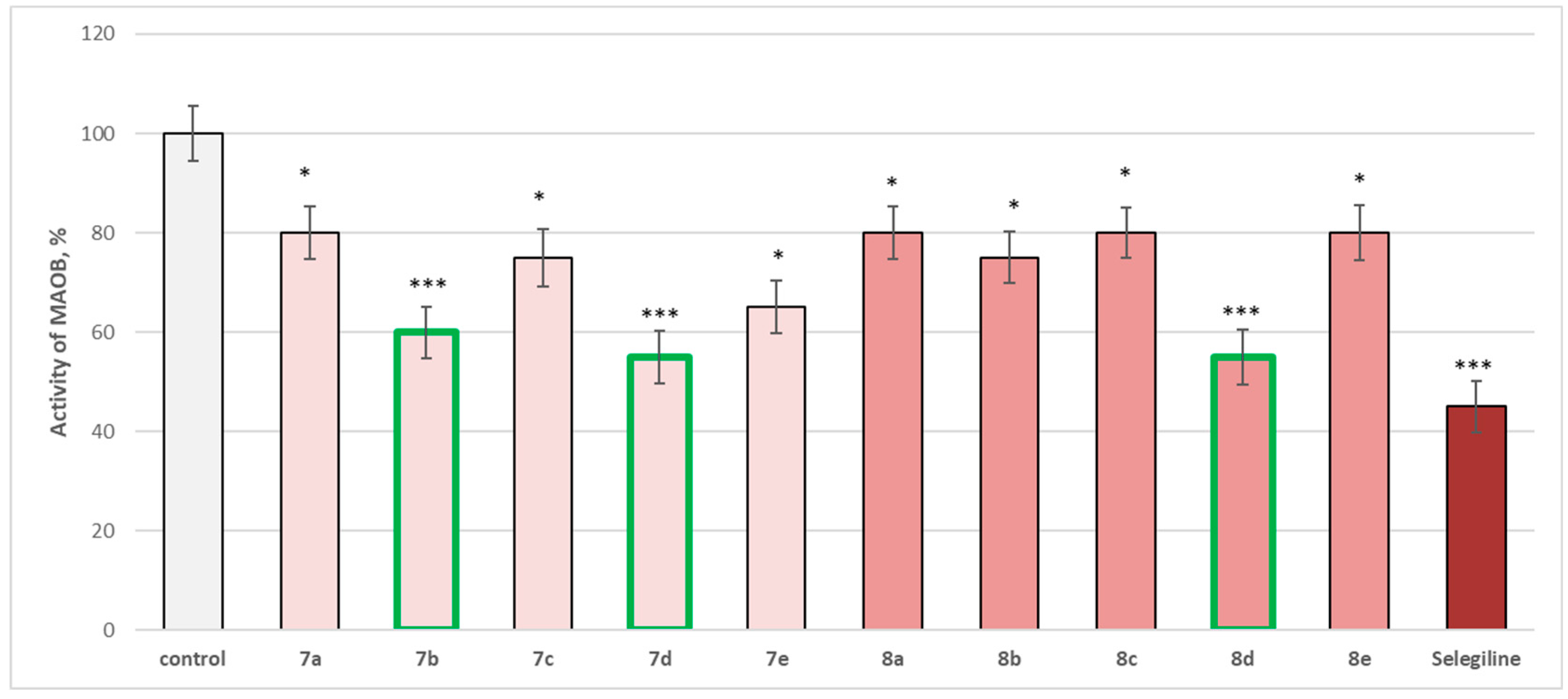
2.4. In Vitro Evaluation of the Enzymatic Activity of the Target Molecules on Human Recombinant MAOA/MAOB Enzyme
2.5. Molecular Docking
2.5.1. Molecular Docking in the Active Site of MAOA
2.5.2. Molecular Docking in the Active Site of MAOB
2.6. Estimation of BBB Permeability by PAMPA
3. Discussion
3.1. In Vitro Subcellular Evaluation of the Neurotoxicity Effect of Substances Administered Alone on Biomarkers, Characterizing the Functional-Metabolic Profile of Brain Synaptosomes, Mitochondria, and Microsomes
3.2. In Vitro Subcellular Evaluation of the Neuroprotective Effect of Target Substances Administered Alone in Different Models of Induced Cellular Toxicity
3.2.1. Effect of Substances in a Model of 6-OHDA-Induced Neurotoxicity in Isolated Rat Brain Synaptosomes
3.2.2. Effect of Substances in a Model of t-BuOOH-Induced Oxidative Stress on Isolated Rat Brain Mitochondria
3.2.3. Effect of Substances in a Model of Fe2+/AA-Induced Oxidative Stress on Isolated Rat Brain Microsomes
3.3. In Vitro Evaluation of the Enzymatic Activity of the Target Molecules on Human Recombinant MAOA/MAOB Enzyme
3.4. Molecular Docking
3.5. Estimation of BBB Permeability by PAMPA
4. Materials and Methods
4.1. Chemistry
4.2. Pharmacological Studies
4.2.1. Chemicals and Reagents
4.2.2. Animals
4.2.3. Preparation of Rat Brain Synaptosomes and Mitochondria
4.2.4. Isolation of the Corresponding Synaptosomal and Mitochondrial Fractions
4.2.5. Incubation of Synaptosomes
4.2.6. Establishing and Applying a Dopamine Model of Neurotoxicity
4.2.7. MTT Assay to Assess Synaptosomal Viability
4.2.8. Determination of Reduced Glutathione (GSH) in Isolated Brain Synaptosomes
4.2.9. Tert-Butyl Hydroperoxide-Induced Oxidative Stress
4.2.10. Determination of Malondialdehyde (MDA) Production in Brain Mitochondria [58]
4.2.11. Determination of GSH Level in Brain Mitochondria [60]
4.2.12. Isolation of Brain Microsomes [61]
4.2.13. Iron/Ascorbate-Induced Lipid Peroxidation (LPO)
4.2.14. Determination of MDA in Brain Microsomes [62]
4.2.15. Determination of Human Recombinant MAOA/B Enzyme Activity
4.2.16. Statistical Methods
4.3. Molecular Docking
4.4. PAMPA-BBB Assay
4.5. Physicochemical Properties Evaluation of the Molecules
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schapira, A.H.V.; Chaudhuri, K.R.; Jenner, P. Non-motor features of Parkinson disease. Nat. Rev. Neur. 2017, 18, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Yankee, E.L. A review on Parkinson’s disease treatment. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, 222–244. [Google Scholar] [CrossRef]
- Rawat, C.S.; Pandey, S. Parkinson’s Disease—An Introduction. In Techniques for Assessment of Parkinsonism for Diagnosis and Rehabilitation; Series in BioEngineering; Arjunan, S.P., Kumar, D.K., Eds.; Springer: Singapore, 2022. [Google Scholar] [CrossRef]
- Sacristán, H.E.; Serra Fulles, J.A. The essentials in Parkinson’s disease. EC Neurol. 2023, 15, 36–67. [Google Scholar]
- Berardelli, A. Pathophysiology of bradykinesia in Parkinson’s disease. Brain 2001, 124, 2131–2146. [Google Scholar] [CrossRef] [PubMed]
- Bortolato, M.; Chen, K.; Shih, J.C. Monoamine oxidase inactivation: From pathophysiology to therapeutics. Adv. Drug Deliv. Rev. 2008, 60, 1527–1533. [Google Scholar] [CrossRef]
- Carradori, S.; Secci, D.; Petzer, J.P. MAO inhibitors and their wider applications: A patent review. Expert. Opin. Ther. Pat. 2018, 28, 211–226. [Google Scholar] [CrossRef]
- Stocchi, F.; Fossati, C.; Torti, M. Rasagiline for the treatment of Parkinson’s disease: An update. Expert. Opin. Pharmacother. 2015, 16, 2231–2241. [Google Scholar] [CrossRef]
- Chen, J.J.; Swope, D.M. Clinical pharmacology of rasagiline: A novel, second-generation propargylamine for the treatment of Parkinson disease. J. Clin. Pharmacol. 2005, 45, 878–894. [Google Scholar] [CrossRef]
- Bainbridge, J.L.; Page, R.L., 2nd; Ruscin, J.M. Elucidating the mechanism of action and potential interactions of MAO-B inhibitors. Neurol. Clin. 2008, 26, 85–96. [Google Scholar] [CrossRef]
- Tan, Y.-Y.; Jenner, P.; Chen, S.-D. Monoamine oxidase-B inhibitors for the treatment of Parkinson’s disease: Past, present, and future. J. Park. Dis. 2022, 12, 477–493. [Google Scholar] [CrossRef]
- Bhardwaj, V.; Gumber, D.; Abbot, V.; Dhiman, S.; Sharma, P. Pyrrole: A resourceful small molecule in key medicinal hetero-aromatics. RSC Adv. 2015, 5, 15233–15266. [Google Scholar] [CrossRef]
- Bavadi, M.; Niknam, K.; Shahraki, O. Novel pyrrole derivatives bearing sulfonamide groups: Synthesis in vitro cytotoxicity evaluation, molecular docking and DFT study. J. Mol. Struct. 2017, 1146, 242–253. [Google Scholar] [CrossRef]
- Girgis, A.S.; Stawinski, J.; Ismail, N.S.M.; Farag, H. Synthesis and QSAR study of novel cytotoxic spiro[3h-indole-3,2′(1′h)-pyrrolo[3,4-c]pyrrole]-2,3′,5′(1h,2′ah,4′h)-triones. Europ. J. Med. Chem. 2012, 47, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Nobuyoshi, A.; Okamoto, A.; Murakata, C.; Tamaoki, T.; Okabe, M.; Saito, H. Synthesis and antitumor activity of duocarmycin derivatives: modification of segment-a of a-ring pyrrole compounds. J. Med. Chem. 1999, 42, 2946–2960. [Google Scholar] [CrossRef]
- Fatahala, S.S.; Nofal, S.; Mahmoud, E.; El-hameed, R.H.A. Pyrrolopyrazoles: Synthesis, evaluation and pharmacological screening as antidepressant agents. Med. Chem. 2019, 15, 911–922. [Google Scholar] [CrossRef]
- Goel, A.; Agarwal, N.; Singh, F.V.; Sharon, A.; Tiwari, P.; Dixit, M.; Pratap, R.; Srivastava, A.K.; Maulik, P.R.; Ram, V.J. Antihyperglycemic activity of 2-methyl-3,4,5-triaryl-1 H-pyrroles in SLM and STZ models. Bioorg. Med. Chem. Lett. 2004, 14, 1089–1092. [Google Scholar] [CrossRef]
- Delia, D.; Lampis, G.; Fioravanti, R.; Biava, M.; Porretta, G.C.; Zanetti, S.; Pompei, R. Bactericidal activities of the pyrrole derivative BM212 against multidrug-resistant and intramacrophagic Mycobacterium tuberculosis strains. Antimicrob. Agents Chemother. 1998, 42, 3035–3037. [Google Scholar] [CrossRef]
- Biava, M.; Porretta, G.C.; Deidda, D.; Pompei, R.; Tafi, A.; Manetti, F. Antimycobacterial compounds. new pyrrole derivatives of BM212. Bioorg. Med. Chem. 2004, 12, 1453–1458. [Google Scholar] [CrossRef]
- Joshi, S.D.; Sheshagiri, R.D.; Kirankumar, M.N.; Aminabhavi, T.M.; Raju, K.V.S.N.; Narayan, R.; Lherbet, C.; Yang, K.S. Synthesis, antimycobacterial screening and ligand-based molecular docking studies on novel pyrrole derivatives bearing pyrazoline, isoxazole and phenyl thiourea moieties. Europ. J. Med. Chem. 2016, 107, 133–152. [Google Scholar] [CrossRef]
- Ragno, R.; Marshall, G.R.; Di Santo, R.; Costi, R.; Massa, S.; Rompei, R.; Artico, M. Antimycobacterial pyrroles: Synthesis, anti-Mycobacterium tuberculosis activity and QSAR studies. Bioorg. Med. Chem. 2000, 8, 1423–1432. [Google Scholar] [CrossRef]
- Georgieva, M. Pyrroles—A New Perspective Class Tuberculostatics, 1st ed.; Taurus Advertising EOOD: Sofia, Bulgaria, 2022. [Google Scholar]
- Bratton, L.D.; Auerbach, B.; Choi, C.; Dillon, L.; Hanselman, J.C.; Larsen, S.D.; Lu, G.; Olsen, K.; Pfefferkorn, J.A.; Robertson, A.; et al. Discovery of pyrrole-based hepatoselective ligands as potent inhibitors of HMG-CoA reductase. Bioorg. Med. Chem. 2007, 15, 5576–5589. [Google Scholar] [CrossRef] [PubMed]
- Tzankova, D.; Aluani, D.; Kondeva-Burdina, M.; Georgieva, M.; Vladimirova, S.; Peikova, L.; Tzankova, V. Antioxidant properties, neuroprotective effects and in vitro safety evaluation of new pyrrole derivatives. Pharm. Chem. J. 2022, 55, 1310–1319. [Google Scholar] [CrossRef]
- Kundu, T.; Pramanik, A. Expeditious and eco-friendly synthesis of new multifunctionalized pyrrole derivatives and evaluation of their antioxidant property. Bioorg. Chem. 2020, 98, 103734. [Google Scholar] [CrossRef]
- La Regina, G.; Silvestri, R.; Artico, M.; Lavecchia, A.; Novellino, E.; Befani, O.; Turini, P.; Agostinelli, E. New pyrrole inhibitors of monoamine oxidase: Synthesis, biological evaluation, and structural determinants of MAO-A and MAO-B selectivity. J. Med. Chem. 2007, 50, 922–931. [Google Scholar] [CrossRef]
- Altintop, M.D.; Sever, B.; Osmaniye, D.; Sağlık, B.N.; Özdemir, A. Design, synthesis, in vitro and in silico evaluation of new pyrrole derivatives as monoamine oxidase inhibitors. Arch. Pharm. 2018, 351, 1800082. [Google Scholar] [CrossRef]
- Kondeva-Burdina, M.; Mateev, E.; Angelov, B.; Tzankova, V.; Georgieva, M. In silico evaluation and in vitro determination of neuroprotective and MAO-B inhibitory effects of pyrrole-based hydrazones: A therapeutic approach to Parkinson’s disease. Molecules 2022, 27, 8485. [Google Scholar] [CrossRef]
- Tzankova, D.; Kuteva, H.; Mateev, E.; Stefanova, D.; Dzhemadan, A.; Yordanov, Y.; Mateeva, A.; Tzankova, V.; Kondeva-Burdina, M.; Zlatkov, A.; et al. Synthesis, DFT study, and in vitro evaluation of antioxidant properties and cytotoxic and cytoprotective effects of new hydrazones on SH-SY5Y neuroblastoma cell lines. Pharmaceuticals 2023, 16, 1198. [Google Scholar] [CrossRef]
- Rankovic, Z. CNS drug design: Balancing physicochemical properties for optimal brain exposure. J. Med. Chem. 2015, 58, 2584–2608. [Google Scholar] [CrossRef]
- Dias, V.; Junn, E.; Mouradian, M.M. The Role of Oxidative Stress in Parkinson’s Disease. J. Park. Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef]
- Gundersen, V. Protein Aggregation in Parkinson’s disease: Protein aggregation in Parkinson’s disease. Acta Neurol. Scand. 2010, 122, 82–87. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s disease and its potential as therapeutic target. Transl. Neurodegener. 2015, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Gerberick, G.F.; Vassallo, J.D.; Foertsch, L.M.; Price, B.B.; Chaney, J.G.; Lepoittevin, J.P. Quantification of chemical peptide reactivity for screening contact allergens: A classification tree lmodel approach. Toxicol. Sci. 2007, 97, 417–427. [Google Scholar] [CrossRef]
- Yarbrough, J.W.; Schultz, T.W. Abiotic sulfhydryl reactivity: A predictor of aquatic toxicity for carbonyl containing alpha, beta-unsaturated compounds. Chem. Res. Toxicol. 2007, 20, 558–562. [Google Scholar] [CrossRef]
- Rebecca, C.T.; Khan, A.; Uygur, B.; Saad, M.; Yeh, H.-W.; DelGaudio, N.; Weber, R.; Alwaseem, H.; Gao, J.; Yang, C.; et al. A mouse model to study glutathione limitation in vivo. bioRxiv 2024, 574722. [Google Scholar] [CrossRef]
- Bohme, A.; Thaens, D.; Paschke, A.; Schuurmann, G. Kinetic glutathione chemoassay to quantify thiol reactivity of organic electrophiles-application to alpha, beta-unsaturated ketones, acrylates, and propiolates. Chem. Res. Toxicol. 2009, 22, 742–750. [Google Scholar] [CrossRef]
- Zhang, W.; Gao, J.; Lu, L.; Bold, T.; Li, X.; Wang, S.; Chang, Z.; Chen, J.; Kong, X.; Zheng, Y.; et al. Intracellular GSH/GST antioxidants system change as an earlier biomarker for toxicity evaluation of iron oxide nanoparticles. NanoImpact 2021, 23, 100338. [Google Scholar] [CrossRef]
- Stokes, A.H.; Freeman, W.M.; Mitchell, S.G.; Burnette, T.A.; Hellmann, G.M.; Vrana, K.E. Induction of GADD45 and GADD153 in neuroblastoma cells by dopamine-induced toxicity. Neurotoxicology 2002, 23, 675–684. [Google Scholar] [CrossRef]
- Drahota, Z.; Krivakova, P.; Cervinkova, Z.; Kmonickova, E.; Lotkova, H.; Kucera, O.; Houstek, J. Tert-butyl hydroperoxide selectively inhibits mitochondrial respiratory-chain enzymes in isolated rat hepatocytes. Physiol. Res. 2005, 54, 67–72. [Google Scholar] [CrossRef]
- Öllinger, K.; Brunk, U.T. Cellular injury induced by oxidative stress is mediated through lysosomal damage. Free Radic. Biol. Med. 1995, 19, 565–574. [Google Scholar] [CrossRef]
- O’Donnell, V.; Burkitt, M.J. Mitochondrial metabolism of a hydroperoxide to free radicals in human endothelial cells: An electron spin resonance spin-trapping investigation. Biochem. J. 1994, 304, 707–713. [Google Scholar] [CrossRef]
- Zaidi, S.I.; Agarwal, R.; Eichler, G.; Rihter, B.D.; Kenney, M.E.; Mukhtar, H. Photodynamic effects of new silicon phthalocyanines: In vitro studies utilizing rat hepatic microsomes and human erythrocyte ghosts as model membrane sources. Photochem. Photobiol. 1993, 58, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Rathitharan, G.; Meyer, J.H.; Furukawa, Y.; Ang, L.C.; Boileau, I.; Guttman, M.; Hornykiewicz, O.; Kish, S.J. Brain monoamine oxidase B and A in human parkinsonian dopamine deficiency disorders. Brain 2017, 40, 2460–2474. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, S.; Hoda, N. A comprehensive review of monoamine oxidase inhibitors as Anti-Alzheimer’s disease agents: A review. Eur. J. Med. Chem. 2020, 206, 112787. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.-H.; Sa, M.; Ju, Y.H.; Park, M.G.; Lee, C.J. Revisiting the role of astrocytic MAOB in Parkinson’s disease. Int. J. Mol. Sci. 2022, 23, 4453. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.W.; Doba, T.; Gabe, E.J.; Hughes, L.; Lee, F.L.; Prasad, L.; Ingoldo, K.U. Autoxidation of biological molecules. 4. Maximizing the antioxidant activity of phenols. J. Am. Chem. Soc. 1985, 107, 7053–7065. [Google Scholar] [CrossRef]
- Lee, C.Y.; Sharma, A.; Semenya, J.; Anamoah, C.; Chapman, K.N.; Barone, V. Computational study of ortho-substituent effects on antioxidant activities of phenolic dendritic antioxidants. Antioxidants 2020, 9, 189. [Google Scholar] [CrossRef]
- Dasgupta, S.; Mukherjee, S.; Sekar, K.; Mukhopadhyay, B.P. The conformational dynamics of wing gates Ile199 and Phe103 on the binding of dopamine and benzylamine substrates in human monoamine oxidase B. J. Biomol. Struct. Dyn. 2021, 39, 1879–1886. [Google Scholar] [CrossRef]
- Van de Waterbeemd, H.; Camenisch, G.; Folkers, G.; Chretien, J.R.; Raevsky, O.A. Estimation of blood-brain barrier crossing of drugs using molecular size and shape, and H-bonding descriptors. J. Drug Target. 1998, 6, 151–165. [Google Scholar] [CrossRef]
- Ghose, A.K.; Herbertz, T.; Hudkins, R.L.; Dorsey, B.D.; Mallamo, J.P. Knowledge-based, central nervous system (CNS) lead selection and lead optimization for CNS drug discovery. ACS Chem. Neurosci. 2012, 3, 50–68. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A boiled-egg to predict gastrointestinal absorption and brain penetration of small molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef]
- Hitchcock, S.A.; Pennington, L.D. Structure-brain exposure relationships. J. Med. Chem. 2006, 49, 7559–7583. [Google Scholar] [CrossRef] [PubMed]
- M Reis, J.; Sinko, B.; HR Serra, C. Parallel artificial membrane permeability assay (PAMPA)-Is it better than Caco-2 for human passive permeability prediction? Mini Rev. Med. Chem. 2010, 10, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Didziapetris, R.; Japertas, P.; Avdeef, A.; Petrauskas, A. Classification analysis of P-glycoprotein substrate specificity. J. Drug Target. 2003, 11, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Taupin, P.; Zini, S.; Cesselin, F.; Ben-Ari, Y.; Roisin, M.P. Subcellular fractionation on Percoll gradient of mossy fiber synaptosomes: Morphological and biochemical characterization in control and degranulated rat hippocampus. J. Neurochem. 1994, 62, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Mungarro-Menchaca, X.; Ferrera, P.; Morán, J.; Arias, C. β-Amyloid peptide induces ultrastructural changes in synaptosomes and potentiates mitochondrial dysfunction in the presence of ryanodine. J. Neurosci. Res. 2002, 68, 89–96. [Google Scholar] [CrossRef]
- Robyt, J.F.; Ackerman, R.J.; Chittenden, C.G. Reaction of protein disulfide groups with Ellman’s reagent: A case study of the number of sulfhydryl and disulfide groups in Aspergillus oryzae α-amylase, papain, and lysozyme. Arch. Biochem. Biophys. 1971, 147, 262–269. [Google Scholar] [CrossRef]
- Karlsson, J.; Emgard, M.; Brundin, P.; Burkitt, M.J. trans-Resveratrol protects embryonic mesencephalic cells from tert-butyl hydroperoxide: Electron paramagnetic resonance spin trapping evidence for a radical scavenging mechanism. J. Neurochem. 2000, 75, 141–150. [Google Scholar] [CrossRef]
- Shirani, M.; Alizadeh, S.; Mahdavinia, M.; Dehghani, M.A. The ameliorative effect of quercetin on bisphenol A-induced toxicity in mitochondria isolated from rats. Environ. Sci. Pollut. Res. Int. 2019, 26, 7688–7696. [Google Scholar] [CrossRef]
- Ravindranath, V.; Anandatheerthavarada, H.K. Preparation of brain microsomes with cytochrome P450 activity using calcium aggregation method. Anal. Biochem. 1990, 187, 310–313. [Google Scholar] [CrossRef]
- Mansuy, D.; Sassi, A.; Dansette, P.M.; Plat, M. A new potent inhibitor of lipid peroxidation in vitro and in vivo, the hepatoprotective drug anisyldithiolthione. Biochem. Biophys. Res. Commun. 1986, 135, 1015–1021. [Google Scholar] [CrossRef]
- Bautista-Aguilera, O.M.; Esteban, G.; Bolea, I.; Nikolic, K.; Agbaba, D.; Moraleda, I.; Iriepa, I.; Samadi, A.; Soriano, E.; Unzeta, M.; et al. Design, synthesis, pharmacological evaluation, QSAR analysis, molecular modeling and ADMET of novel donepezil-indolyl hybrids as multipotent cholinesterase/monoamine oxidase inhibitors for the potential treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2014, 21, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Doytchinova, I.; Atanasova, M.; Valkova, I.; Stavrakov, G.; Philipova, I.; Zhivkova, Z.; Zheleva-Dimitrova, D.; Konstantinov, S.; Dimitrov, I. Novel hits for acetylcholinesterase inhibition derived by docking-based screening on ZINC database. J. Enzym. Inhib. Med. Chem. 2018, 33, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Di, L.; Kerns, E.H.; Fan, K.; McConnell, O.J.; Carter, G.T. High throughput artificial membrane permeability assay for blood-brain barrier. Eur. J. Med. Chem. 2003, 38, 223–232. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]




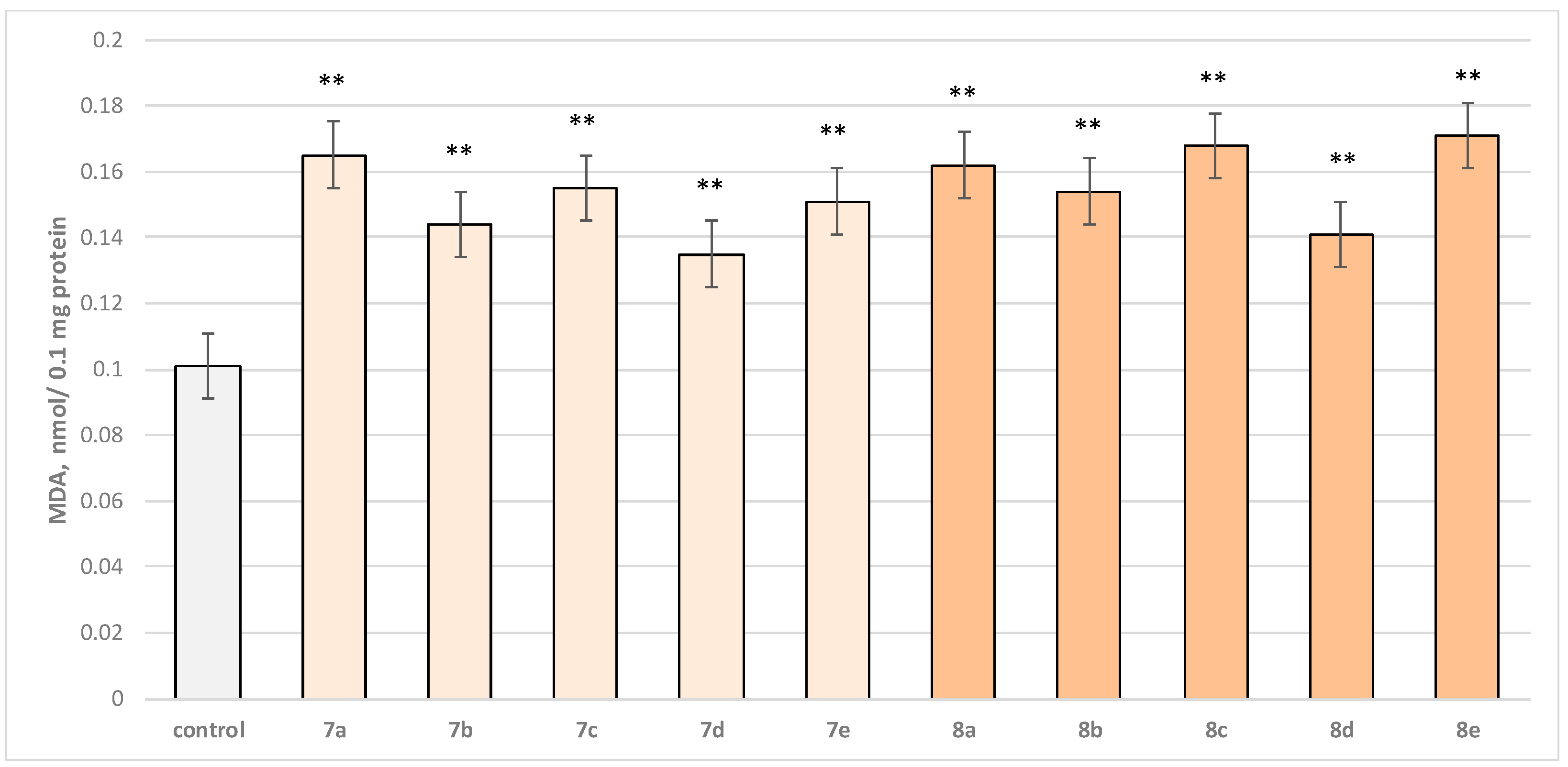








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds IDs | IC50 (EC50), (µM ± SD) hMAOA | IC50 (EC50), (µM ± SD) hMAOB | SI |
|---|---|---|---|
| 7a | >100 | 0.812 ± 0.10 | >123 |
| 7b | >100 | 0.551 ± 0.10 | >181 |
| 7c | >100 | 0.815 ± 0.20 | >123 |
| 7d | >100 | 0.491 ± 0.09 | >204 |
| 7e | >100 | 0.810 ± 0.09 | >123 |
| 8a | >100 | 0.821 ± 0.10 | >122 |
| 8b | >100 | 0.815 ± 0.10 | >123 |
| 8c | >100 | 0.814 ± 0.20 | >123 |
| 8d | >100 | 0.490 ± 0.09 | >204 |
| 8e | >100 | 0.817 ± 0.09 | >122 |
| Selegiline | - | 0.320 ± 0.20 | |
| Clorgilin | 18.74 ± 0.096 | - | >123 |
| Compound | Glide kcal/mol | ChemPLP | IFD kcal/mol |
|---|---|---|---|
| 7a | n.a | 39.81 | n.a |
| 7b | n.a | 50.47 | n.a |
| 7c | n.a | 22.19 | n.a |
| 7d | n.a | 13.58 | n.a |
| 7e | n.a | 33.67 | n.a |
| 8a | n.a | 36.54 | n.a |
| 8b | n.a | 43.09 | n.a |
| 8c | n.a | 35.57 | n.a |
| 8d | n.a | 0.68 | n.a |
| 8e | n.a | 29.85 | n.a |
| Harmine | −6.05 | 78.50 | −9.53 |
| Compound | Glide kcal/mol | ChemPLP Fitness Score | IFD kcal/mol | Intermolecular Stabilization |
|---|---|---|---|---|
| 7a | n.a | 126.52 | −13.80 | H-bonds (Tyr188; Tyr435); π–π (Tyr326) |
| 7b | n.a | 121.99 | −13.71 | H-bonds (Cys172; Ile198); π–π (Tyr326; Tyr435 |
| 7c | n.a | 117.94 | −13.83 | H-bond (Gln206); π–π (Tyr60; Tyr398) |
| 7d | n.a | 103.00 | −14.45 | H-bond (Tyr435) |
| 7e | n.a | 115.90 | −13.66 | Halogen bond (Trp119) |
| 8a | n.a | 136.49 | −13.16 | π–π (Trp119) |
| 8b | n.a | 137.88 | −14.27 | H-bond (Cys172); π–π (Phe343; Tyr398) |
| 8c | n.a | 133.28 | −13.73 | H-bond (Cys172); π–π (Phe343) |
| 8d | n.a | 121.03 | −14.65 | H-bonds (Pro102; Cys172 Gln206); π–π (Tyr398) |
| 8e | n.a | 123.95 | −14.55 | H-bonds (Cys172; Tyr326); π–π (Tyr326; Phe343; Tyr398) |
| ** Safinamide | −12.19 | 168.27 | −15.20 | H-bonds (Gln206; H2O) |
| PAMPA ID | Mw g/mol | PAMPA BBB −logPe |
|---|---|---|
| 7 | 380.00 | 4.864 ± 0.001 |
| 7a | 511.42 | 4.849 ± 0.043 |
| 7b | 502.80 | 4.416 ± 0.022 |
| 7c | 513.35 | 4.601 ± 0.040 |
| 7d | 544.40 | 4.819 ± 0.004 |
| 7e | 528.40 | 4.814 ± 0.15 |
| 8 | 393.28 | 4.538 ± 0.001 |
| 8a | 525.45 | 5.268 ± 0.014 |
| 8b | 516.82 | 4.444 ± 0.004 |
| 8c | 527.37 | 4.631 ± 0.047 |
| 8d | 558.43 | 4.464 ± 0.026 |
| 8e | 542.43 | 5.714 ± 0.041 |
| Theophylline | 180.167 | 6.553 ± 0.025 low |
| Corticosterone | 346.467 | 5.202 ± 0.020 medium |
| Propranolol HCl | 259.349 | 4.389 ± 0.014 high |
| ID | PAMPA BBB −logPe | MW g/mol | logP | pKa | fA | PSA, Å2 | FRB | HBD | HBA |
|---|---|---|---|---|---|---|---|---|---|
| 7b | 4.416 | 502.79 | 4.79 | 10.64 | 0.00 | 72.69 | 8 | 1 | 6 |
| 7d | 4.819 | 544.39 | 3.84 | 9.18 | 0.02 | 111.38 | 11 | 2 | 9 |
| 8d | 4.464 | 558.42 | 4.04 | 12.19 | 0.00 | 111.38 | 12 | 2 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgieva, M.; Mateev, E.; Valkova, I.; Kuteva, H.; Tzankova, D.; Stefanova, D.; Yordanov, Y.; Lybomirova, K.; Zlatkov, A.; Tzankova, V.; et al. Neurotoxicity, Neuroprotection, In Vitro MAOA/MAOB Inhibitory Activity Assessment, Molecular Docking, and Permeability Assay Studies of Newly Synthesized Hydrazones Containing a Pyrrole Ring. Molecules 2024, 29, 4338. https://doi.org/10.3390/molecules29184338
Georgieva M, Mateev E, Valkova I, Kuteva H, Tzankova D, Stefanova D, Yordanov Y, Lybomirova K, Zlatkov A, Tzankova V, et al. Neurotoxicity, Neuroprotection, In Vitro MAOA/MAOB Inhibitory Activity Assessment, Molecular Docking, and Permeability Assay Studies of Newly Synthesized Hydrazones Containing a Pyrrole Ring. Molecules. 2024; 29(18):4338. https://doi.org/10.3390/molecules29184338
Chicago/Turabian StyleGeorgieva, Maya, Emilio Mateev, Iva Valkova, Hristina Kuteva, Diana Tzankova, Denitsa Stefanova, Yordan Yordanov, Karolina Lybomirova, Alexander Zlatkov, Virginia Tzankova, and et al. 2024. "Neurotoxicity, Neuroprotection, In Vitro MAOA/MAOB Inhibitory Activity Assessment, Molecular Docking, and Permeability Assay Studies of Newly Synthesized Hydrazones Containing a Pyrrole Ring" Molecules 29, no. 18: 4338. https://doi.org/10.3390/molecules29184338
APA StyleGeorgieva, M., Mateev, E., Valkova, I., Kuteva, H., Tzankova, D., Stefanova, D., Yordanov, Y., Lybomirova, K., Zlatkov, A., Tzankova, V., & Kondeva-Burdina, M. (2024). Neurotoxicity, Neuroprotection, In Vitro MAOA/MAOB Inhibitory Activity Assessment, Molecular Docking, and Permeability Assay Studies of Newly Synthesized Hydrazones Containing a Pyrrole Ring. Molecules, 29(18), 4338. https://doi.org/10.3390/molecules29184338