Effects of L-Proline on the Stability of Mulberry Anthocyanins and the Mechanism of Interaction between L-Proline and Cyanidin-3-O-Glycoside


Abstract

1. Introduction
2. Results and Discussion
2.1. Effects of Amino Acids on Mulberry Anthocyanins in the Presence of Constant Light and AA
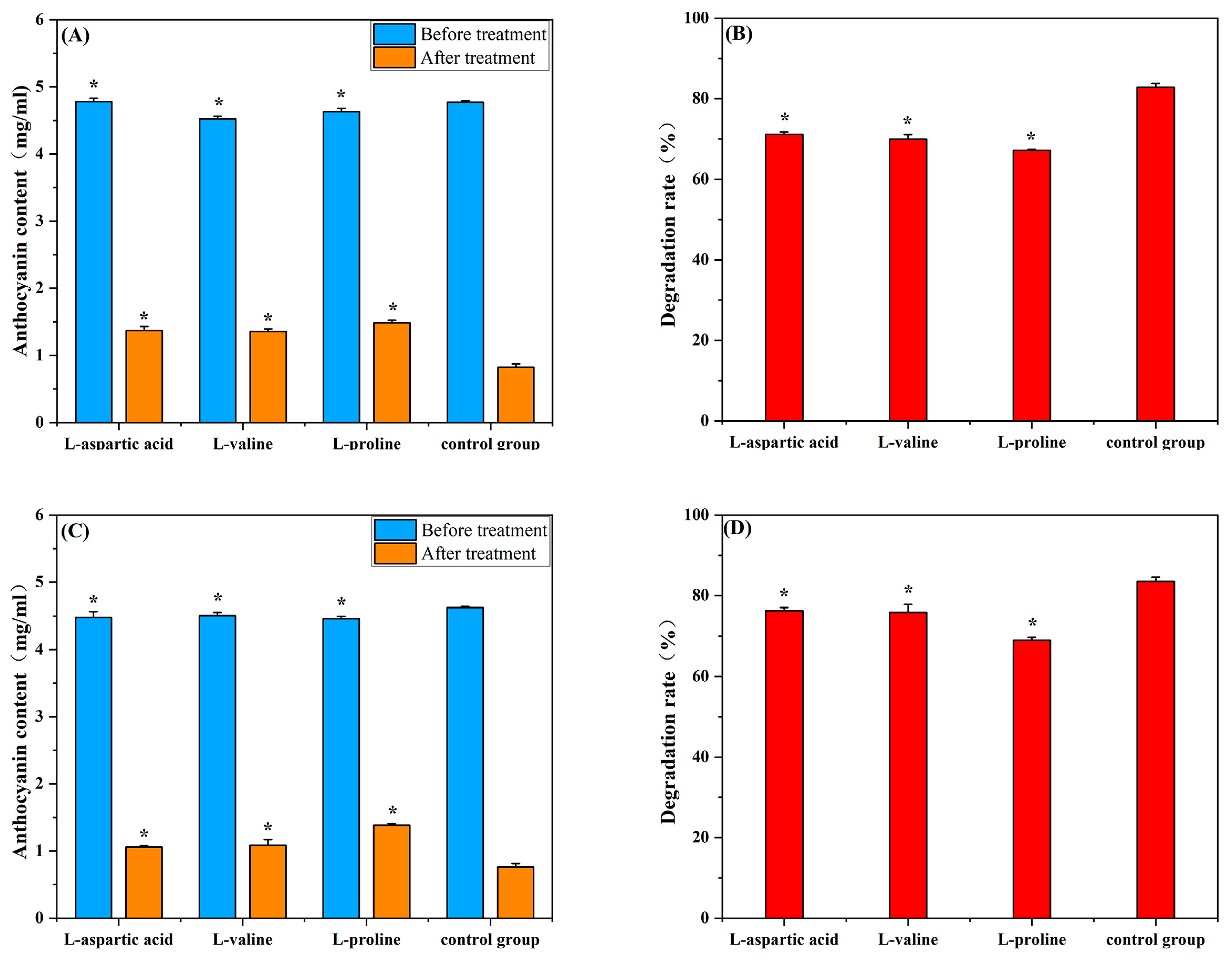
2.2. Effects of Amino Acids on Degradation Kinetics of Mulberry Anthocyanins
2.3. Fourier Infrared Spectroscopy
2.4. XRD Pattern
2.5. NMR Hydrogen Spectrum Analysis

2.6. Molecular Docking Analysis
2.7. Molecular Dynamics Simulation Results

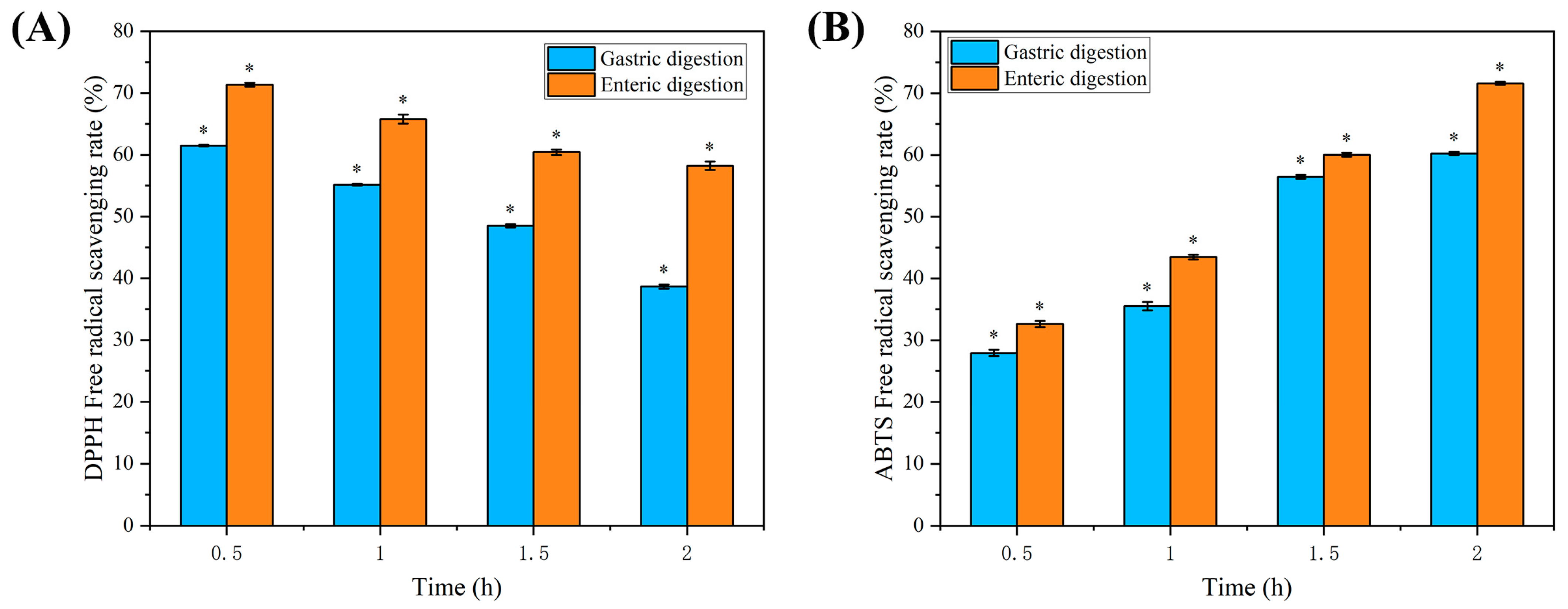
2.8. Analysis of the Results of In Vitro Assay for Digestive Antioxidant Activity
3. Materials and Methods
3.1. Materials and Ingredients
3.2. Mulberry Anthocyanin Extraction
3.3. Preparation of Mulberry Anthocyanin–Amino Acid Solution
3.4. Anthocyanin—Amino Acid—AA Mixed Solution Preparation
3.5. Estimation of Mulberry Anthocyanins
3.6. Analysis of Mulberry Anthocyanin Degradation Kinetics
3.7. Spectrum Instrumental Analysis
3.7.1. FT-IR Analysis
3.7.2. X-ray Diffraction Analysis
3.7.3. 1H NMR Analysis
3.8. Molecular Docking
3.9. Molecular Dynamics Approach to Simulation Calculations
3.10. Preparation of Simulated Digested Samples
3.11. Determination of Antioxidant Activity
3.11.1. Assay for DPPH Free Radical Scavenging Capacity
3.11.2. Assay for ABTS Free Radical Scavenging Capacity
3.12. Data Handling and Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, Q.; Zhao, L. The Mulberry (Morus alba L.) Fruit—A Review of Characteristic Components and Health Benefits. J. Agric. Food Chem. 2017, 65, 10383–10394. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Liu, F.; Xiong, L. Medicinal Parts of Mulberry (Leaf, Twig, Root Bark, and Fruit) and Compounds Thereof Are Excellent Traditional Chinese Medicines and Foods for Diabetes Mellitus. J. Funct. Foods 2023, 106, 105619. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Serali, O.; Unal, N.; Capanoglu, E. Antioxidant Activity and Polyphenol Composition of Black Mulberry (Morus nigra L.) Products. J. Berry Res. 2013, 3, 41–51. [Google Scholar] [CrossRef]
- Li, Y. Enhanced Physicochemical Stabilities of Cyanidin-3-O-Glucoside via Combination with Silk Fibroin Peptide. Food Chem. 2021, 355, 129479. [Google Scholar] [CrossRef]
- Hao, X.; Guan, R.; Huang, H.; Yang, K.; Wang, L.; Wu, Y. Anti-inflammatory Activity of cyanidin-3-O-glucoside and cyanidin-3-O-glucoside Liposomes in THP-1 Macrophages. Food Sci. Nutr. 2021, 9, 6480–6491. [Google Scholar] [CrossRef]
- Cui, H.; Guo, J.; Wang, D.; Yang, J. Stability of anthocyanins during processing and research progress. Food Ferment. Ind. 2024, 50, 388–397, (In Chinese, with English abstract). [Google Scholar] [CrossRef]
- Türkyılmaz, M.; Hamzaoğlu, F.; Ünal, H.; Özkan, M. Influence of Amino Acid Addition on the Thermal Stability of Anthocyanins in Pomegranate (Punica granatum L., Cv. Hicaznar) and Orange (Citrus sinensis L. Osbeck, Cv. Valencia) Juice Blend. Food Chem. 2022, 370, 131061. [Google Scholar] [CrossRef]
- Bingöl, A.; Türkyılmaz, M.; Özkan, M. Increase in Thermal Stability of Strawberry Anthocyanins with Amino Acid Copigmentation. Food Chem. 2022, 384, 132518. [Google Scholar] [CrossRef] [PubMed]
- Gençdağ, E.; Özdemir, E.E.; Demirci, K.; Görgüç, A.; Yılmaz, F.M. Copigmentation and Stabilization of Anthocyanins Using Organic Molecules and Encapsulation Techniques. Curr. Plant Biol. 2022, 29, 100238. [Google Scholar] [CrossRef]
- Nie, M.; Wang, L.; Lu, S.; Wang, Y.; Zheng, M.; Fang, Z. Protective Effect of Amino Acids on the Stability of Bayberry Anthocyanins and the Interaction Mechanism between L-Methionine and Cyanidin-3-O-Glycoside. Food Chem. 2022, 396, 133689. [Google Scholar] [CrossRef]
- Wu, D.; Li, Z.; Zhang, Y.; Zhang, Y.; Ren, G.; Zeng, Y.; Liu, H.; Guan, W.; Zhao, X.; Li, P.; et al. Proline Uptake Promotes Activation of Lymphoid Tissue Inducer Cells to Maintain Gut Homeostasis. Nat. Metab. 2023, 5, 1953–1968. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Li, N.; Han, X.; Guo, J.; Zhao, Y.; Liu, G.; Huang, W.; Zhan, J. Influence of Different Sterilization Treatments on the Color and Anthocyanin Contents of Mulberry Juice during Refrigerated Storage. Innov. Food Sci. Emerg. Technol. 2018, 48, 1–10. [Google Scholar] [CrossRef]
- Zang, Z.; Chou, S.; Geng, L.; Si, X.; Ding, Y.; Lang, Y.; Cui, H.; Gao, N.; Chen, Y.; Wang, M.; et al. Interactions of Blueberry Anthocyanins with Whey Protein Isolate and Bovine Serum Protein: Color Stability, Antioxidant Activity, in Vitro Simulation, and Protein Functionality. LWT 2021, 152, 112269. [Google Scholar] [CrossRef]
- Chen, Y.; Belwal, T.; Xu, Y.; Ma, Q.; Li, D.; Li, L.; Xiao, H.; Luo, Z. Updated Insights into Anthocyanin Stability Behavior from Bases to Cases: Why and Why Not Anthocyanins Lose during Food Processing. Crit. Rev. Food Sci. Nutr. 2023, 63, 8639–8671. [Google Scholar] [CrossRef] [PubMed]
- Sendri, N.; Singh, S.; Sharma, B.; Purohit, R.; Bhandari, P. Effect of Co-Pigments on Anthocyanins of Rhododendron Arboreum and Insights into Interaction Mechanism. Food Chem. 2023, 426, 136571. [Google Scholar] [CrossRef]
- Zhang, Y.-j. The Copigmentation Effect of Amino Acid on Blackberry Anthocyanin Stability and The Mechanism Investigation. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2018. [Google Scholar]
- Fan, L.; Wang, Y.; Xie, P.; Zhang, L.; Li, Y.; Zhou, J. Copigmentation Effects of Phenolics on Color Enhancement and Stability of Blackberry Wine Residue Anthocyanins: Chromaticity, Kinetics and Structural Simulation. Food Chem. 2019, 275, 299–308. [Google Scholar] [CrossRef]
- Lin, M.; Sun, C.; Gao, Q.; Zhang, Z.; Liang, Y.; Wang, S. Effect of Five Polyphenols on the Stability of Purple Cabbage Anthocyanins in Simulated Beverage Systems Containing L-Ascorbic Acid. Food Packag. Shelf Life 2023, 37, 101065. [Google Scholar] [CrossRef]
- Cheng, J.; Liu, J.-H.; Prasanna, G.; Jing, P. Spectrofluorimetric and Molecular Docking Studies on the Interaction of Cyanidin-3-O-Glucoside with Whey Protein, β-Lactoglobulin. Int. J. Biol. Macromol. 2017, 105, 965–972. [Google Scholar] [CrossRef]
- Huang, J.; Hu, Z.; Chin, Y.; Pei, Z.; Yao, Q.; Chen, J.; Li, D.; Hu, Y. Improved Thermal Stability of Roselle Anthocyanin by Co-Pigmented with Oxalic Acid: Preparation, Characterization and Enhancement Mechanism. Food Chem. 2023, 410, 135407. [Google Scholar] [CrossRef]
- Xing, C.; Chen, P.; Zhang, L. Computational Insight into Stability-Enhanced Systems of Anthocyanin with Protein/Peptide. Food Chem. Mol. Sci. 2023, 6, 100168. [Google Scholar] [CrossRef]
- Kou, X.; Su, D.; Pan, F.; Xu, X.; Meng, Q.; Ke, Q. Molecular Dynamics Simulation Techniques and Their Application to Aroma Compounds/Cyclodextrin Inclusion Complexes: A Review. Carbohydr. Polym. 2024, 324, 121524. [Google Scholar] [CrossRef] [PubMed]
- Sajini, T.; Mathew, B. A Brief Overview of Molecularly Imprinted Polymers: Highlighting Computational Design, Nano and Photo-Responsive Imprinting. Talanta Open 2021, 4, 100072. [Google Scholar] [CrossRef]
- Zhong, Z.; Huang, Z. The computation basis of software CS Chem3D and its application in organic chemistry. Guang Zhou Chem. Ind. Technol. 2002, 2, 20–23, (In Chinese, with English abstract). [Google Scholar]
- Zhao, L.; Pan, F.; Mehmood, A.; Zhang, Y.; Hao, S.; Rehman, A.U.; Li, J.; Wang, C.; Wang, Y. Protective Effect and Mechanism of Action of Xanthan Gum on the Color Stability of Black Rice Anthocyanins in Model Beverage Systems. Int. J. Biol. Macromol. 2020, 164, 3800–3807. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wu, T.; Li, Y.; Liu, D.; Sun, J.; Bai, W. Study on the Mechanism of Interaction between Mulberry Anthocyanins and Yeast Mannoprotein. Food Chem. 2023, 405, 135024. [Google Scholar] [CrossRef]
- Zhao, X.; Ding, B.-W.; Qin, J.-W.; He, F.; Duan, C.-Q. Intermolecular Copigmentation between Five Common 3-O-Monoglucosidic Anthocyanins and Three Phenolics in Red Wine Model Solutions: The Influence of Substituent Pattern of Anthocyanin B Ring. Food Chem. 2020, 326, 126960. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, N.; Wu, G.-F.; He, F.; Lan, Y.-B.; Duan, C.-Q. Intermolecular Copigmentation between Anthocyanidin-3,5-O-Diglucosides and Three Phenolic Compounds: Insights from Experimental and Theoretical Studies. Food Chem. Adv. 2022, 1, 100111. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total Phenolics, Flavonoids, Anthocyanins and Antioxidant Activity Following Simulated Gastro-Intestinal Digestion and Dialysis of Apple Varieties: Bioaccessibility and Potential Uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Y.; Deng, H.; Yuan, H.; Fan, X.; Yang, H.; Tan, S. In Vitro and in Vivo Bioaccessibility, Antioxidant Activity, and Color of Red Radish Anthocyanins as Influenced by Different Drying Methods. Food Chem. X 2023, 18, 100633. [Google Scholar] [CrossRef]
- Liang, L.; Wu, X.; Zhao, T.; Zhao, J.; Li, F.; Zou, Y.; Mao, G.; Yang, L. In Vitro Bioaccessibility and Antioxidant Activity of Anthocyanins from Mulberry (Morus atropurpurea Roxb.) Following Simulated Gastro-Intestinal Digestion. Food Res. Int. 2012, 46, 76–82. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In Vitro Bio-Accessibility and Antioxidant Activity of Grape Polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Wang, D.-Q.; Cui, H.; Zong, K.-L.; Hu, H.; Yang, J.-T. Extraction of Functional Natural Products Employing Microwave-Assisted Aqueous Two-Phase System: Application to Anthocyanins Extraction from Mulberry Fruits. Prep. Biochem. Biotechnol. 2023, 54, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, B.; Zhang, D.; Xiu, Z. Aqueous Two-Phase Extraction of 1,3-Propanediol from Glycerol-Based Fermentation Broths. Sep. Purif. Technol. 2009, 66, 472–478. [Google Scholar] [CrossRef]
- Yuan, Y.; Tian, Y.; Gao, S.; Zhang, X.; Gao, X.; He, J. Effects of Environmental Factors and Fermentation on Red Raspberry Anthocyanins Stability. LWT 2023, 173, 114252. [Google Scholar] [CrossRef]
- Hou, Y.; Vasileva, E.A.; Mishchenko, N.P.; Carne, A.; McConnell, M.; Bekhit, A.E.-D.A. Extraction, Structural Characterization and Stability of Polyhydroxylated Naphthoquinones from Shell and Spine of New Zealand Sea Urchin (Evechinus chloroticus). Food Chem. 2019, 272, 379–387. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, C.; Zhang, J.; Zhang, L. Interaction of Preheated Whey Protein Isolate with Rose Anthocyanin Extracts in Beverage Model System: Influence on Color Stability, Astringency and Mechanism. Food Chem. 2023, 412, 135507. [Google Scholar] [CrossRef]
- Chandra Singh, M.; Price, W.E.; Kelso, C.; Charlton, K.; Probst, Y. Impact of Molar Absorbance on Anthocyanin Content of the Foods. Food Chem. 2022, 386, 132855. [Google Scholar] [CrossRef]
- Pham, T.N.; Le, X.T.; Pham, V.T.; Le, H.T. Effects of Process Parameters in Microwave-Assisted Extraction on the Anthocyanin-Enriched Extract from Rhodomyrtus tomentosa (Ait.) Hassk and Its Storage Conditions on the Kinetic Degradation of Anthocyanins in the Extract. Heliyon 2022, 8, e09518. [Google Scholar] [CrossRef]
- Cheng, Y.; Chen, X.; Yang, T.; Wang, Z.; Chen, Q.; Zeng, M.; Qin, F.; Chen, J.; He, Z. Storage Stability and Multi-Spectroscopy Analysis of the Ternary Complex Induced by Mulberry Anthocyanin Extract Interacting with Whey Protein Isolate and Rutin under Acidic Conditions. Food Hydrocoll. 2023, 143, 108911. [Google Scholar] [CrossRef]
- Khodaiyan, F.; Parastouei, K. Co-Optimization of Pectin and Polyphenols Extraction from Black Mulberry Pomace Using an Eco-Friendly Technique: Simultaneous Recovery and Characterization of Products. Int. J. Biol. Macromol. 2020, 164, 1025–1036. [Google Scholar] [CrossRef]
- Ma, Y.-L.; Wang, Y.; Wu, Z.-F.; Mei, J.; Zhang, W.-Q.; Shang, Y.-F.; Thakur, K.; Wei, Z.-J. Exploring the Effect of in Vitro Digestion on the Phenolics and Antioxidant Activity of Lycium Barbarum Fruit Extract. Food Biosci. 2023, 51, 102255. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constant Light | Ascorbic Acid | |||||
|---|---|---|---|---|---|---|
| k (day−1) | t1/2 (day) | R2 | k (day−1) | t1/2 (day) | R2 | |
| Control group | 0.23133 | 2.9964 | 0.95705 | 0.23866 | 2.9043 | 0.96410 |
| L-aspartic acid group | 0.19740 | 3.5114 | 0.98726 | 0.20741 | 3.3419 | 0.86409 |
| L-valine group | 0.19885 | 3.4858 | 0.90633 | 0.21639 | 3.2032 | 0.94333 |
| L-proline group | 0.19313 | 3.5890 | 0.94759 | 0.17852 | 3.8827 | 0.92333 |
| MODE | Affinity (kcal/mol) | Distance from Best Mode | Binding Conformation | |
|---|---|---|---|---|
| Rmsd l.b. | Rmsd u.b. | |||
| 1 | −1.7 | 0 | 0 |  |
| 2 | −1.7 | 7.116 | 9.627 |  |
| 3 | −1.6 | 5.650 | 8.191 |  |
| 4 | −1.6 | 5.437 | 7.887 | 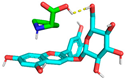 |
| 5 | −1.6 | 3.321 | 8.037 | 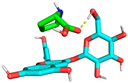 |
| 6 | −1.6 | 6.442 | 9.144 |  |
| 7 | −1.6 | 6.994 | 9.600 |  |
| 8 | −1.6 | 5.841 | 9.045 | 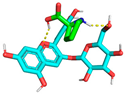 |
| 9 | −1.5 | 7.258 | 8.747 |  |
| Stretching | Bending | Stretch-Bending | Torsion | Non-1,4 VDW | 1,4 VDW | Dipole/Dipole | Total | |
|---|---|---|---|---|---|---|---|---|
| Initial | 0.5107 | 9.2850 | 0.3184 | 17.2005 | −21.5507 | 16.1389 | −15.6499 | 16.2466 |
| Termination | 0.3996 | 5.8711 | 0.2272 | 15.8223 | −24.0872 | 13.5361 | −11.6041 | 0.1649 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, H.; Li, X.; Ji, Y.; Zhao, S.; Yang, J. Effects of L-Proline on the Stability of Mulberry Anthocyanins and the Mechanism of Interaction between L-Proline and Cyanidin-3-O-Glycoside. Molecules 2024, 29, 4544. https://doi.org/10.3390/molecules29194544
Cui H, Li X, Ji Y, Zhao S, Yang J. Effects of L-Proline on the Stability of Mulberry Anthocyanins and the Mechanism of Interaction between L-Proline and Cyanidin-3-O-Glycoside. Molecules. 2024; 29(19):4544. https://doi.org/10.3390/molecules29194544
Chicago/Turabian StyleCui, Haipeng, Xianbao Li, Yuan Ji, Shengxu Zhao, and Jianting Yang. 2024. "Effects of L-Proline on the Stability of Mulberry Anthocyanins and the Mechanism of Interaction between L-Proline and Cyanidin-3-O-Glycoside" Molecules 29, no. 19: 4544. https://doi.org/10.3390/molecules29194544
APA StyleCui, H., Li, X., Ji, Y., Zhao, S., & Yang, J. (2024). Effects of L-Proline on the Stability of Mulberry Anthocyanins and the Mechanism of Interaction between L-Proline and Cyanidin-3-O-Glycoside. Molecules, 29(19), 4544. https://doi.org/10.3390/molecules29194544