

Antibacterial and Antibiofilm Effects of Photodynamic Treatment with Curcuma L. and Trans-Cinnamaldehyde against Listeria monocytogenes
 , , and
, , and
Abstract
:1. Introduction
2. Results
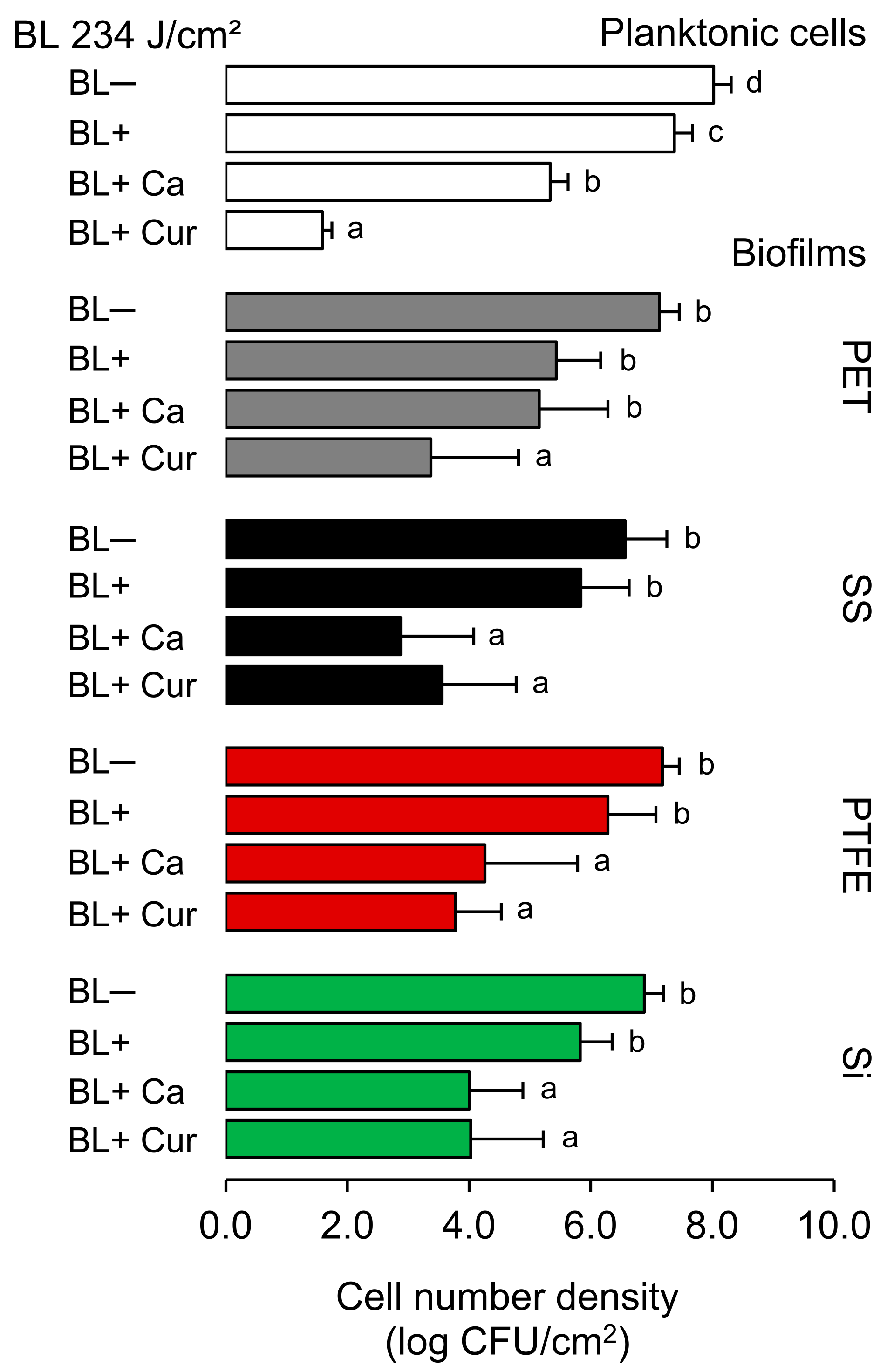
2.1. Antibacterial and Antibiofilm Activity
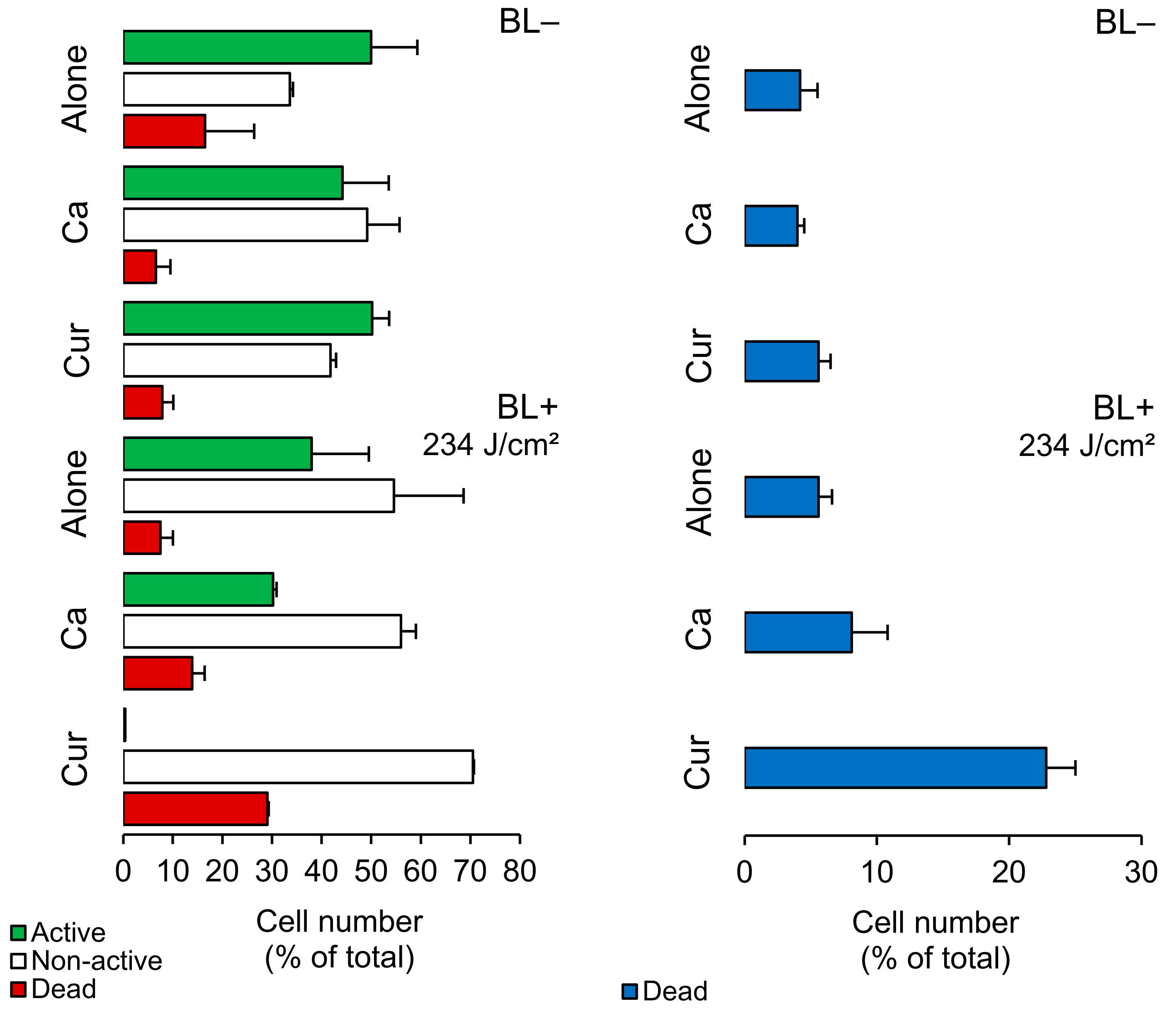
2.2. Planktonic Cells—FCM Analysis
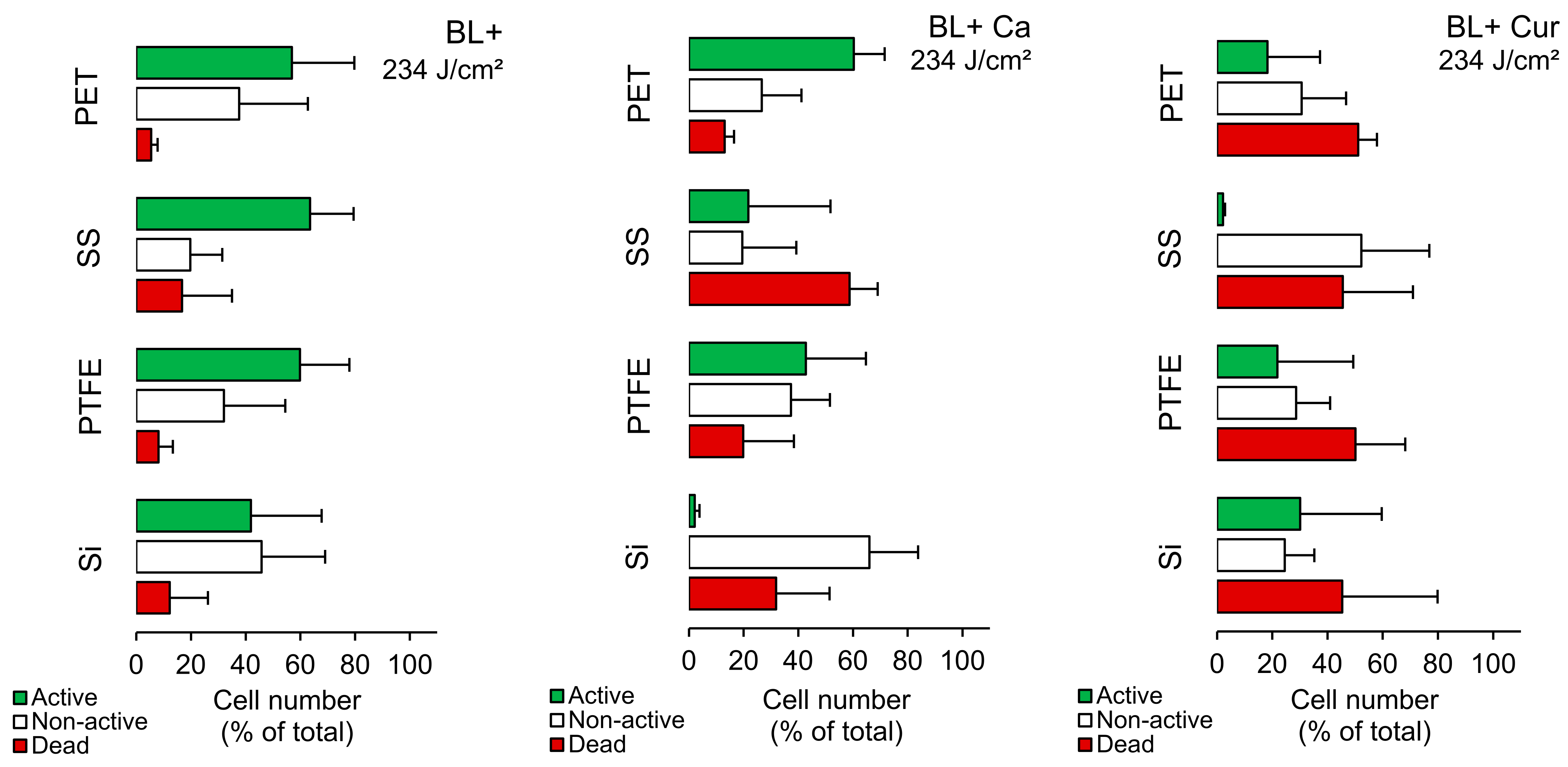
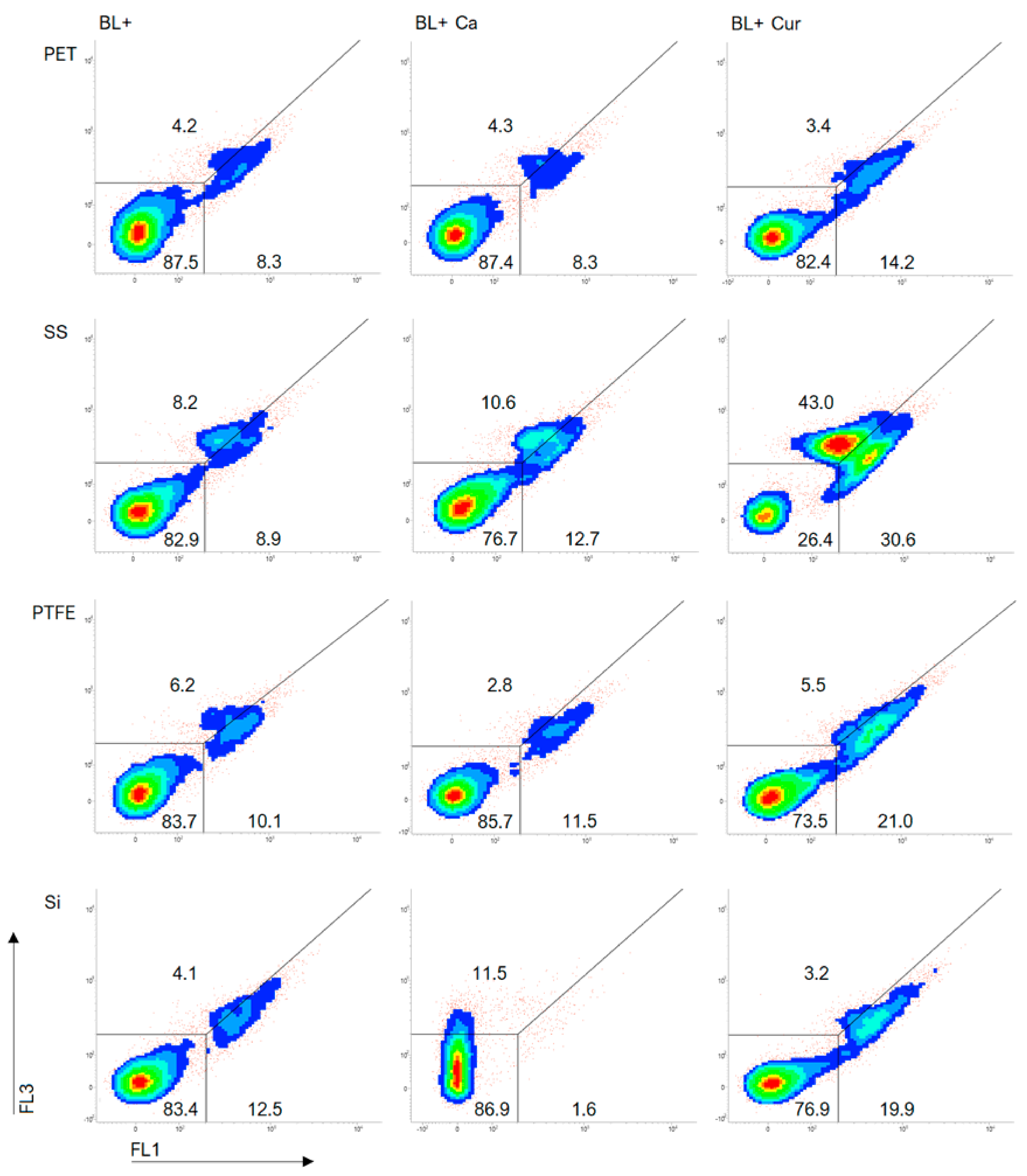
2.3. Biofilms—FCM Analysis
2.4. Adhesion Potential and Hydrophobicity
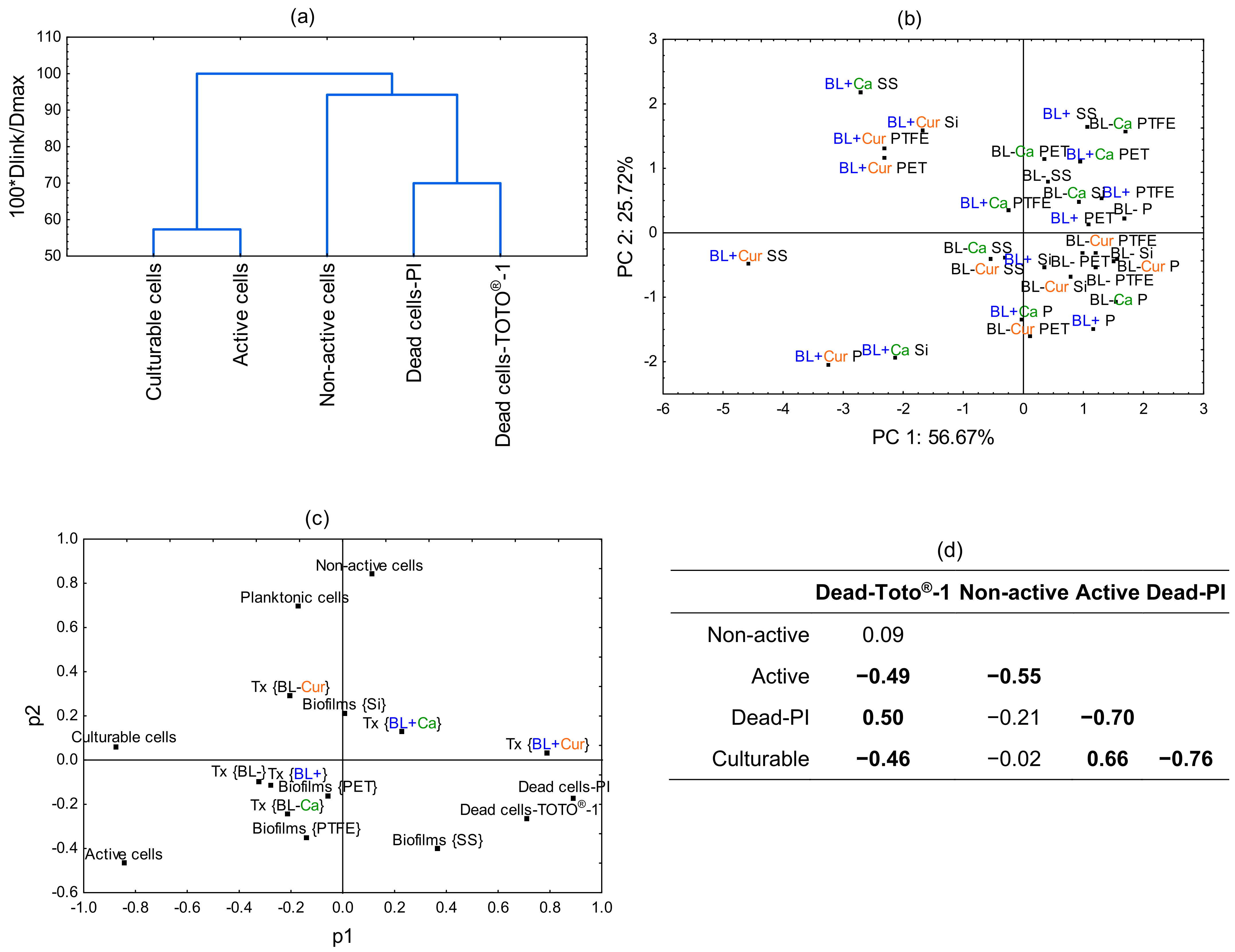
2.5. Principal Component Analysis (PCA)
3. Discussion
4. Materials and Methods
4.1. Bacterium and Culture Conditions
4.2. Planktonic Cells and Biofilms
4.3. Material Surfaces
4.4. Hydrophobicity and Free Energy of Adhesion
4.5. Phytochemicals
4.6. Photodynamic Treatments
4.7. CFU Assay
4.8. FCM Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodríguez-Campos, D.; Rodríguez-Melcón, C.; Alonso-Calleja, C.; Capita, R. Persistent Listeria monocytogenes isolates from a poultry-processing facility form more biofilm but do not have a greater resistance to disinfectants than sporadic strains. Pathogens 2019, 8, 250. [Google Scholar] [CrossRef]
- Colagiorgi, A.; Bruini, I.; Di Ciccio, P.A.; Zanardi, E.; Ghidini, S.; Ianieri, A. Listeria monocytogenes biofilms in the wonderland of food industry. Pathogens 2017, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Leung, A.W.; Hua, H.; Rao, X.; Xu, C. Photodynamic action of LED-activated curcumin against Staphylococcus aureus involving intracellular ROS increase and membrane damage. Int. J. Photoenergy 2014, 2014, 637601. [Google Scholar] [CrossRef]
- Costa, L.; Tomé, J.P.; Neves, M.G.; Tomé, A.C.; Cavaleiro, J.A.; Faustino, M.A.; Cunha, Â.; Gomes, N.C.; Almeida, A. Evaluation of resistance development and viability recovery by a non-enveloped virus after repeated cycles of aPDT. Antiviral Res. 2011, 91, 278–282. [Google Scholar] [CrossRef]
- Zhu, S.; Song, Y.; Pei, J.; Xue, F.; Cui, X.; Xiong, X.; Li, C. The application of photodynamic inactivation to microorganisms in food. Food Chem. X 2021, 12, 100150. [Google Scholar] [CrossRef]
- Maisch, T. Strategies to optimize photosensitizers for photodynamic inactivation of bacteria. J. Photochem. Photobiol. B Biol. 2015, 150, 2–10. [Google Scholar] [CrossRef]
- Draszanowska, A.; Karpińska-Tymoszczyk, M.; Simões, M.; Olszewska, M.A. The effect of turmeric rhizome on the inhibition of lipid oxidation and the quality of vacuum-packed pork meatloaf. Appl. Sci. 2022, 12, 10641. [Google Scholar] [CrossRef]
- Dovigo, L.N.; Carmello, J.C.; De Souza Costa, C.A.; Vergani, C.E.; Brunetti, I.L.; Bagnato, V.S.; Pavarina, A.C. Curcumin-mediated photodynamic inactivation of Candida albicans in a murine model of oral candidiasis. Med. Mycol. 2013, 51, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, F.; Mereddy, R.; Sultanbawa, Y. The effect of photosensitization mediated by curcumin on storage life of fresh date (Phoenix dactylifera L.) fruit. Food Control 2018, 93, 305–309. [Google Scholar] [CrossRef]
- de Oliveira, E.F.; Tikekar, R.; Nitin, N. Combination of aerosolized curcumin and UV-A light for the inactivation of bacteria on fresh produce surfaces. Food Res. Int. 2018, 114, 133–139. [Google Scholar] [CrossRef]
- de Oliveira, E.F.; Tosati, J.V.; Tikekar, R.V.; Monteiro, A.R.; Nitin, N. Antimicrobial activity of curcumin in combination with light against Escherichia coli O157:H7 and Listeria innocua: Applications for fresh produce sanitation. Postharvest Biol. Technol. 2018, 137, 86–94. [Google Scholar] [CrossRef]
- Lin, Y.; Hu, J.; Li, S.; Hamzah, S.S.; Jiang, H.; Zhou, A.; Zeng, S.; Lin, S. Curcumin-based photodynamic sterilization for preservation of fresh-cut hami melon. Molecules 2019, 24, 2374. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, R.; Zhang, H.; Jin, G.-Q.; Wang, B.-W.; Zhu, M.; Zhang, J.; Gao, S.; Zhang, J.-L. Porpholactam-cinnamaldehyde conjugates for promoting ROS generation in photodynamic therapy. J. Porphyr. Phthalocyanines 2021, 25, 1111. [Google Scholar] [CrossRef]
- Bernardes, P.C.; de Andrade, N.J.; Ferreira, S.O.; de Sá, J.P.; Araújo, E.A.; Delatorre, D.M.; Luiz, L.M. Assessment of hydrophobicity and roughness of stainless steel adhered by an isolate of Bacillus cereus from a dairy plant. Braz. J. Microbiol. 2010, 41, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Lin, J.; Li, H.; Shen, Z.; Wang, Y.; Velkov, T.; Shen, J. The natural product curcumin as an antibacterial agent: Current achievements and problems. Antioxidants 2022, 11, 459. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chen, B.; Li, H.; Zeng, Q.-H.; Wang, J.J.; Liu, H.; Pan, Y.; Zhao, Y. Enhanced antibacterial and antibiofilm functions of the curcumin-mediated photodynamic inactivation against Listeria monocytogenes. Food Control 2020, 108, 106886. [Google Scholar] [CrossRef]
- Bonifácio, D.; Martins, C.; David, B.; Lemos, C.; Neves, M.G.P.M.S.; Almeida, A.; Pinto, D.C.G.A.; Faustino, M.A.F.; Cunha, Â. Photodynamic inactivation of Listeria innocua biofilms with food-grade photosensitizers: A curcumin-rich extract of Curcuma longa vs commercial curcumin. J. Appl. Microbiol. 2018, 125, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, Z.; Cao, B.; Wu, J.; Wang, Y.; Xue, Y.; Xu, J.; Xue, C.; Tang, Q.J. The effect of novel photodynamic activation method mediated by curcumin on oyster shelf life and quality. Food Res. Int. 2016, 87, 204–210. [Google Scholar] [CrossRef]
- Tao, R.; Zhang, F.; Tang, Q.-J.; Xu, C.-S.; Ni, Z.-J.; Meng, X.-H. Effects of curcumin-based photodynamic treatment on the storage quality of fresh-cut apples. Food Chem. 2019, 274, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Temba, B.A.; Fletcher, M.T.; Fox, G.P.; Harvey, J.; Okoth, S.A.; Sultanbawa, Y. Curcumin-based photosensitization inactivates Aspergillus flavus and reduces aflatoxin B1 in maize kernels. Food Microbiol. 2019, 82, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, M.A.; Dev Kumar, G.; Hur, M.; Diez-Gonzalez, F. Inactivation of dried cells and biofilms of Listeria monocytogenes by exposure to blue light at different wavelengths and the influence of surface materials. Appl. Environ. Microbiol. 2023, 89, e01147-23. [Google Scholar] [CrossRef]
- Lin, S.L.; Hu, J.M.; Tang, S.S.; Wu, X.Y.; Chen, Z.Q.; Tang, S.Z. Photodynamic inactivation of methylene blue and tungsten-halogen lamp light against food pathogen Listeria monocytogenes. Photochem. Photobiol. 2012, 88, 985–991. [Google Scholar] [CrossRef]
- Seidi Damyeh, M.; Mereddy, R.; Netzel, M.E.; Sultanbawa, Y. An insight into curcumin-based photosensitization as a promising and green food preservation technology. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1727–1759. [Google Scholar] [CrossRef]
- Liao, Q.; Kim, E.J.; Tang, Y.; Xu, H.; Yu, D.-G.; Song, W.; Kim, B.J. Rational design of hyper-crosslinked polymers for biomedical applications. J. Polym. Sci. 2023. early view. [Google Scholar] [CrossRef]
- Agel, M.R.; Baghdan, E.; Pinnapireddy, S.R.; Lehmann, J.; Schäfer, J.; Bakowsky, U. Curcumin loaded nanoparticles as efficient photoactive formulations against gram-positive and gram-negative bacteria. Colloids Surf. 2019, 178, 460–468. [Google Scholar] [CrossRef]
- Gonelimali, F.D.; Lin, J.; Miao, W.; Xuan, J.; Charles, F.; Chen, M.; Hatab, S.R. Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front. Microbiol. 2018, 9, 1639. [Google Scholar] [CrossRef]
- Khan, N.; Abbasi, A.M.; Dastagir, G.; Nazir, A.; Shah, G.M.; Shah, M.M.; Shah, M.H. Ethnobotanical and antimicrobial study of some selected medicinal plants used in Khyber Pakhtunkhwa (KPK) as a potential source to cure infectious diseases. BMC Complement. Altern. Med. 2014, 14, 122. [Google Scholar] [CrossRef]
- Lencova, S.; Zdenkova, K.; Demnerova, K.; Stiborova, H. Antibacterial and antibiofilm effect of natural substances and their mixtures over Listeria monocytogenes, Staphylococcus aureus and Escherichia coli. LWT 2022, 154, 112777. [Google Scholar] [CrossRef]
- Upadhyay, A.; Johny, A.K.; Amalaradjou, M.A.R.; Ananda Baskaran, S.; Kim, K.S.; Venkitanarayanan, K. Plant-derived antimicrobials reduce Listeria monocytogenes virulence factors in vitro, and down-regulate expression of virulence genes. Int. J. Food Microbiol. 2012, 157, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, L.; Han, J.; Dong, P.; Luo, X.; Zhang, Y.; Zhu, L. Inhibition of biofilm formation and related gene expression of Listeria monocytogenes in response to four natural antimicrobial compounds and sodium hypochlorite. Front. Microbiol. 2021, 11, 617473. [Google Scholar] [CrossRef] [PubMed]
- Gędas, A.; Draszanowska, A.; den Bakker, H.; Diez-Gonzalez, F.; Simões, M.; Olszewska, M.A. Prevention of surface colonization and anti-biofilm effect of selected phytochemicals against Listeria innocua strain. Colloids Surf. B 2023, 228, 113391. [Google Scholar] [CrossRef]
- Teixeira, P.; Lopes, Z.; Azeredo, J.; Oliveira, R.; Vieira, M.J. Physico-chemical surface characterization of a bacterial population isolated from a milking machine. Food Microbiol. 2005, 22, 247–251. [Google Scholar] [CrossRef]
- van Oss, C.J. Hydrophobicity and hydrophilicity of biosurfactants. Curr. Opin. Coll. Int. Sci. 1997, 2, 503–512. [Google Scholar] [CrossRef]
- Kawacka, I.; Olejnik-Schmidt, A.; Schmidt, M.; Sip, A. Natural plant-derived chemical compounds as Listeria monocytogenes inhibitors in vitro and in food model systems. Pathogens 2021, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Li, Y.; Wu, M.X. Bacteria-specific pro-photosensitizer kills multidrug-resistant Staphylococcus aureus and Pseudomonas aeruginosa. Commun. Biol. 2021, 4, 408. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Abreu, A.C.; Ferreira, C.; Saavedra, M.J.; Simões, L.C.; Simões, M. Antibacterial activity and mode of action of selected glucosinolate hydrolysis products against bacterial pathogens. J. Food Sci. Technol. 2015, 52, 4737–4748. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Simões, L.C.; Saavedra, M.J.; Simões, M. The action of selected isothiocyanates on bacterial biofilm prevention and control. Int. Biodeterior. Biodegrad. 2014, 86, 25–33. [Google Scholar] [CrossRef]
- van Oss, C.J.; Chaudhury, M.K.; Good, R.J. Monopolar surfaces. Adv. Colloid Interface Sci. 1987, 28, 35–64. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Free Energy of Adhesion (mJ/m2) | Contact Angles (°) * | Surface Tension Components (mJ/m2) | ΔGTOT (mJ/m2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Surface | ΔGblsLW | ΔGblsAB | ΔGadhesion | θW | θF | θB | γSLW | γS+ | γS− | γSAB | |
| PET | 3.4 | 3.4 | 0.0 | 57.0 ± 18.6 | 71.2 ± 4.3 | 65.2 ± 14.3 | 22.3 | 0 | 49.9 | 0 | 40.7 |
| SS | −3.2 | 6.2 | 2.9 | 53.2 ± 17.9 | 67.2 ± 3.1 | 54.5 ± 14.5 | 27.7 | 0 | 52.4 | 0 | 43.5 |
| PTFE | 0.0 | −4.4 | −4.4 | 63.0 ± 36.1 | 73.5 ± 10.2 | 59.8 ± 6.0 | 25.1 | 0 | 41.7 | 0 | 28.2 |
| Si | 2.4 | 1.3 | 3.7 | 70.8 ± 38.9 | 88.1 ± 10.6 | 59.6 ± 20.7 | 25.2 | 0 | 46.1 | 0 | 34.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimińska, A.; Lipska, I.; Gajewska, J.; Draszanowska, A.; Simões, M.; Olszewska, M.A. Antibacterial and Antibiofilm Effects of Photodynamic Treatment with Curcuma L. and Trans-Cinnamaldehyde against Listeria monocytogenes. Molecules 2024, 29, 685. https://doi.org/10.3390/molecules29030685
Zimińska A, Lipska I, Gajewska J, Draszanowska A, Simões M, Olszewska MA. Antibacterial and Antibiofilm Effects of Photodynamic Treatment with Curcuma L. and Trans-Cinnamaldehyde against Listeria monocytogenes. Molecules. 2024; 29(3):685. https://doi.org/10.3390/molecules29030685
Chicago/Turabian StyleZimińska, Aleksandra, Izabela Lipska, Joanna Gajewska, Anna Draszanowska, Manuel Simões, and Magdalena A. Olszewska. 2024. "Antibacterial and Antibiofilm Effects of Photodynamic Treatment with Curcuma L. and Trans-Cinnamaldehyde against Listeria monocytogenes" Molecules 29, no. 3: 685. https://doi.org/10.3390/molecules29030685