The Transcriptome of BT-20 Breast Cancer Cells Exposed to Curcumin Analog NC2603 Reveals a Relationship between EGR3 Gene Modulation and Cell Migration Inhibition

 , , , , , , , ,
, , , , , , , ,
Abstract

1. Introduction
2. Results
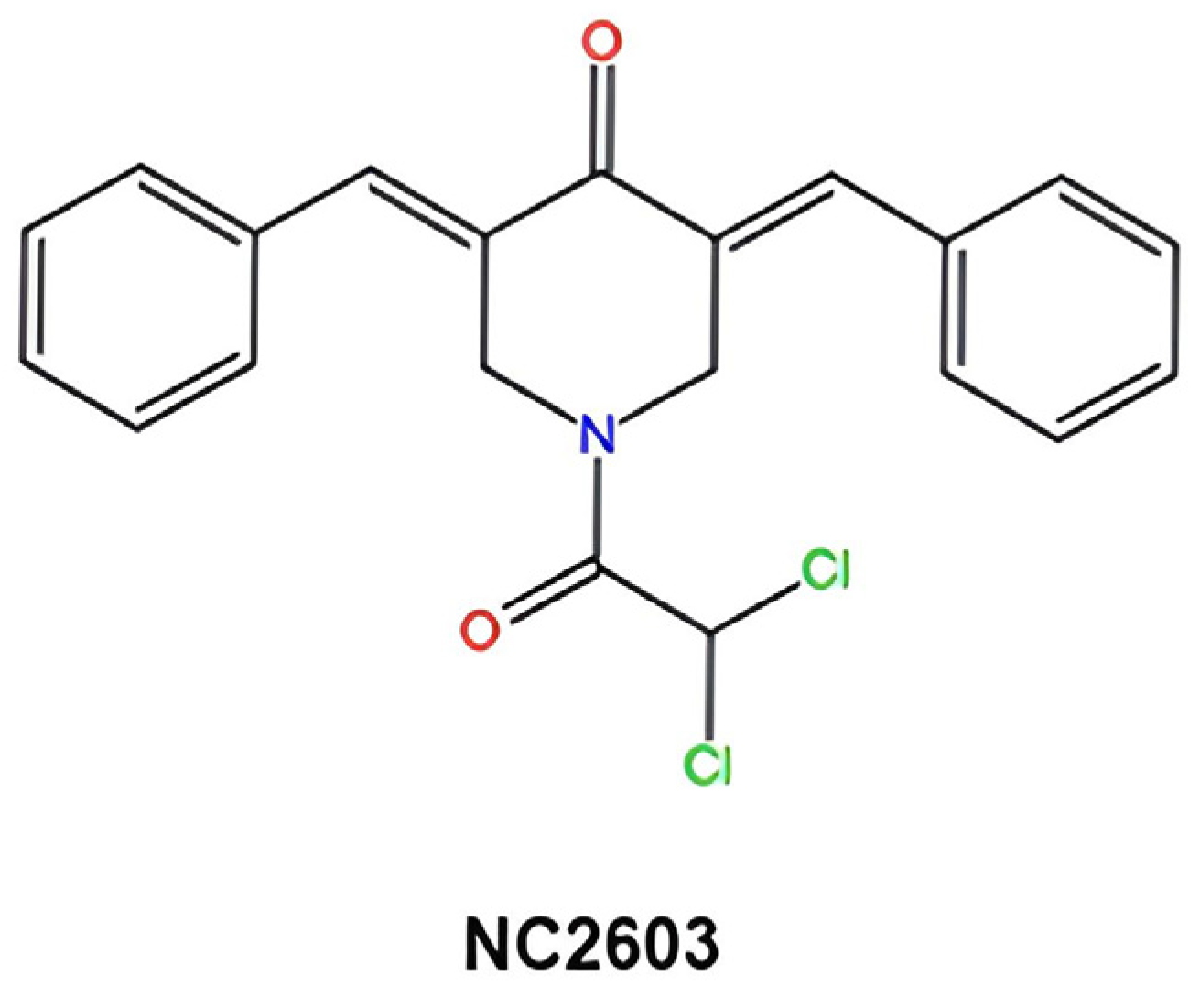
2.1. Cell Viability Suppression by Curcumin Analog NC2603
2.2. Cell Migration Inhibition by Curcumin and Analog NC2603 at 0.75 μM
2.3. Differential Gene Expression Analysis: Uncovering Key Genes Regulated by Analog NC2603
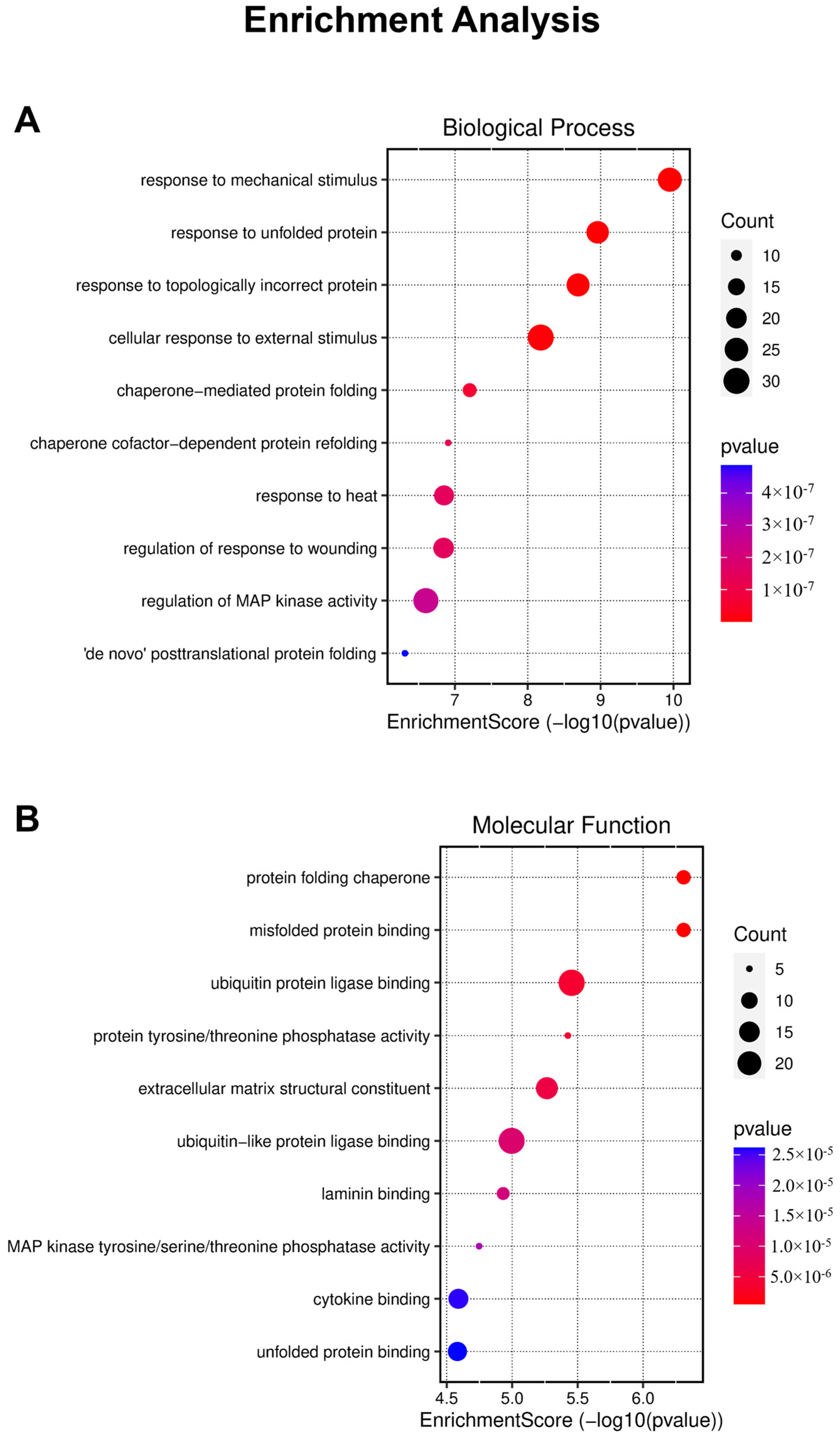
2.4. Enrichment Analysis
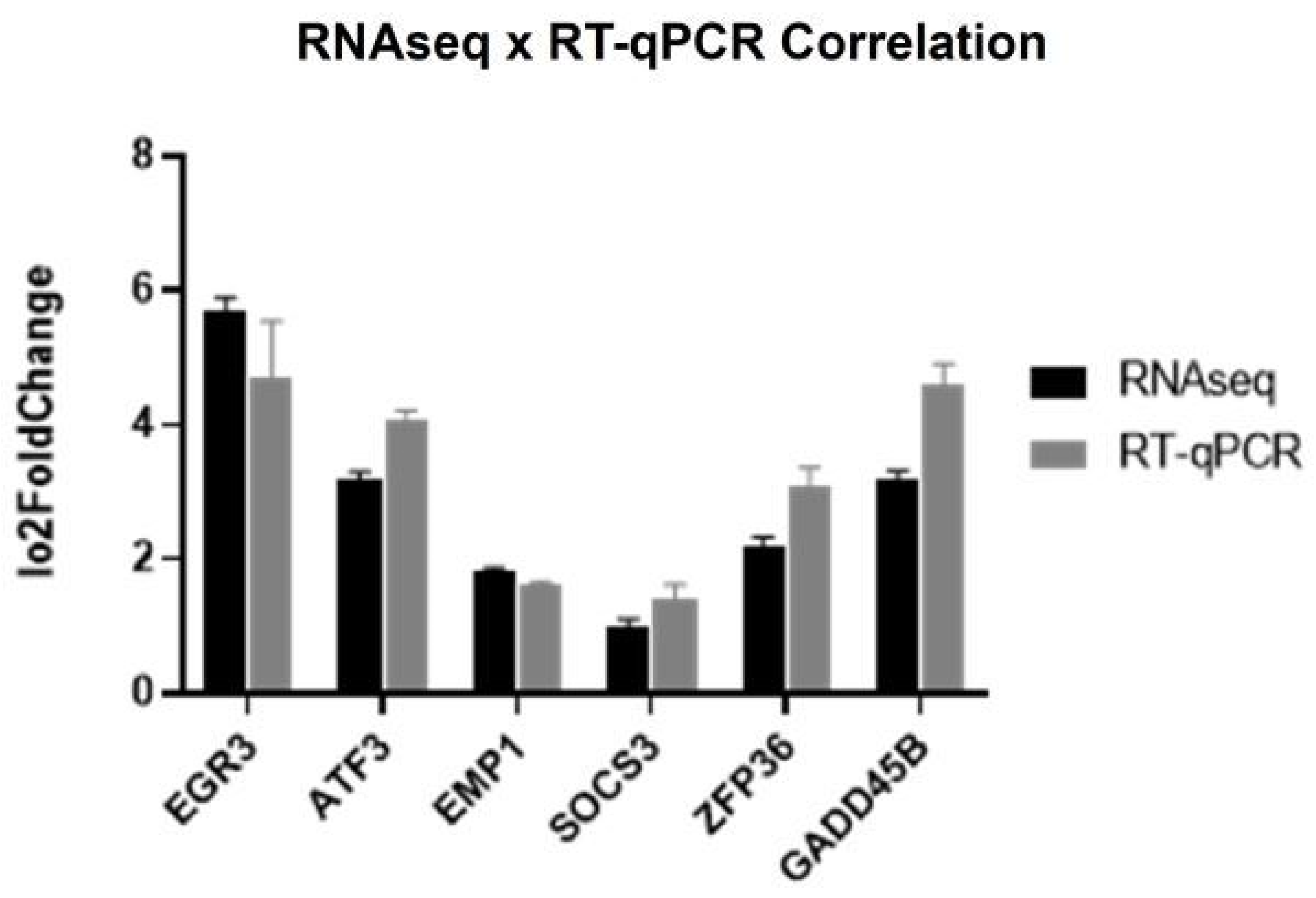
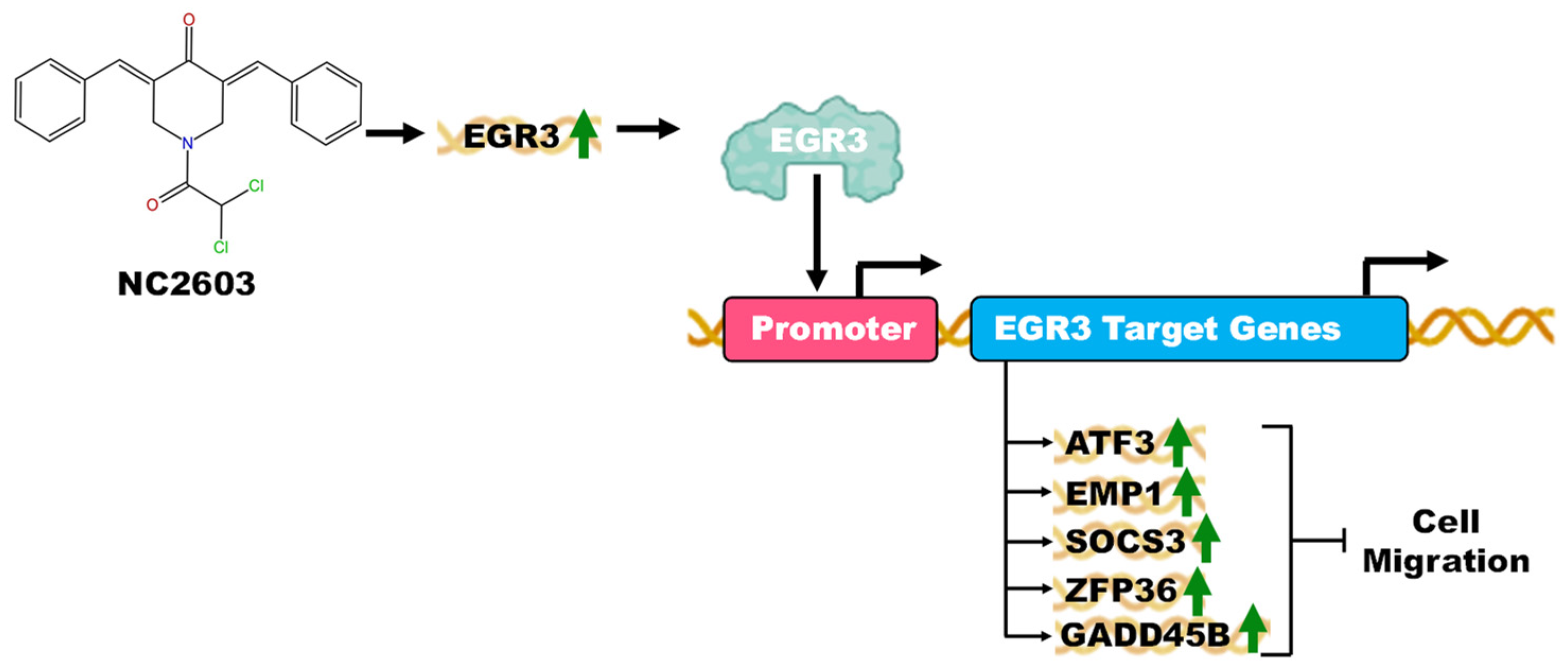
2.5. Validation of EGR3 Gene Targets: Unveiling the Migration Inhibition Mechanism
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Viability
4.4. Scratch Assay
4.5. Transwell Migration Assay
4.6. Next Generation Sequencing
4.7. RT-qPCR
4.8. Enrichment Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roser, M.; Ritchie, H. Cancer—Our World in Data. Available online: https://ourworldindata.org/cancer (accessed on 3 May 2022).
- Chen, W.; Hoffmann, A.D.; Liu, H.; Liu, X. Organotropism: New Insights into Molecular Mechanisms of Breast Cancer Metastasis. NPJ Precis. Oncol. 2018, 2, 4. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Pommier, Y. Topoisomerase I Inhibitors: Camptothecins and Beyond. Nat. Rev. Cancer 2006, 6, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Lou, H.X. Strategies to Diversify Natural Products for Drug Discovery. Med. Res. Rev. 2018, 38, 1255–1294. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Lu, J.J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospect 2021, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Tommonaro, G. Curcumin and Cancer. Nutrients 2019, 11, 2376. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, M.; Corsini, E.; Rosini, M.; Racchi, M.; Lanni, C. Immunomodulators Inspired by Nature: A Review on Curcumin and Echinacea. Molecules 2018, 23, 2778. [Google Scholar] [CrossRef] [PubMed]
- Mbese, Z.; Khwaza, V.; Aderibigbe, B.A. Curcumin and Its Derivatives as Potential Therapeutic Agents in Prostate, Colon and Breast Cancers. Molecules 2019, 24, 4386. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Jitoe, A.; Isobe, J.; Nakatani, N.; Yonemori, S. Anti-Oxidative and Anti-Inflammatory Curcumin-Related Phenolics from Rhizomes of Curcuma Domestica. Phytochemistry 1993, 32, 1557–1560. [Google Scholar] [CrossRef]
- Sueth-Santiago, V.; Mendes-Silva, G.P.; Decoté-Ricardo, D.; De Lima, M.E.F. Curcumina, o Pó Dourado Do Açafrão-Da-Terra: Introspecções Sobre Química e Atividades Biológicas. Quim. Nova 2015, 38, 538–552. [Google Scholar]
- Lin, L.; Shi, Q.; Nyarko, A.K.; Bastow, K.F.; Wu, C.C.; Su, C.Y.; Shih, C.C.Y.; Lee, K.H. Antitumor Agents. 250. Design and Synthesis of New Curcumin Analogues as Potential Anti-Prostate Cancer Agents. J. Med. Chem. 2006, 49, 3963–3972. [Google Scholar] [CrossRef]
- Robinson, T.P.; Ehlers, T.; Hubbard IV, R.B.; Bai, X.; Arbiser, J.L.; Goldsmith, D.J.; Bowen, J.P. Design, Synthesis, and Biological Evaluation of Angiogenesis Inhibitors: Aromatic Enone and Dienone Analogues of Curcumin. Bioorg. Med. Chem. Lett. 2003, 13, 115–117. [Google Scholar] [CrossRef]
- Hossain, M.; Das, U.; Dimmock, J.R. Recent Advances in α,β-Unsaturated Carbonyl Compounds as Mitochondrial Toxins. Eur. J. Med. Chem. 2019, 183, 111687. [Google Scholar] [CrossRef]
- He, G.; Feng, C.; Vinothkumar, R.; Chen, W.; Dai, X.; Chen, X.; Ye, Q.; Qiu, C.; Zhou, H.; Wang, Y.; et al. Curcumin Analog EF24 Induces Apoptosis via ROS-Dependent Mitochondrial Dysfunction in Human Colorectal Cancer Cells. Cancer Chemother. Pharmacol. 2016, 78, 1151–1161. [Google Scholar] [CrossRef]
- Shin, S.H.; Kim, I.; Lee, J.E.; Lee, M.; Park, J.W. Loss of EGR3 Is an Independent Risk Factor for Metastatic Progression in Prostate Cancer. Oncogene 2020, 39, 5839–5854. [Google Scholar] [CrossRef]
- Hossain, M.; Das, S.; Das, U.; Doroudi, A.; Zhu, J.; Dimmock, J.R. Novel Hybrid Molecules of 3,5-Bis(Benzylidene)-4-Piperidones and Dichloroacetic Acid Which Demonstrate Potent Tumour-Selective Cytotoxicity. Bioorg. Med. Chem. Lett. 2020, 30, 126878. [Google Scholar] [CrossRef]
- WHO. Cancer Today. Available online: https://gco.iarc.fr/today/online-analysis-table?v=2020&mode=cancer&mode_population=continents&population=900&populations=900&key=asr&sex=0&cancer=39&type=1&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&group_cancer=1&i (accessed on 2 March 2022).
- Ko, C.C.; Yeh, L.R.; Kuo, Y.T.; Chen, J.H. Imaging Biomarkers for Evaluating Tumor Response: RECIST and Beyond. Biomark. Res. 2021, 9, 52. [Google Scholar] [CrossRef]
- Wang, N.; Feng, T.; Liu, X.; Liu, Q. Curcumin Inhibits Migration and Invasion of Non-Small Cell Lung Cancer Cells through up-Regulation of MiR-206 and Suppression of PI3K/AKT/MTOR Signaling Pathway. Acta Pharm. 2020, 70, 399–409. [Google Scholar] [CrossRef]
- Shen, H.; Shen, J.; Pan, H.; Xu, L.; Sheng, H.; Liu, B.; Yao, M. Curcumin Analog B14 Has High Bioavailability and Enhances the Effect of Anti-breast Cancer Cells in Vitro and in Vivo. Cancer Sci. 2021, 112, 815. [Google Scholar] [CrossRef]
- Noh, D.Y.; Han, W.; Toi, M. Translational Research in Breast Cancer; Springer: Berlin/Heidelberg, Germany, 2021; Volume 1187. [Google Scholar] [CrossRef]
- Park, M.; Kim, D.; Ko, S.; Kim, A.; Mo, K.; Yoon, H. Breast Cancer Metastasis: Mechanisms and Therapeutic Implications. Int. J. Mol. Sci. 2022, 23, 6806. [Google Scholar] [CrossRef]
- Morita, K.; Okamura, T.; Sumitomo, S.; Iwasaki, Y.; Fujio, K.; Yamamoto, K. Emerging Roles of Egr2 and Egr3 in the Control of Systemic Autoimmunity. Rheumatology 2016, 55, ii76–ii81. [Google Scholar] [CrossRef]
- Suzuki, T.; Inoue, A.; Miki, Y.; Moriya, T.; Akahira, J.I.; Ishida, T.; Hirakawa, H.; Yamaguchi, Y.; Hayashi, S.I.; Sasano, H. Early Growth Responsive Gene 3 in Human Breast Carcinoma: A Regulator of Estrogen-Meditated Invasion and a Potent Prognostic Factor. Endocr. Relat. Cancer 2007, 14, 279–292. [Google Scholar] [CrossRef]
- Li, X.Z.; Tu, Y.J.; Zhou, T.; Zhang, J.B.; Xiao, R.W.; Yang, D.W.; Zhang, P.F.; You, P.T.; Zheng, X.H. MicroRNA-483-5p Predicts Poor Prognosis and Promotes Cancer Metastasis by Targeting EGR3 in Nasopharyngeal Carcinoma. Front. Oncol. 2021, 11, 720835. [Google Scholar] [CrossRef]
- Wu, J.; Huang, Y.; Zhou, X.; Xiang, Z.; Yang, Z.; Meng, D.; Wu, D.; Zhang, J.; Yang, J. ATF3 and Its Emerging Role in Atherosclerosis: A Narrative Review. Cardiovasc. Diagn. Ther. 2022, 12, 926–942. [Google Scholar] [CrossRef]
- Liu, S.; Shi, J.; Wang, L.; Huang, Y.; Zhao, B.; Ding, H.; Liu, Y.; Wang, W.; Chen, Z.; Yang, J. Loss of EMP1 Promotes the Metastasis of Human Bladder Cancer Cells by Promoting Migration and Conferring Resistance to Ferroptosis through Activation of PPAR Gamma Signaling. Free Radic. Biol. Med. 2022, 189, 42–57. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Wu, H.; Wang, H.; Yao, L.; Deng, Z.; Zhou, Y. EMP1 Regulates Cell Proliferation, Migration, and Stemness in Gliomas through PI3K-AKT Signaling and CD44. J. Cell Biochem. 2019, 120, 17142–17150. [Google Scholar] [CrossRef]
- Kodama, S.; Negishi, M. Pregnane X Receptor PXR Activates the GADD45β Gene, Eliciting the P38 MAPK Signal and Cell Migration. J. Biol. Chem. 2011, 286, 3570. [Google Scholar] [CrossRef]
- Xiao, Y.; Li, Y.; Shi, D.; Wang, X.; Dai, S.; Yang, M.; Kong, L.; Chen, B.; Huang, X.; Lin, C.; et al. MEX3C-Mediated Decay of SOCS3 MRNA Promotes JAK2/STAT3 Signaling to Facilitate Metastasis in Hepatocellular Carcinoma. Cancer Res. 2022, 82, 4191–4205. [Google Scholar] [CrossRef]
- Park, J.M.; Lee, T.H.; Kang, T.H. Roles of Tristetraprolin in Tumorigenesis. Int. J. Mol. Sci. 2018, 19, 3384. [Google Scholar] [CrossRef]
- Song, B.; Shen, S.; Fu, S.; Fu, J. HSPA6 and Its Role in Cancers and Other Diseases. Mol. Biol. Rep. 2022, 49, 10565–10577. [Google Scholar] [CrossRef]
- Shen, S.; Wei, C.; Fu, J. RNA-Sequencing Reveals Heat Shock 70-KDa Protein 6 (HSPA6) as a Novel Thymoquinone-Upregulated Gene That Inhibits Growth, Migration, and Invasion of Triple-Negative Breast Cancer Cells. Front. Oncol. 2021, 11, 667995. [Google Scholar] [CrossRef] [PubMed]
- Komoto, T.T.; Bitencourt, T.A.; Silva, G.; Beleboni, R.O.; Marins, M.; Fachin, A.L.; Komoto, T.T.; Bitencourt, T.A.; Silva, G.; Beleboni, R.O.; et al. Gene Expression Response of Trichophyton Rubrum during Coculture on Keratinocytes Exposed to Antifungal Agents. Evid. Based Complement. Altern. Med. 2015, 2015, 180535. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | |
|---|---|---|
| Forward 5′-3′ | Reverse 5′-3′ | |
| EGR3 | GACATCGGTCTGACCAACGAG | GGCGAACTTTCCCAAGTAGGT |
| ATF3 | GTTTGAGGATTTTGCTAACCTGAC | AGCTGCAATCTTATTTCTTTCTCGT |
| EMP1 | GCCAATGTCTGGTTGGTTTCC | GAGGGCATCTTCACTGGCATA |
| SOCS3 | GACCAGCGCCACTTCTTCA | CTGGATGCGCAGGTTCTTG |
| ZFP36 | GACTGAGCTATGTCGGACCTT | GAGTTCCGTCTTGTATTTGGGG |
| GADD45B | ATTGCAACATGACGCTGGAAGAGC | GGATGAGCGTGAAGTGGATT |
| GAPDH | GACCACAGTCCATGCCATCACT | TCCACCACCCTGTTGCTGTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishimura, F.G.; Sampaio, B.B.; do Couto, G.O.; da Silva, A.D.; da Silva, W.J.; Peronni, K.C.; Evangelista, A.F.; Hossain, M.; Dimmock, J.R.; Bandy, B.; et al. The Transcriptome of BT-20 Breast Cancer Cells Exposed to Curcumin Analog NC2603 Reveals a Relationship between EGR3 Gene Modulation and Cell Migration Inhibition. Molecules 2024, 29, 1366. https://doi.org/10.3390/molecules29061366
Nishimura FG, Sampaio BB, do Couto GO, da Silva AD, da Silva WJ, Peronni KC, Evangelista AF, Hossain M, Dimmock JR, Bandy B, et al. The Transcriptome of BT-20 Breast Cancer Cells Exposed to Curcumin Analog NC2603 Reveals a Relationship between EGR3 Gene Modulation and Cell Migration Inhibition. Molecules. 2024; 29(6):1366. https://doi.org/10.3390/molecules29061366
Chicago/Turabian StyleNishimura, Felipe Garcia, Beatriz Borsani Sampaio, Gabrielly Oliveira do Couto, Aryane Dias da Silva, Wanessa Julia da Silva, Kamila Chagas Peronni, Adriane Feijó Evangelista, Mohammad Hossain, Jonathan R. Dimmock, Brian Bandy, and et al. 2024. "The Transcriptome of BT-20 Breast Cancer Cells Exposed to Curcumin Analog NC2603 Reveals a Relationship between EGR3 Gene Modulation and Cell Migration Inhibition" Molecules 29, no. 6: 1366. https://doi.org/10.3390/molecules29061366
APA StyleNishimura, F. G., Sampaio, B. B., do Couto, G. O., da Silva, A. D., da Silva, W. J., Peronni, K. C., Evangelista, A. F., Hossain, M., Dimmock, J. R., Bandy, B., Beleboni, R. O., Marins, M., & Fachin, A. L. (2024). The Transcriptome of BT-20 Breast Cancer Cells Exposed to Curcumin Analog NC2603 Reveals a Relationship between EGR3 Gene Modulation and Cell Migration Inhibition. Molecules, 29(6), 1366. https://doi.org/10.3390/molecules29061366