

Anti-Tumor Activity and Mechanism of Silibinin Based on Network Pharmacology and Experimental Verification
 , , ,
, , , 
Abstract
1. Introduction
2. Results
2.1. Network Pharmacology Analysis
2.1.1. Target Proteins of Silibinin
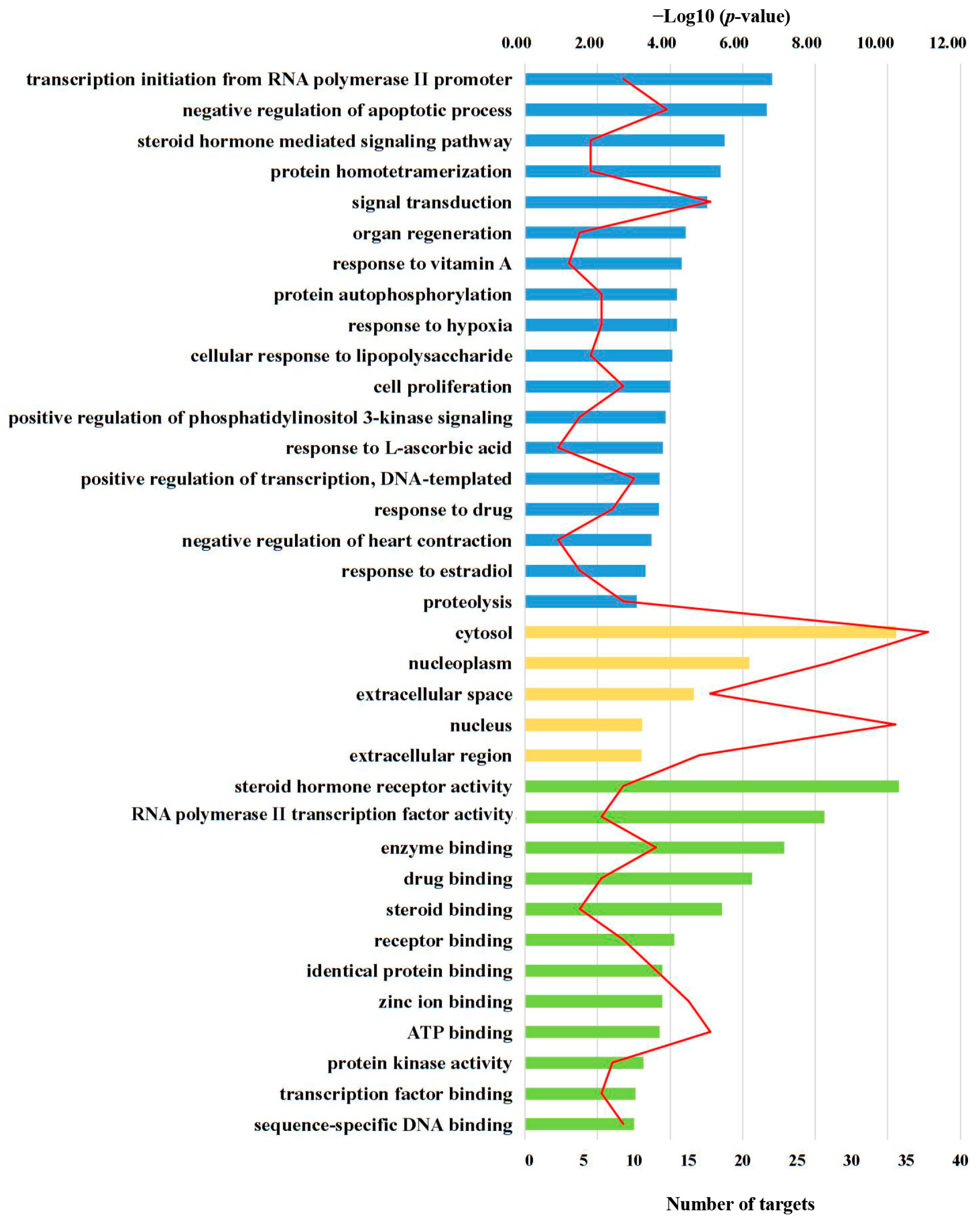
2.1.2. GO Enrichment Analysis for Targets
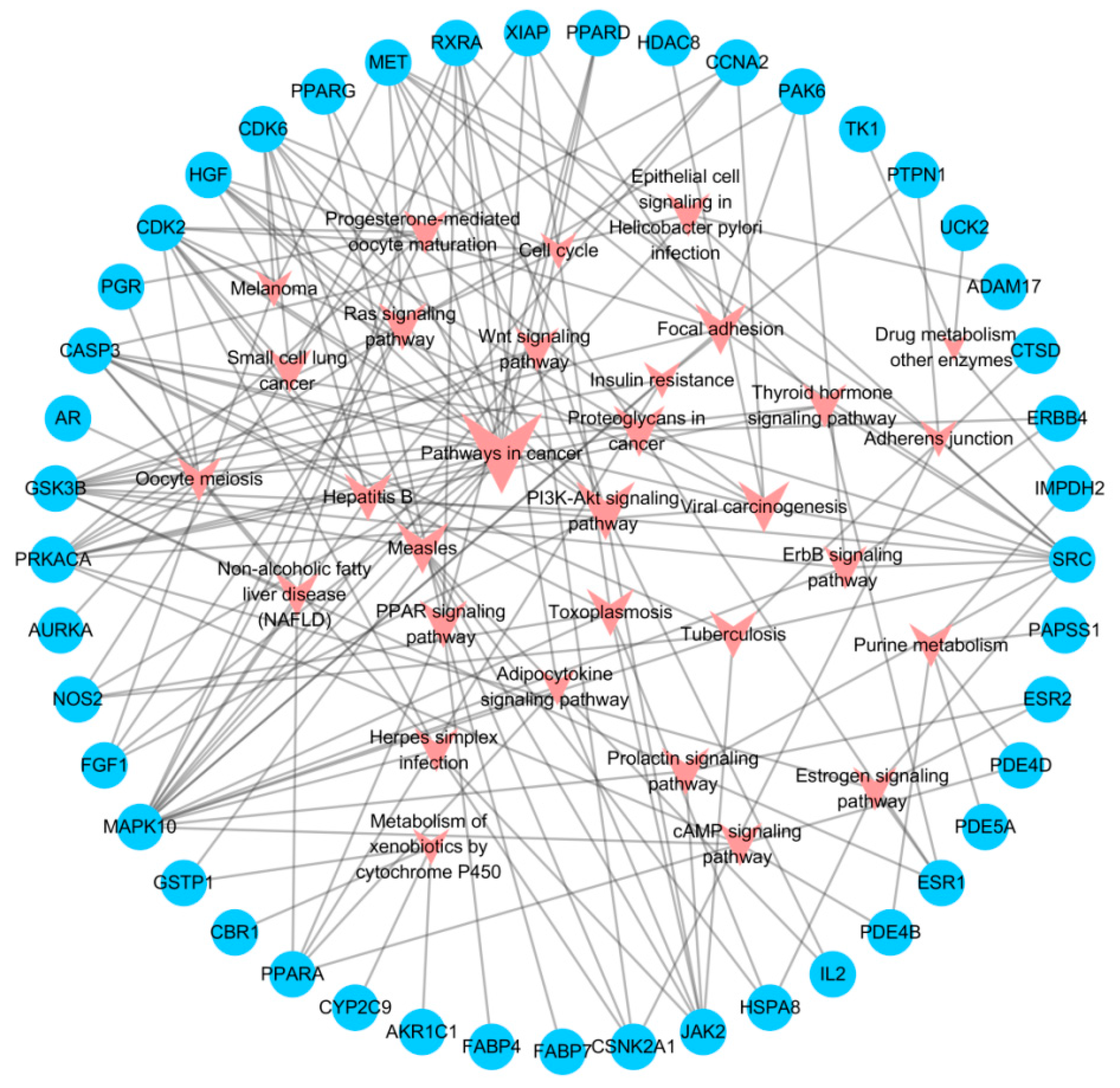
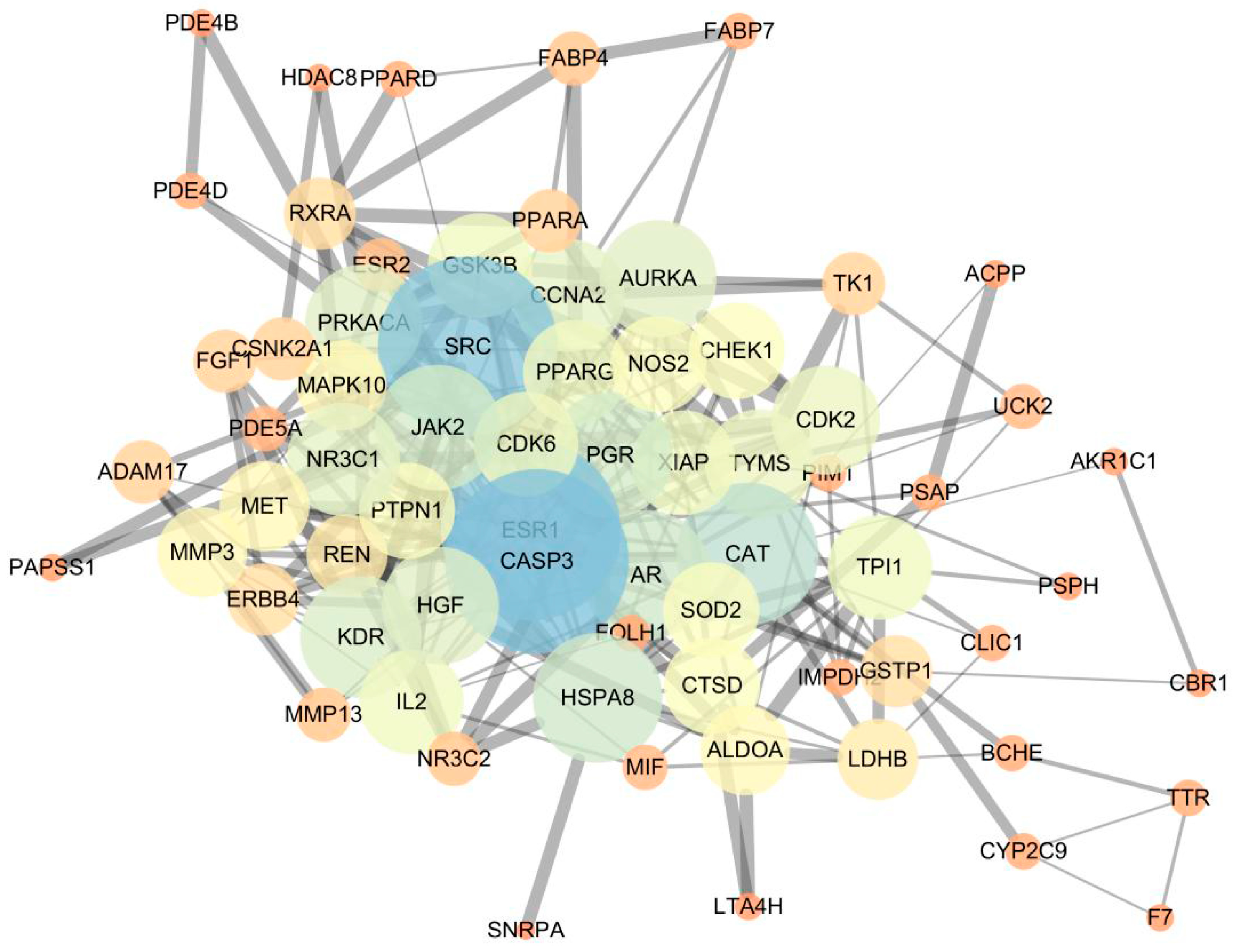
2.1.3. Network Construction and Mechanism Analysis
2.2. Silibinin Inhibited ACC2 Cell Viability and Proliferation
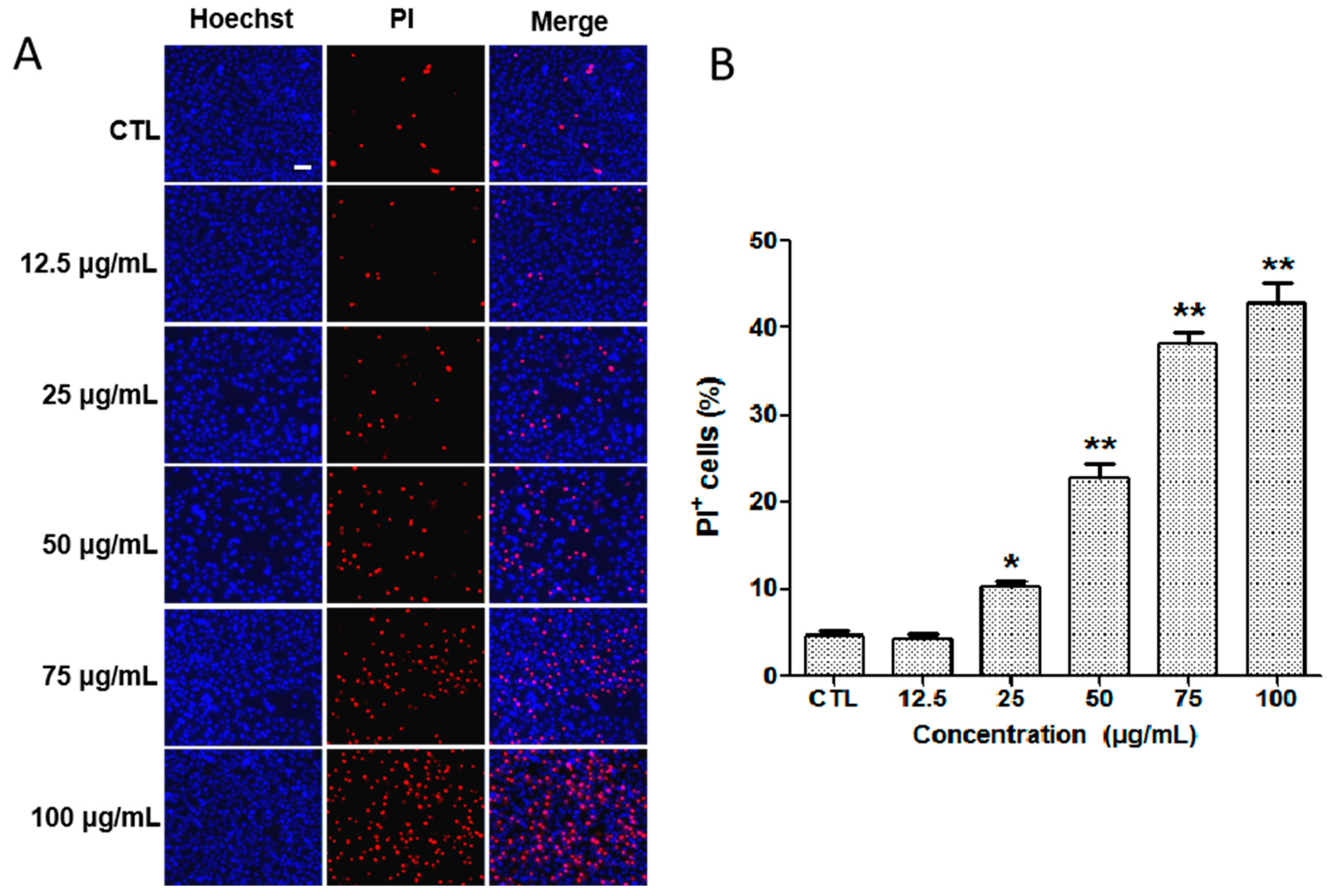
2.3. Detection of Apoptosis by Hoechst 33342/PI Staining In Vitro
2.4. Cell Migration Effect of Silibinin In Vitro
2.5. Effect of Silibinin on Tumor-Related Targets In Vitro
2.6. Effect of Silibinin on Tumor-Bearing Mice
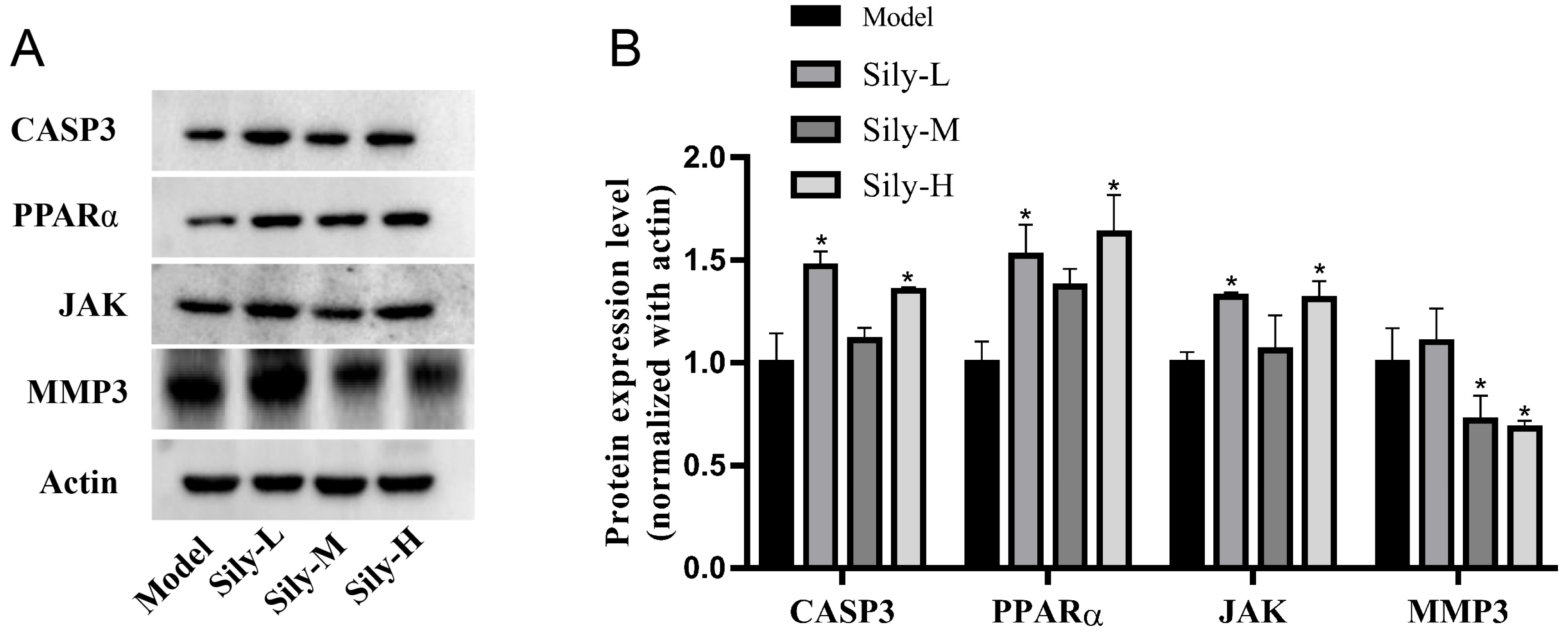
2.7. Effect of Silibinin on Tumor-Related Targets In Vivo
3. Material and Methods
3.1. Network Pharmacology Analysis
3.1.1. Target Fishing
3.1.2. KEGG Pathway and Gene Ontology (GO) Terms Analysis
3.1.3. Network Construction and Analysis
3.2. Chemicals and Reagents
3.3. Cell Culture and Treatments
3.4. MTT Assay
3.5. Cell Proliferation
3.6. Cell Apoptosis
3.7. Wound Closure Assay
3.8. Animals and Treatment
3.9. Histopathological Analysis
3.10. Western Blot Analysis
3.11. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xing, X.; Wang, X.; He, W. Advances in research on tumor immunotherapy and its drug development. J. China Pharmaceut. 2021, U52, 10–19. [Google Scholar]
- Jiang, Z. Research progress on antitumor effect of berberine. World Latest Med. Inf. 2021, 21, 27–28. [Google Scholar]
- Liu, S.H.; Chen, P.S.; Huang, C.C.; Hung, Y.T.; Lee, M.Y.; Lin, W.H.; Lin, Y.C.; Lee, A.Y. Unlocking the mystery of the therapeutic effects of Chinese medicine on cancer. Front. Pharmacol. 2020, 11, 601785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Mao-Mao; Shao, M.M.; Wang, M.C. Therapeutic potential of natural flavonoids in pulmonary arterial hypertension: A review. Phytomedicine 2024, 128, 155535. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Li, X.; He, L.; Zheng, Y.; Lu, H.; Li, J.; Zhong, L.; Tong, R.; Jiang, Z.; Shi, J.; et al. Antidiabetic Potential of Flavonoids from Traditional Chinese Medicine: A Review. Am. J. Chin. Med. 2019, 47, 933–957. [Google Scholar] [CrossRef] [PubMed]
- Nanjing University of Chinese Medicine. Dictionary of Traditional Chinese Medicine; Shanghai Science and Technology Press: Shanghai, China, 2005; p. 729. [Google Scholar]
- Liang, Z.; Yang, Y.; Wang, H.; Yi, W.; Yan, X.; Yan, J.; Li, Y.; Feng, Y.; Yu, S.; Yang, J.; et al. Inhibition of SIRT1 signaling sensitizes the antitumor activity of silybin against human lung adenocarcinoma cells in vitro and in vivo. Mol. Cancer Ther. 2014, 13, 1860–1872. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, Y. The clinical application of Silybum marianum (L.). Gaertn. Med. J. 2019, 13, 21. [Google Scholar]
- Tao, R.T.; Liu, P.H.; Ran, L.T. Effect of silibinin on proliferation and invasion in oral epithelial KB cell line. Mod. Oncol. 2020, 28, 1629–1633. [Google Scholar]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of People’s Republic of China; Medical Science Press of China: Beijing, China, 2020. [Google Scholar]
- Deep, G.; Agarwal, R. Antimetastatic efficacy of silibinin: Molecular mechanisms and therapeutic potential against cancer. Cancer Metastasis Rev. 2010, 29, 447–463. [Google Scholar] [CrossRef] [PubMed]
- Liu, X. The pharmacological effects and clinical evaluation of silymarin. Guide China Med. 2012, 14, 66–67. [Google Scholar]
- Flaig, T.W.; Glodé, M.; Gustafson, D.; Bokhoven, A.; Tao, Y.; Wilson, S.; Su, L.; Li, Y.; Harrison, G.; Agarwal, R.; et al. A study of high-dose oral silybin-phytosome followed by prostatectomy in patients with localized prostate cancer. Prostate 2010, 70, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liang, W.C.; Pan, W.L.; Law, W.K.; Hu, J.S.; Ip, D.T.M.; Waye, M.M.Y.; Ng, T.B.; Wan, D.C.C. Silibinin, a novel chemokine receptor type 4 antagonist, inhibits chemokine ligand 12-induced migration in breast cancer cells. Phytomedicine 2014, 21, 1310–1317. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Lee, H.J.; Rho, J.K.; Chung, S.Y.; Lee, C.G.; Yang, K.; Kim, S.H.; Lee, M.; Shin, I.S.; Kim, J.S. The ameliorative effect of silibinin against radiation-induced lung injury: Protection of normal tissue without decreasing therapeutic efficacy in lung cancer. BMC Pulm. Med. 2015, 15, 68. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.Q.; Yue, S.J.; Tang, Y.P.; Chen, Y.Y.; Zhou, G.S.; Zhang, J.; Zhu, Z.H.; Liu, P.; Duan, J.A. A network pharmacology approach to investigate the blood enriching mechanism of Dangguibuxue Decoction. J. Ethnopharmacol. 2019, 235, 227–242. [Google Scholar] [CrossRef]
- Zhao, M.; Chen, Y.; Wang, C.; Xiao, W.; Chen, S.; Zhang, S.; Yang, L.; Li, Y. Systems pharmacology dissection of multi-scale mechanisms of action of Huo-Xiang-Zheng-Qi Formula for the treatment of gastrointestinal diseases. Front. Pharmacol. 2019, 9, 1448. [Google Scholar] [CrossRef]
- Wu, B.; Li, C.; Kan, H.; Zhang, Y.; Rao, X.; Liu, Y.; Zhao, P. Hypolipidemic and Antithrombotic Effect of 6’-O-Caffeoylarbutin from Vaccinium dunalianum Based on Zebrafish Model, Network Pharmacology, and Molecular Docking. Molecules 2024, 29, 780. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Li, Y.; Zhao, J.; Wang, Y.; Li, Y.; Wang, Y. The mechanism of triptolide in the treatment of connective tissue disease-related interstitial lung disease based on network pharmacology and molecular docking. Ann. Med. 2022, 54, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.J.; Yan, X.L.; Dong, B.S.; Yang, W.N.; Su, S.B.; Zhang, H. A network pharmacology approach to investigating the mechanism of Tanshinone IIA for the treatment of liver fibrosis. J. Ethnopharmacol. 2020, 253, 112689. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- DeSouza, N.; Zhou, M.; Shan, Y. Chronic Myeloid Leukemia: Methods and Protocols; Springer: New York, NY, USA, 2016. [Google Scholar]
- Pereira, P.D.; Serra-Caetano, A.; Cabrita, M.; Bekman, E.; Braga, J.; Rino, J.; Santus, R.; Filipe, P.L.; Sousa, A.E.; Ferreira, J.A. Quantification of cell cycle kinetics by EdU (5-ethynyl-2′-deoxyuridine)-coupled-fluorescence-intensity analysis. Oncotarget 2017, 8, 40514–40532. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zeng, F.; Zhang, M.; Pan, Z.; Chen, Y.; Zeng, Y.; Xu, Y.; Xu, Q.; Huang, Y. Green synthesis of hyaluronic acid-based silver nanoparticles and their enhanced delivery to CD44+ cancer cells. RSC Adv. 2015, 5, 43733–43740. [Google Scholar] [CrossRef]
- Chung, C.Y.; Murphy-Ullrich, J.E.; Erickson, H.P. Mitogenesis, cell migration, and loss of focal adhesions induced by tenascin-C interacting with its cell surface receptor, annexin II. Mol. Biol. Cell 1996, 7, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Win, N.N.; Awale, S.; Esumi, H.; Tezuka, Y.; Kadota, S. Novel anticancer agents, kayeassamins A and B from the flower of Kayea assamica of Myanmar. Bioorg. Med. Chem. Lett. 2008, 18, 4688–4691. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic. Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef] [PubMed]
- Keiser, M.J.; Roth, B.L.; Armbruster, B.N.; Ernsberger, P.; Irwin, J.J.; Shoichet, B.K. Relating protein pharmacology by ligand chemistry. Nat. Biotechnol. 2007, 25, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.; Szklarczyk, D.; Franceschini, A.; Mering, C.; Jensen, L.J.; Bork, P. STITCH 3: Zooming in on protein–chemical interactions. Nucleic. Acids Res. 2012, 40, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.D.; Gao, J.; Luo, M. Identification of key pathways and hub genes in basal-like breast cancer using bioinformatics analysis. Onco Targets Ther. 2019, 12, 1319–1331. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, S.; Kulkarni, P.; Thummuri, D.; Jeengar, M.K.; Naidu, V.G.M.; Alvala, M.; Redddy, G.B.; Ramakrishna, S. Piperlongumine, an alkaloid causes inhibition of PI3K/Akt/mTOR signaling axis to induce caspase-dependent apoptosis in human triple-negative breast cancer cells. Apoptosis 2014, 19, 1148–1164. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, X.; Li, X.; Stoika, R.; Wang, X.; Lin, H.; Ma, Y.; Wang, R.; Liu, K. Synthesis of hydrophobically modified berberine derivatives with high anticancer activity through modulation of the MAPK pathway. New J. Chem. 2020, 44, 14024–14034. [Google Scholar] [CrossRef]
- Zhang, M.Q.; Ren, X.; Zhao, Q.; Yue, S.J.; Fu, X.M.; Li, X.; Chen, K.X.; Guo, Y.W.; Shao, C.L.; Wang, C.Y. Hepatoprotective effects of total phenylethanoid glycosides from Acanthus ilicifolius L. against carbon tetrachloride-induced hepatotoxicity. J. Ethnopharmacol. 2020, 256, 112795. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.; Bhatia, N.; Condon, M.S.; Bosland, M.C.; Agarwal, C.; Agarwal, R. Antiproliferative and apoptotic effects of silibinin in rat prostate cancer cells. Prostate 2002, 53, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Zhu, Y.; Lu, Q.; Zhang, J.; Ge, Z.; Zheng, S. Silymarin causes caspases activation and apoptosis in K562 leukemia cells through inactivation of Akt pathway. Toxicology 2006, 227, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Gao, Y.; Zhang, L.; Zeng, J.; He, D.; Sun, Y. Silibinin inhibits cell growth and induces apoptosis by caspase activation, down-regulating survivin and blocking EGFR-ERK activation in renal cell carcinoma. Cancer Lett. 2008, 272, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.K.; Singh, R.P.; Agarwal, C.; Chan, D.C.; Agarwal, R. Silibinin strongly synergizes human prostate carcinoma DU145 cells to doxorubicin-induced growth inhibition, G2/M arrest, and apoptosis. Clin. Cancer Res. 2002, 8, 3512–3519. [Google Scholar] [PubMed]
- Dhanalakshmi, S.; Agarwal, P.; Glode, L.M.; Agarwal, R. Silibinin sensitizes human prostate carcinoma DU145 cells to cisplatin- and carboplatin-induced growth inhibition and apoptotic death. Int. J. Cancer 2003, 106, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Peng, J.; Liu, W.; He, X.; Cui, L.; Chen, X.; Yang, M.; Liu, H.; Liu, S.; Wang, H. Elevated cleaved caspase-3 is associated with shortened overall survival in several cancer types. Int. J. Clin. Exp. Pathol. 2014, 7, 5057–5070. [Google Scholar] [PubMed]
- Tsukamoto, T.; Kajiwara, K.; Nada, S.; Okada, M. Src mediates TGF-beta-induced intraocular pressure elevation in glaucoma. J. Cell Physiol. 2019, 234, 1730–1744. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.; Kim, S.J.; Seelmann, A.; Veit, B.; Shepard, B.; Im, E.; Rhee, S.H. Src family kinase tyrosine phosphorylates toll-like receptor 4 to dissociate MyD88 and Mal/Tirap, suppressing LPS-induced inflammatory responses. Biochem. Pharmacol. 2018, 147, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Feng, Z.; Li, Q.; Qi, Z.; Zhang, Y. Inhibition of ROS-mediated activation Src-MAPK/AKT signaling by orientin alleviates H2O2-induced apoptosis in PC12 cells. Drug Des. Devel. Ther. 2018, 12, 3973–3984. [Google Scholar] [CrossRef] [PubMed]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. Febs J. 2017, 278, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Warnecke-Eberz, U.; Metzger, R.; Holscher, A.H.; Drebber, U.; Bollschweiler, E. Diagnostic marker signature for esophageal cancer from transcriptome analysis. Tumor Biol. 2016, 37, 6349–6358. [Google Scholar] [CrossRef] [PubMed]
- Gobin, E.; Bagwell, K.; Wagner, J.; Mysona, D.; Sandirasegarane, S.; Smith, N.; Bai, S.; Sharma, A.; Schleifer, R.; She, J.X. A pan-cancer perspective of matrix metalloproteases (MMP) gene expression profile and their diagnostic/prognostic potential. BMC Cancer 2019, 19, 581. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Li, Z.; Ding, R.; Cheng, M.; Huang, H.; Liu, A.; Kang, M.; He, H.; Xu, Y.; Shao, J.; et al. Extensive serum biomarker analysis in patients with nasopharyngeal carcinoma. Cytokine 2019, 118, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Yang, Y. Identification and analysis of genes associated with head and neck squamous cell carcinoma by integrated bioinformatics methods. Mol. Genet. Genomic Med. 2019, 7, e857. [Google Scholar] [CrossRef] [PubMed]
- Routti, H.; Berg, M.K.; Lille-Langøy, R.; Øygarden, L.; Harju, M.; Dietz, R.; Sonne, C.; Goksøyr, A. Environmental contaminants modulate the transcriptional activity of polar bear (Ursus maritimus) and human peroxisome proliferator-activated receptor alpha (PPARA). Sci. Rep. 2019, 9, 6918. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Wagner, M.; Xiao, R.; Kim, K.H.; Feng, D.; Lazar, M.A.; Moore, D.D. Nutrient-sensing nuclear receptors coordinate autophagy. Nature 2014, 516, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Pizzimenti, S.; Laurora, S.; Briatore, F.; Ferretti, C.; Dianzani, M.U.; Barrera, G. Synergistic effect of 4-hydroxynonenal and PPAR ligands in controlling human leukemic cell growth and differentiation. Free Radic. Biol. Med. 2002, 32, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Nie, S. Advances in study on JAK/STAT3 signaling pathway and its role in gastrointestinal diseases. Chin. J. Gastroenterol. 2014, 19, 301–304. [Google Scholar]
- Tang, L.; Zhu, S.; Peng, W.; Yin, X.; Tan, C.; Yang, Y. Epigenetic identification of mitogen-activated protein kinase 10 as a functional tumor suppressor and clinical significance for hepatocellular carcinoma. Peer J. 2021, 9, e10810. [Google Scholar] [CrossRef] [PubMed]
- Zi, X.; Agarwal, R. Silibinin decreases prostate-specific antigen with cell growth inhibition via G1 arrest, leading to differentiation of prostate carcinoma cells: Implications for prostate cancer intervention. Proc. Natl. Acad. Sci. USA 1999, 96, 7490–7495. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Target | Uniprot ID | Target Name |
|---|---|---|---|
| T1 | cGMP-specific 3,5-cyclic phosphodiesterase | T94879 | PDE5A |
| T2 | Estrogen receptor | T89534 | ESR1 |
| T3 | Stromelysin-1 | T86702 | MMP3 |
| T4 | Cholinesterase | T99799 | BCHE |
| T5 | Mitogen-activated protein kinase 10 | T85421 | MAPK10 |
| T6 | Estrogen receptor beta | T80896 | ESR2 |
| T7 | Transthyretin | T86462 | TTR |
| T8 | Proto-oncogene serine/threonine-protein kinase Pim-1 | T50594 | PIM1 |
| T9 | U1 small nuclear ribonucleoprotein A | P09012 | SNRPA |
| T10 | Glutathione S-transferase P | T21669 | GSTP1 |
| T11 | Tyrosine-protein phosphatase non-receptor type 1 | T89529 | PTPN1 |
| T12 | Peroxisome proliferator-activated receptor gamma | P37231 | PPARG |
| T13 | Cathepsin D | T67102 | CTSD |
| T14 | Macrophage migration inhibitory factor | T39977 | MIF |
| T15 | Cell division protein kinase 2 | P24941 | CDK2 |
| T16 | Serine/threonine-protein kinase Chk1 | T62449 | CHK1 |
| T17 | Mineralocorticoid receptor | T72168 | MR |
| T18 | Leukotriene A-4 hydrolase | T03691 | LTA4H |
| T19 | Progesterone receptor | T22939 | PGR |
| T20 | Cyclin-A2 | T58470 | CCNA2 |
| T21 | cAMP-specific 3,5-cyclic phosphodiesterase 4B | T10265 | PDE4B |
| T22 | Glutamate carboxypeptidase 2 | T97071 | FOLH1 |
| T23 | Collagenase 3 | T34296 | MMP13 |
| T24 | Phosphoserine phosphatase | P78330 | PSPH |
| T25 | Peroxisome proliferator-activated receptor delta | Q03181 | PPARD |
| T26 | Proto-oncogene tyrosine-protein kinase Src | T85943 | SRC |
| T27 | Thymidylate synthase | T98397 | TYMS |
| T28 | Androgen receptor | T11211 | AR |
| T29 | Cell division protein kinase 6 | T89361 | CDK6 |
| T30 | Aldo-keto reductase family 1 member C1 | Q04828 | AKR1C1 |
| T31 | cAMP-specific 3,5-cyclic phosphodiesterase 4D | T02001 | PDE4D |
| T32 | Casein kinase II subunit alpha | P68400 | CSNK2A1 |
| T33 | Chloride intracellular channel protein 1 | O00299 | CLIC1 |
| T34 | Heparin-binding growth factor 1 | T18639 | FGF1 |
| T35 | Glycogen synthase kinase-3 beta | T70977 | GSK3B |
| T36 | cAMP-dependent protein kinase catalytic subunit alpha | P17612 | PRKACA |
| T37 | Carbonyl reductase [NADPH] 1 | T70518 | CBR1 |
| T38 | Inosine-5-monophosphate dehydrogenase 2 | T89360 | IMPDH2 |
| T39 | L-lactate dehydrogenase B chain | P07195 | LDHB |
| T40 | Heat shock cognate 71 kDa protein | P11142 | HSPA8 |
| T41 | Superoxide dismutase [Mn], mitochondrial | P04179 | SOD2 |
| T42 | Retinoic acid receptor RXR-alpha | T13726 | RXRA |
| T43 | Caspase-3 | T57943 | CASP3 |
| T44 | Vascular endothelial growth factor receptor 2 | P35968 | VEGFR2 |
| T45 | Coagulation factor VII | T43332 | F7 |
| T46 | Cytochrome P450 2C9 | T19244 | CYP2C9 |
| T47 | Triosephosphate isomerase | T59130 | TPI |
| T48 | Peroxisome proliferator-activated receptor alpha | T86591 | PPARα |
| T49 | Catalase | T01597 | CAT |
| T50 | Hepatocyte growth factor receptor | T40474 | MET |
| T51 | Serine/threonine-protein kinase 6 | O14965 | AURKA |
| T52 | Prostatic acid phosphatase | T93283 | ACPP |
| T53 | Serine/threonine-protein kinase PAK 6 | Q9NQU5 | PAK6 |
| T54 | Disintegrin and metalloproteinase domain-containing protein 17 | P78536 | ADAM17 |
| T55 | Thymidine kinase, cytosolic | P04183 | TK1 |
| T56 | Protein-glutamine gamma-glutamyltransferase E | Q08188 | TGM3 |
| T57 | Histone deacetylase 8 | T28887 | HDAC8 |
| T58 | Receptor tyrosine-protein kinase erbB-4 | T92057 | ELNE |
| T59 | Fructose-bisphosphate aldolase A | P04075 | ERBB4 |
| T60 | Uridine-cytidine kinase 2 | Q9BZX2 | ALDOA |
| T61 | Fatty acid-binding protein, adipocyte | T07217 | UCK2 |
| T62 | Proactivator polypeptide | P07602 | FABP4 |
| T63 | Tyrosine-protein kinase JAK2 | T87554 | PSAP |
| T64 | Hepatocyte growth factor | P14210 | JAK2 |
| T65 | Renin | P00797 | HGF |
| T66 | Fatty acid-binding protein, brain | O15540 | REN |
| T67 | Bifunctional 3-phosphoadenosine 5-phosphosulfate synthetase 1 | O43252 | FABP7 |
| T68 | Baculoviral IAP repeat-containing protein 4 | P98170 | PAPSS1 |
| T69 | Endoplasmic reticulum mannosyl-oligosaccharide 1,2-alpha-mannosidase | Q9UKM7 | XIAP |
| T70 | Nitric oxide synthase, inducible | T02703 | MAN1B1 |
| T71 | Interleukin-2 | T61698 | NOS2 |
| T72 | Glucocorticoid receptor | P04150 | IL2 |
| T73 | Angiopoietin-1 receptor | Q02763 | NR3C1 |
| T74 | NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 4-like 2 | Q9NRX3 | NDUFA4L2 |
| T75 | Prostaglandin D2 receptor | Q13258 | PGD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Wang, D.; Yang, X.; Liu, C.; Li, X.; Zhang, X.; Liu, K.; Zhang, Y.; Zhang, M.; Wang, C.; et al. Anti-Tumor Activity and Mechanism of Silibinin Based on Network Pharmacology and Experimental Verification. Molecules 2024, 29, 1901. https://doi.org/10.3390/molecules29081901
Li P, Wang D, Yang X, Liu C, Li X, Zhang X, Liu K, Zhang Y, Zhang M, Wang C, et al. Anti-Tumor Activity and Mechanism of Silibinin Based on Network Pharmacology and Experimental Verification. Molecules. 2024; 29(8):1901. https://doi.org/10.3390/molecules29081901
Chicago/Turabian StyleLi, Peihai, Dexu Wang, Xueliang Yang, Changyu Liu, Xiaobin Li, Xuanming Zhang, Kechun Liu, Yun Zhang, Mengqi Zhang, Changyun Wang, and et al. 2024. "Anti-Tumor Activity and Mechanism of Silibinin Based on Network Pharmacology and Experimental Verification" Molecules 29, no. 8: 1901. https://doi.org/10.3390/molecules29081901
APA StyleLi, P., Wang, D., Yang, X., Liu, C., Li, X., Zhang, X., Liu, K., Zhang, Y., Zhang, M., Wang, C., & Wang, R. (2024). Anti-Tumor Activity and Mechanism of Silibinin Based on Network Pharmacology and Experimental Verification. Molecules, 29(8), 1901. https://doi.org/10.3390/molecules29081901