The preparation of the mRNA template coding for the 10th fibronection type III domain (21 out of 94 amino acids were randomized) began with the
in vitro transcription from a PCR-DNA library (10
12 different sequences) according to the method described in reference [
5] (
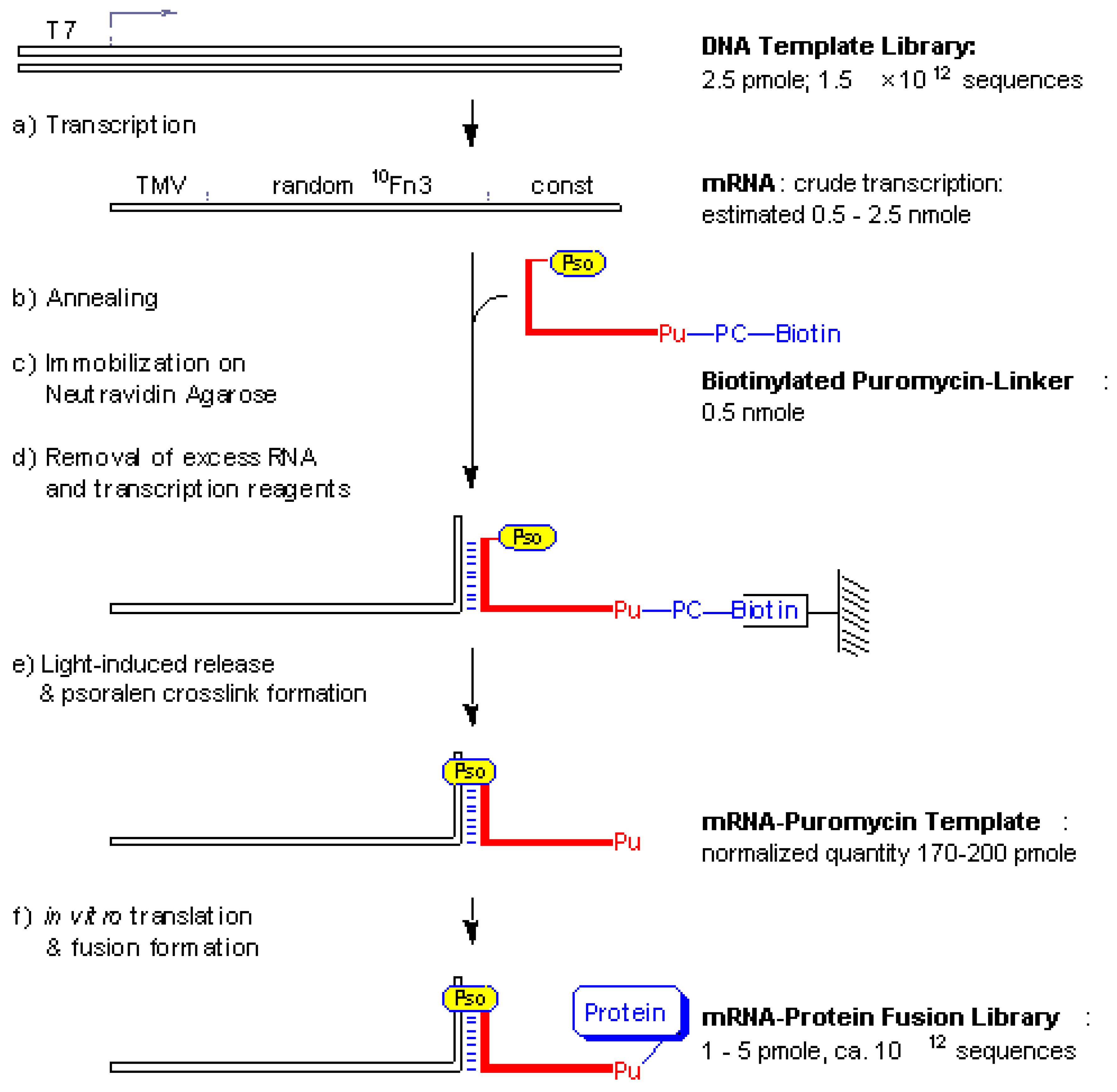
Fig. 1). The freshly transcribed mRNA was annealed to a limiting amount of puromycin-linker carrying a photo-cleavable biotin group for affinity purification on solid-phase (
Fig. 2). The hybridized mRNA template-linker complex was then incubated with neutravidin agarose beads followed by washing to remove excess mRNA and transcription buffer components. UV-irradiation of the beads led to psoralen interstrand-crosslink formation and simultaneous cleavage of the biotin-tag. A planned quantity of pure mRNA-puromycin template could be eluted from the solid phase. Polyacrylamide gel analysis indicated a ligation yield of > 80 % for the mRNA-puromycin template which was directly used for
in vitro translation and fusion formation in rabbit reticulocyte lysate to produce the mRNA-protein fusion library [
7].
Figure 1.
Schematic outline of mRNA-protein fusion synthesis. A double stranded DNA template library was
in vitro transcribed to produce the mRNA template (a) which was annealed with a limiting amount of biotinylated puromycin-linker (b) and subsequently captured on neutravidin beads (c) After a washing step to remove unbound mRNA (d) the beads were irradiated with UV light (e) Elution yielded the photo-ligated mRNA-puromycin template which was directly subjected to
in vitro translation to form the mRNA-protein fusions library (f) T7, transcription promoter sequence; TMV, a portion of the tobacco mosaic virus 5’-UTR with good initiation codon context; random
10Fn3, a sequence coding for a randomized 10th fibronection type III domain; const, a constant sequence which codes for the linker hybridization and crosslinking sites as detailed in
Fig. 2; Pu, puromycin; Pso, psoralen; PC-biotin, a photo-cleavable biotin-tag described in
Fig. 2.
Figure 1.
Schematic outline of mRNA-protein fusion synthesis. A double stranded DNA template library was
in vitro transcribed to produce the mRNA template (a) which was annealed with a limiting amount of biotinylated puromycin-linker (b) and subsequently captured on neutravidin beads (c) After a washing step to remove unbound mRNA (d) the beads were irradiated with UV light (e) Elution yielded the photo-ligated mRNA-puromycin template which was directly subjected to
in vitro translation to form the mRNA-protein fusions library (f) T7, transcription promoter sequence; TMV, a portion of the tobacco mosaic virus 5’-UTR with good initiation codon context; random
10Fn3, a sequence coding for a randomized 10th fibronection type III domain; const, a constant sequence which codes for the linker hybridization and crosslinking sites as detailed in
Fig. 2; Pu, puromycin; Pso, psoralen; PC-biotin, a photo-cleavable biotin-tag described in
Fig. 2.
Figure 2.
Sequence design of the mRNA template 3’-end and the puromycin-linker. The mRNA template 3'-constant region codes for a linker hybridization sequence, a UAA stop codon and a dA
18 tag sequence. The biotinylated puromycin-linker sequence shown in red carries a 5’- terminal psoralen moiety (Pso), followed by a hybridization region and a flexible tether with puromycin (Pu) at the 3’-end and carrying a photo-cleavable biotin-tag. A 10 base complementary region between mRNA 3’-end and the photo-linker was found to be sufficient to achieve efficient hybridization and photo-crosslink formation. The use of 2'-OMe RNA in the linker hybridization sequence was intended to increase the stability of the 10 base-pair duplex; also, it protected the mRNA from cleavage by any RNase H activity during translation in rabbit reticulocyte lysate. The hydroxymethyl trioxsalen moiety (psoralen) was tethered to the 5'- phosphate through a hexamethylene spacer [
8] (psoralen C6, Glen Research).
Figure 2.
Sequence design of the mRNA template 3’-end and the puromycin-linker. The mRNA template 3'-constant region codes for a linker hybridization sequence, a UAA stop codon and a dA
18 tag sequence. The biotinylated puromycin-linker sequence shown in red carries a 5’- terminal psoralen moiety (Pso), followed by a hybridization region and a flexible tether with puromycin (Pu) at the 3’-end and carrying a photo-cleavable biotin-tag. A 10 base complementary region between mRNA 3’-end and the photo-linker was found to be sufficient to achieve efficient hybridization and photo-crosslink formation. The use of 2'-OMe RNA in the linker hybridization sequence was intended to increase the stability of the 10 base-pair duplex; also, it protected the mRNA from cleavage by any RNase H activity during translation in rabbit reticulocyte lysate. The hydroxymethyl trioxsalen moiety (psoralen) was tethered to the 5'- phosphate through a hexamethylene spacer [
8] (psoralen C6, Glen Research).
We found the above protocol to be useful for efficient synthesis of mRNA-peptide and mRNA-protein fusion libraries from a number of template libraries different in length and complexity. Efficient mRNA affinity purification on neutravidin agarose carrying the biotinylated puromycin-linker replaced the laborious mRNA gel purification step used in previous protocols [
3,
5]. Crosslink-formation of the hybridized mRNA on solid-phase and simultaneous biotin-tag cleavage yielded pure mRNA-puromycin template for direct use in an
in vitro translation system. The ease with which the amount of synthesized mRNA-puromycin template can be controlled by hybridization with limiting amounts of linker allowed us to keep the reaction volumes and stoichiometry of the subsequent fusion preparation synthesis constant and independent from the yield of the preceding transcription reaction. This feature of the present protocol should prove most useful in an automated
in vitro protein evolution process which involves repetitive cycles of fusion library synthesis and selection and where manual quantification and adjustments only will lead to a time-lag in the production process.

{kind=link}
{kind=link}

