Exochelin Production in Mycobacterium neoaurum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
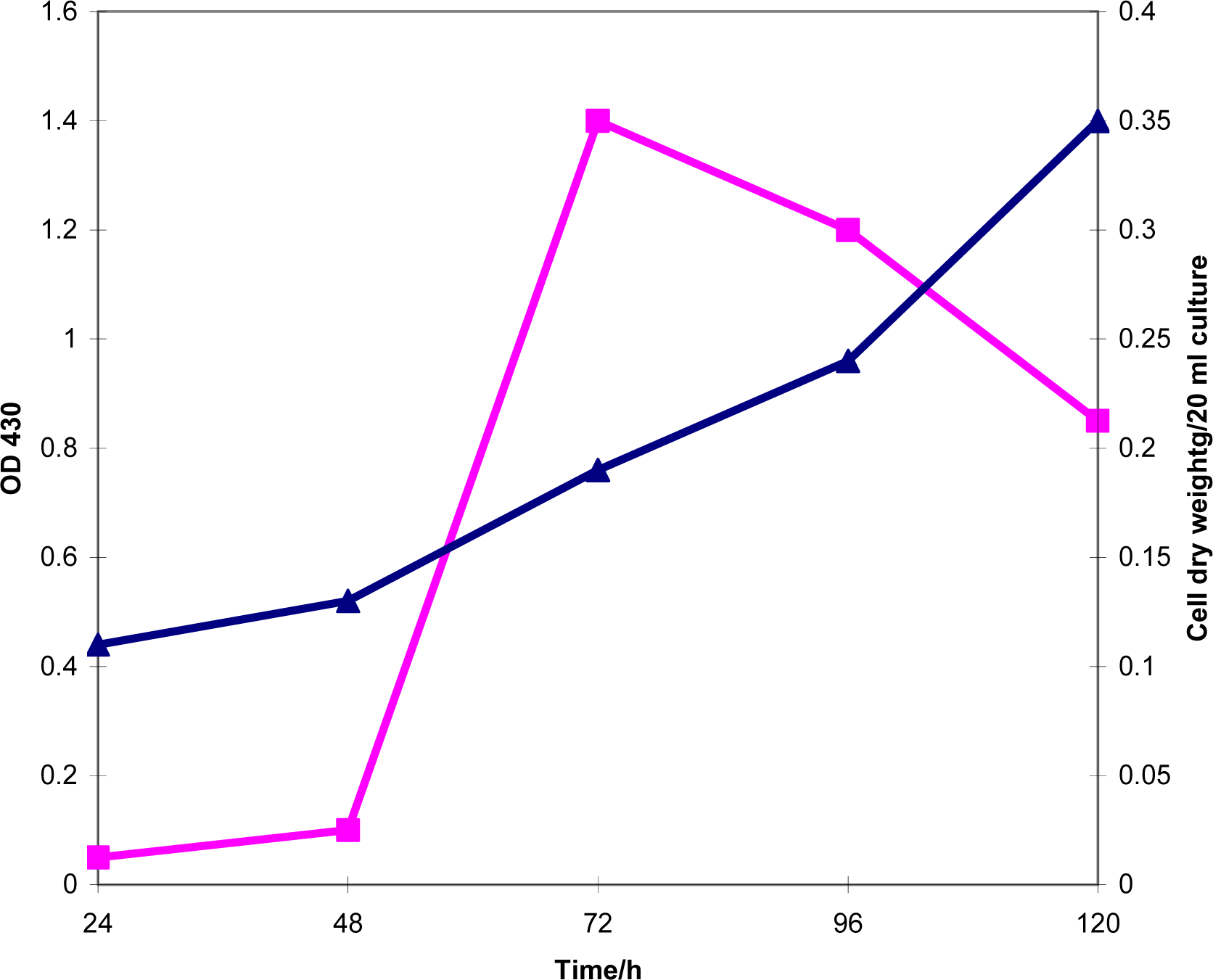
2.1. Production of exochelin in iron-deficiency, zinc-deficiency and iron-sufficiency conditions
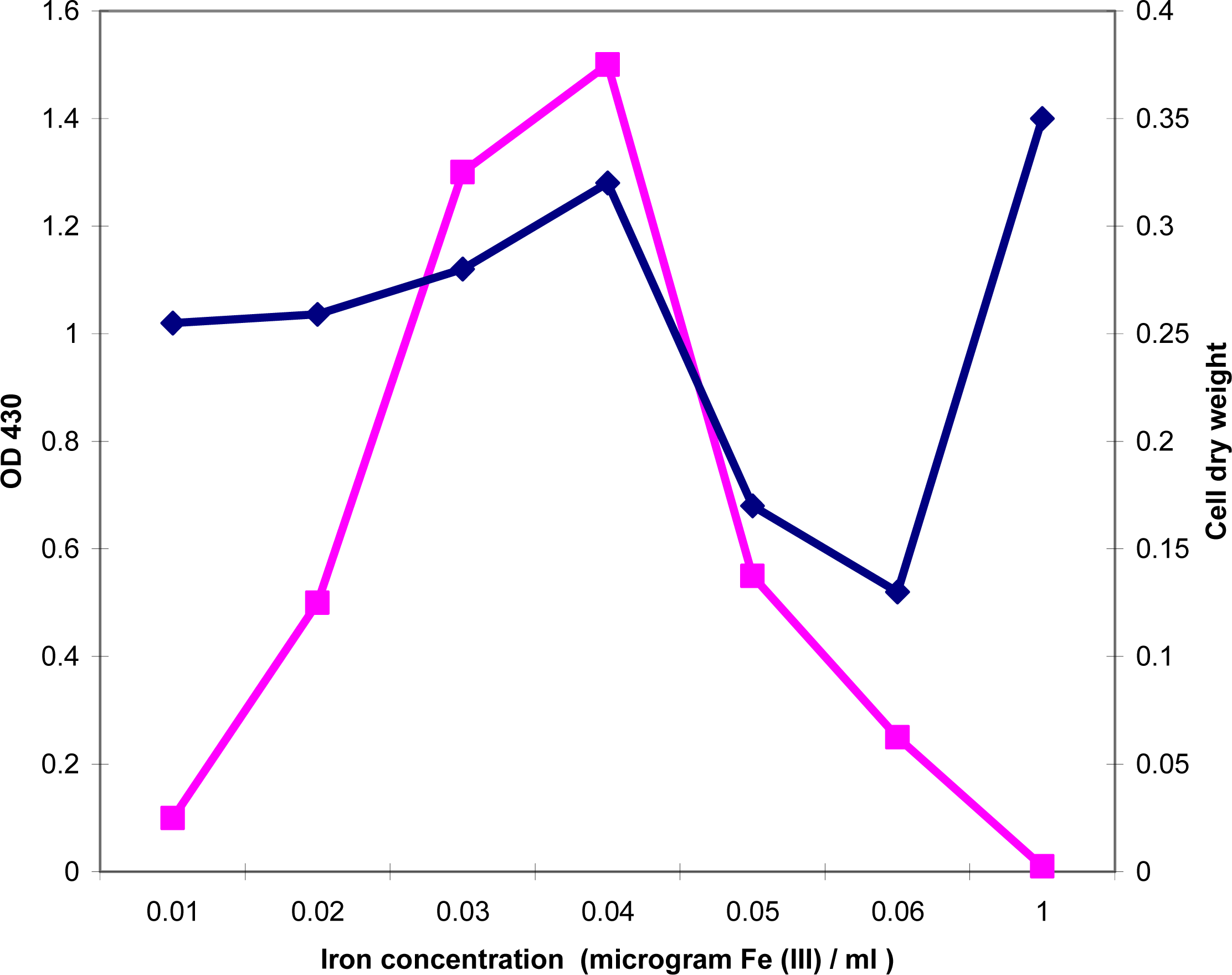
2.2. Production of exochelin in media containing various concentrations of Fe(III)
3. Discussion
4. Experimental Section
4.1. Bacterial strains, growth media and culture conditions
4.2. Purification and detection of exochelin
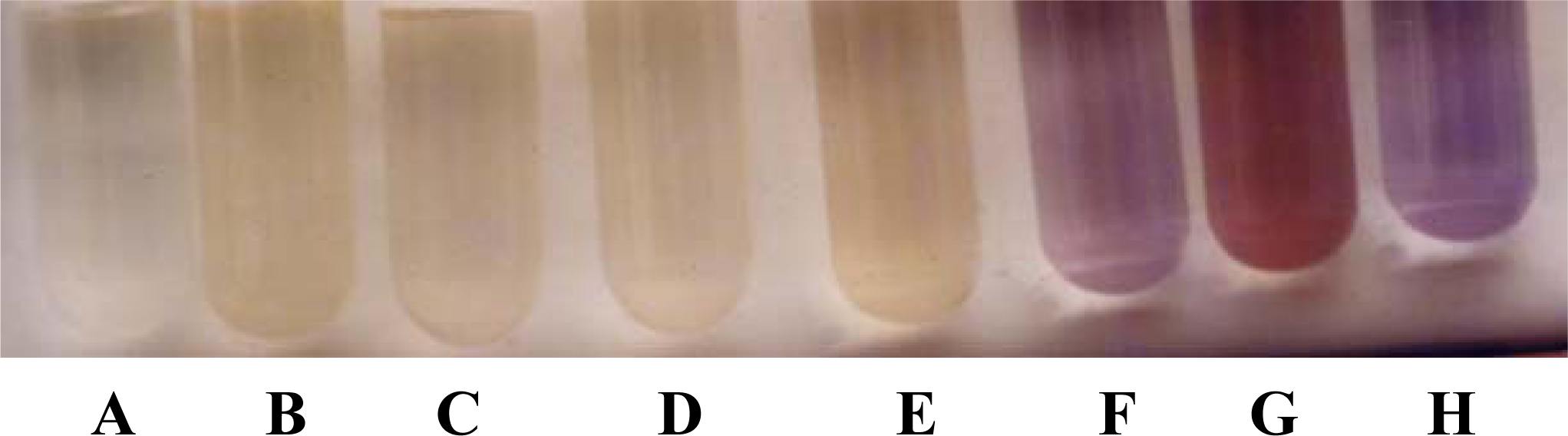
4.3. Chrome azural S assay
4.4. Estimation of cell dry weight
5. Conclusions
Acknowledgments
References
- Loper, JE; Henkels, MD. Availability of iron to Pseudomonas fluorescens in rhizosphere and bulk soil evaluated with an ice nucleation reporter gene. Appl. Environ. Microbiol 1997, 63, 99–105. [Google Scholar]
- Whipps, JM. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar]
- Miethke, M; Marahiel, MA. Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar]
- Messenger, AJM; Ratledge, C. Comprehensive Biotechnology; Pengamon Press: Oxford, 1984; pp. 275–295. [Google Scholar]
- Zhu, M; Arceneaux, JEL; Beggs, ML; Byers, BR; Eisenach, KD; Lundrigan, MD. Exochelin genes in Mycobacterium smegmatis: Identification of an ABC transporter and two nonribosomal peptide synthetase genes. Mol. Microbiol 1998, 29, 629–639. [Google Scholar]
- de Voss, JJ; Rutter, K; Schroeder, BG; Barry, CE, III. Iron acquisition and metabolism by mycobacteria. J. Bacteriol 1999, 181, 4443–4451. [Google Scholar]
- Gobin, J; Moore, CH; Reeve, JR, Jr; Wong, DK; Gibson, BW; Horwitz, MA. Iron acquisition by Mycobacterium tuberculosis: isolation and characterization of a family of ironbinding exochelins. Proc. Natl. Acad. Sci. USA 1995, 92, 5189–5193. [Google Scholar]
- O’Sullivan, DJ; O’Gara, F. Traits of fluorescent Pseudomonas spp. involved in suppression of plant root pathogens. Microbiol. Rev 1992, 56, 662–676. [Google Scholar]
- Ratledge, C; Hall, MJ. Influence of metal ions on the formation of mycobactin and salicylic acid in Mycobacterium smegmatis grown in static culture. J. Bacteriol 1971, 108, 314–319. [Google Scholar]
- Macham, LP; Stephenson, MC; Ratledge, C. Iron transport in Mycobacterium smegmatis: The isolation, purification and function of exochelin. J. Gen. Microbiol 1977, 101, 41–49. [Google Scholar]
- Sharman, GJ; Williams, DH; Ewing, DF; Ratledge, C. Determination of the structure of exochelin MN, the extracellular siderophore from Mycobacterium neoaurum. Chem. Biol 1995, 2, 553–561. [Google Scholar]
- McCready, KA; Ratledge, C. Amounts of iron, heme and related compounds in Mycobacterium smegmatis grown in various concentrations of iron. Biochem. Soc. Trans 1978, 6, 421–423. [Google Scholar]
- Loper, JE; Henkels, MD. Availability of iron to Pseudomonas fluorescens in rhizosphere and bulk soil evaluated with an ice nucleation reporter gene. Appl. Environ. Microbiol 1997, 63, 99–105. [Google Scholar]
- Macham, LP; Ratledge, C; Nocton, JC. Extracellular iron acquisition by mycobacteria: Role of exochelins and evidence against the participation of mycobactin. Infect. Immun 1975, 12, 1242–1251. [Google Scholar]
- Barclay, R; Ratledge, C. Iron-binding compounds of Mycobacterium avium, M. intracellulare, M. scrofulaceum, and mycobactin-dependent M. paratuberculosis and M. avium. J. Bacteriol 1983, 153, 1138–1146. [Google Scholar]
- Lewin, R. How microorganisms transport iron. Science 1984, 225, 401–402. [Google Scholar]
- Litwin, CM; Calderwood, SB. Role of iron in regulation of virulence genes. Clin. Microbiol. Rev. 1993, 6, 137–147. [Google Scholar]
- Loper, JE; Henkels, MD. Utilization of heterologous siderophores enhances levels of iron available to Pseudomonas putida in the rhizosphere. Appl. Environ. Microbiol 1999, 65, 5357–5363. [Google Scholar]
- Leeman, M; den Ouden, FM; van Pelt, JA; Dirkx, FPM; Steijl, H; Bakker, PAHM; Schippers, B. Iron availability affects induction of systemic resistance to Fusarium wilt of radish by Pseudomonas fluorescens. Phytopathology 1996, 86, 149–155. [Google Scholar]
- Winder, FG; O’Hara, C. Effects of iron deficiency and of zinc deficiency on the activities of some enzymes in Mycobacterium smegmatis. Biochem. J 1964, 90, 122–126. [Google Scholar]
- Schwyn, B; Neilands, JB. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar]




© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chan, K.-G. Exochelin Production in Mycobacterium neoaurum. Int. J. Mol. Sci. 2009, 10, 345-353. https://doi.org/10.3390/ijms10010345
Chan K-G. Exochelin Production in Mycobacterium neoaurum. International Journal of Molecular Sciences. 2009; 10(1):345-353. https://doi.org/10.3390/ijms10010345
Chicago/Turabian StyleChan, Kok-Gan. 2009. "Exochelin Production in Mycobacterium neoaurum" International Journal of Molecular Sciences 10, no. 1: 345-353. https://doi.org/10.3390/ijms10010345
APA StyleChan, K.-G. (2009). Exochelin Production in Mycobacterium neoaurum. International Journal of Molecular Sciences, 10(1), 345-353. https://doi.org/10.3390/ijms10010345