Antioxidant Effects of the Ethanol Extract from Flower of Camellia japonica via Scavenging of Reactive Oxygen Species and Induction of Antioxidant Enzymes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
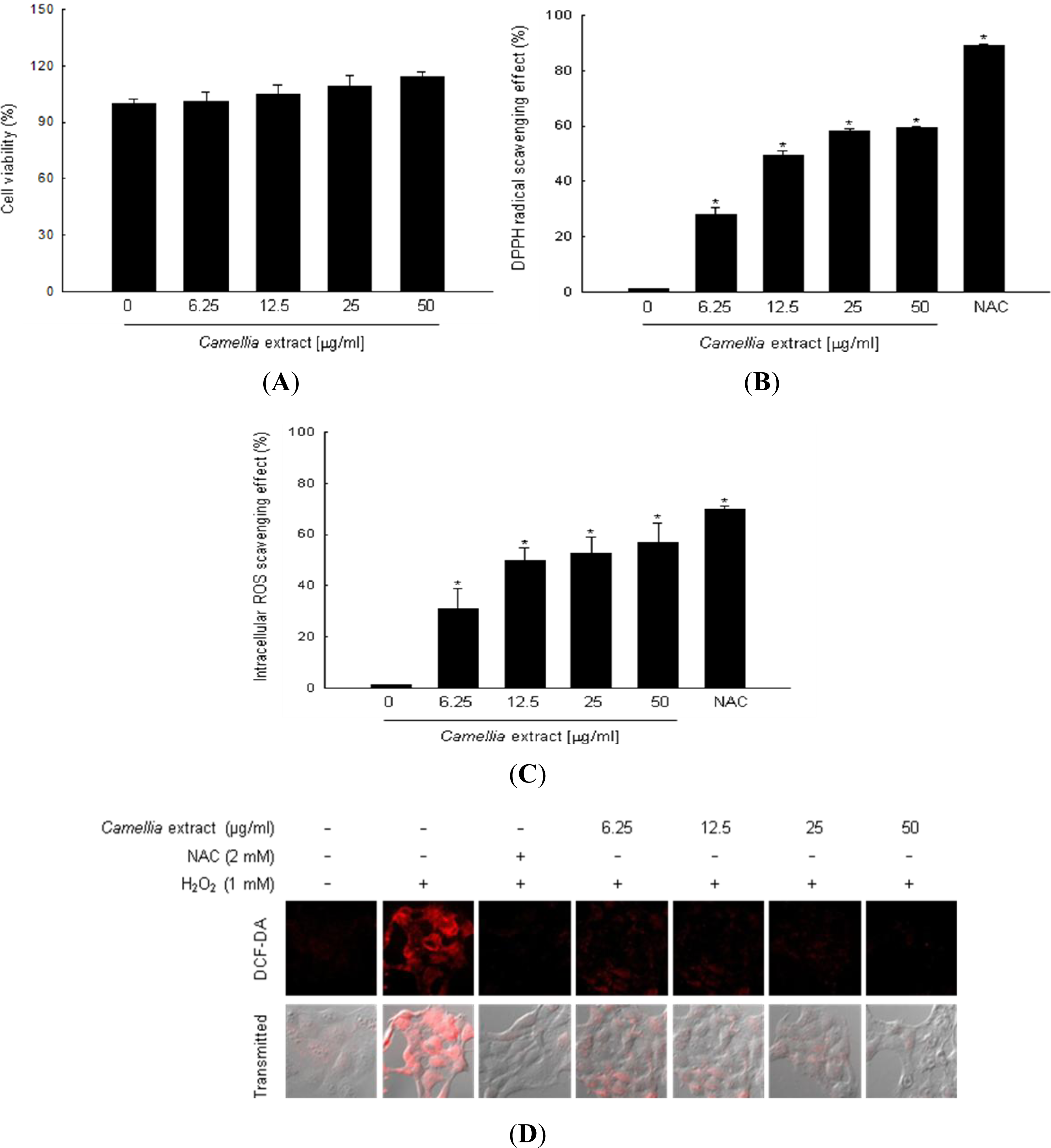
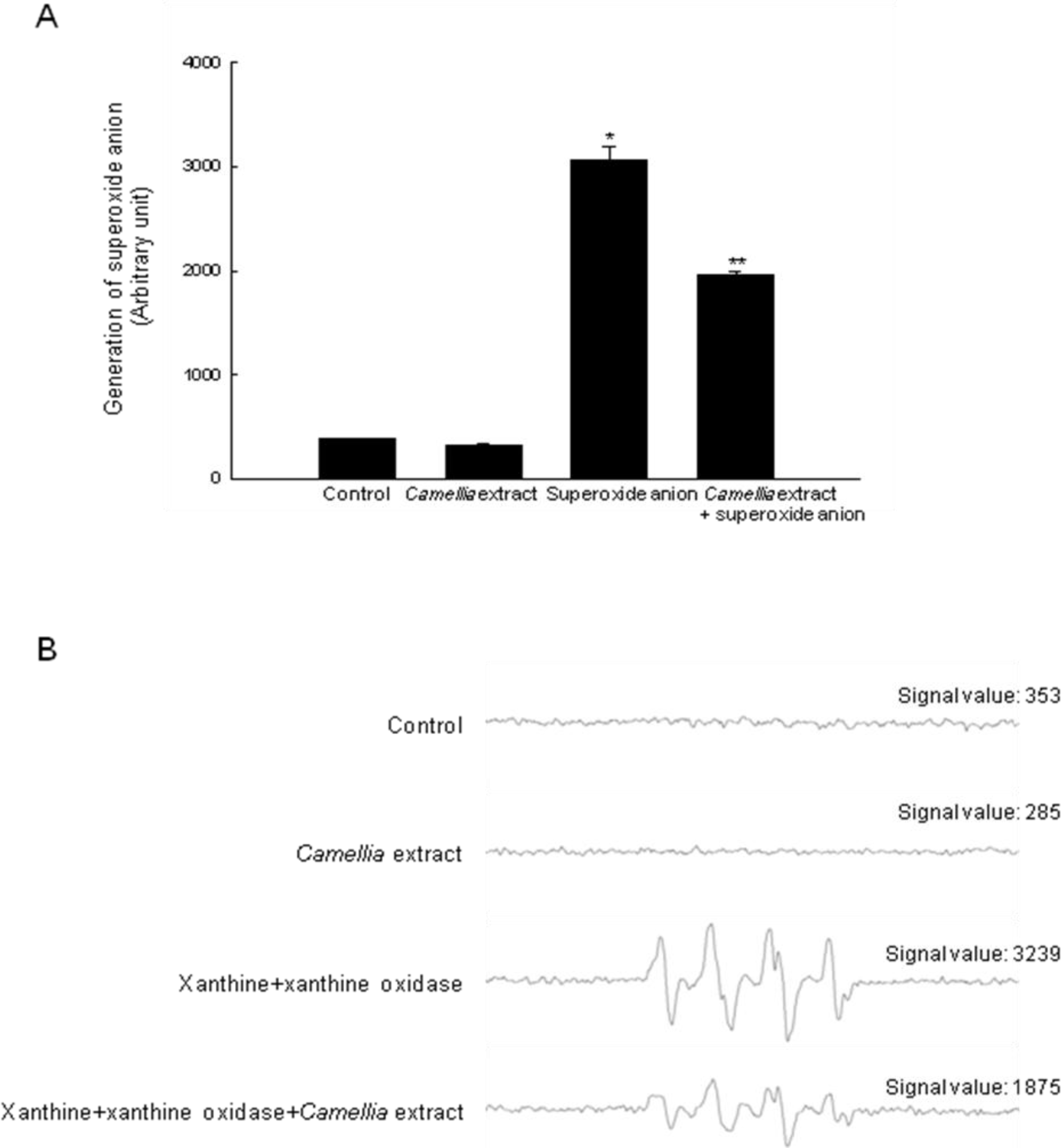
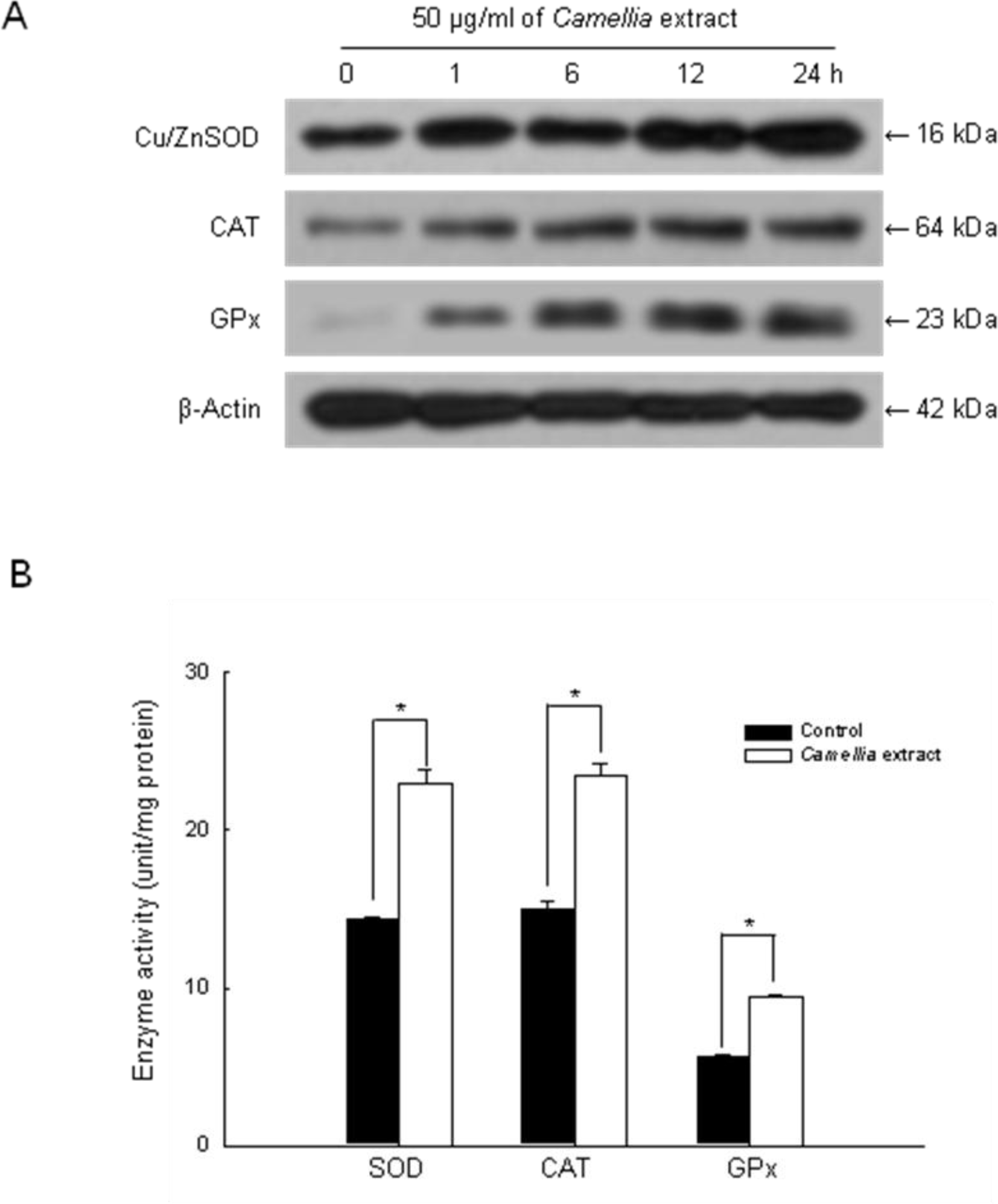
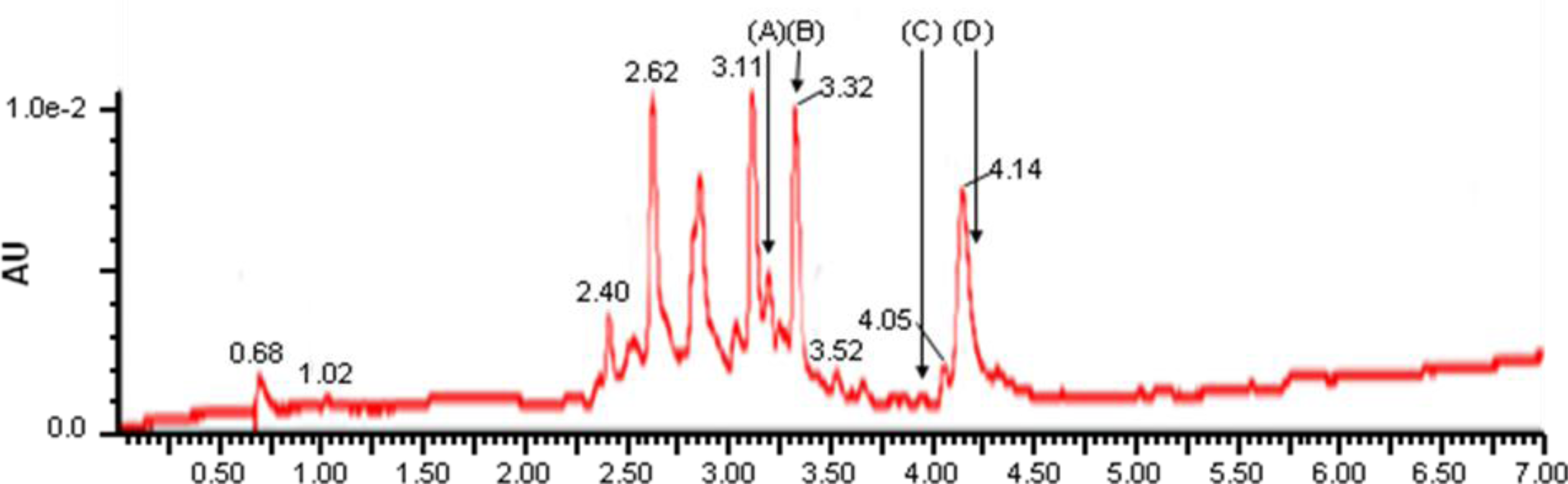
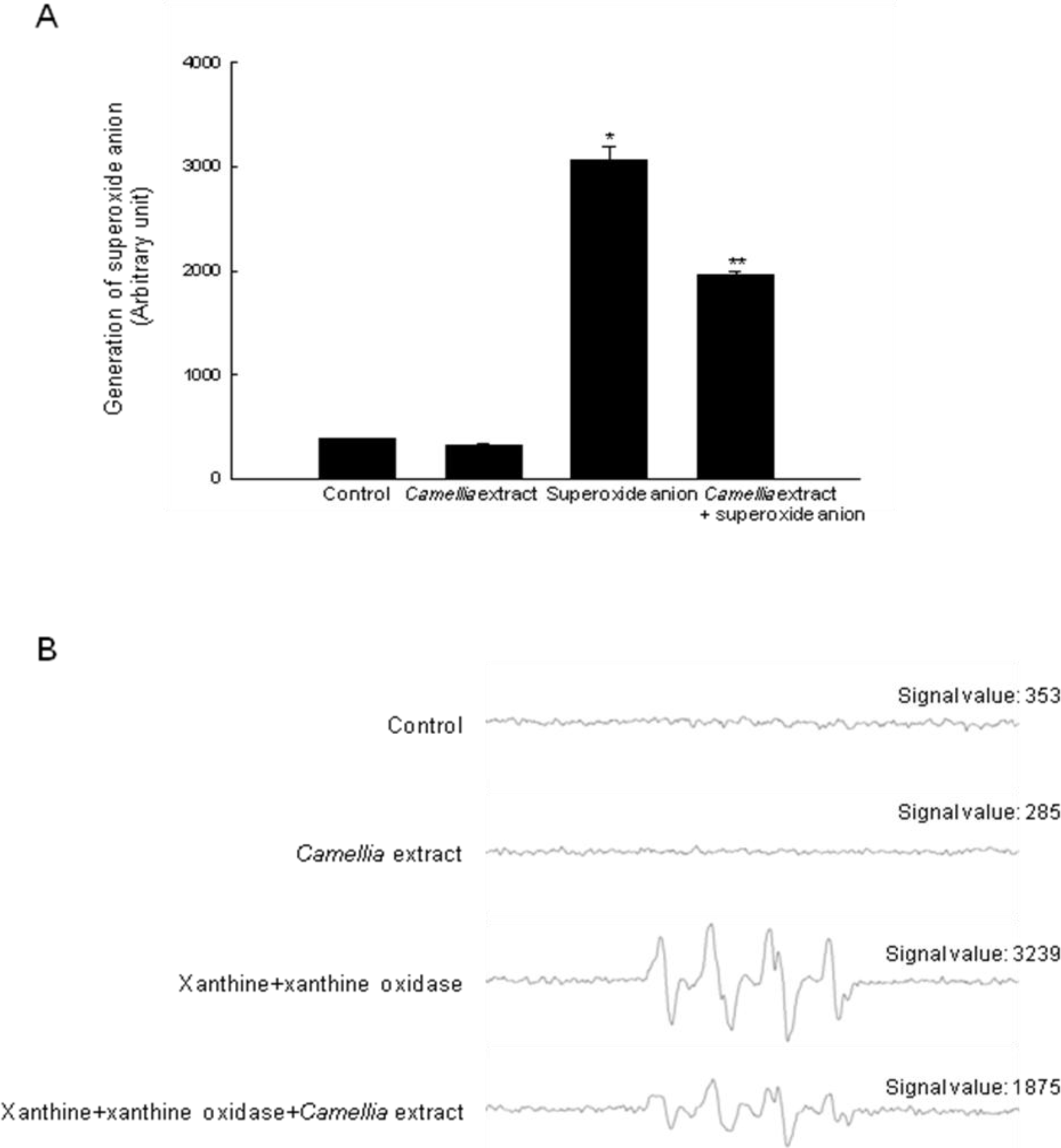
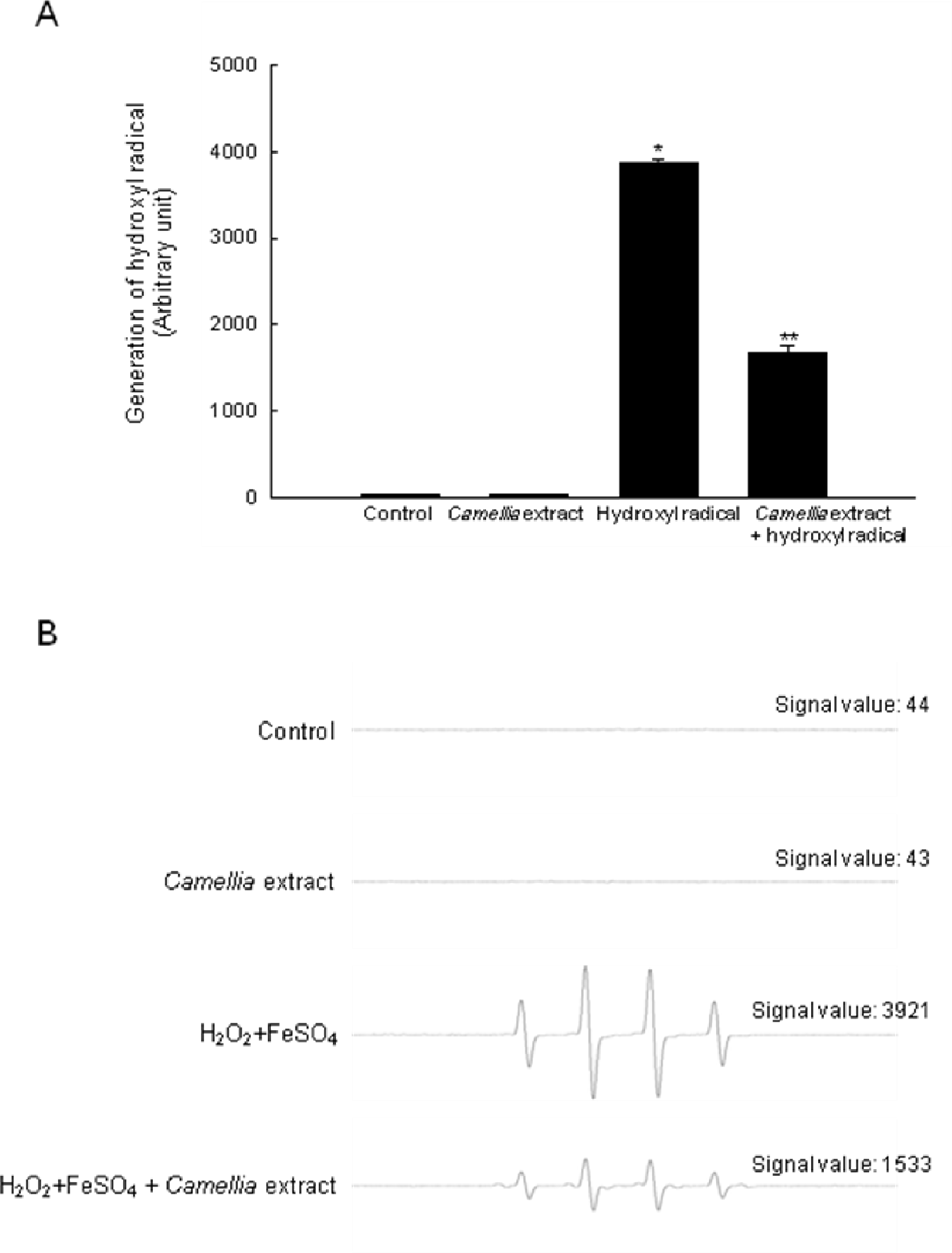
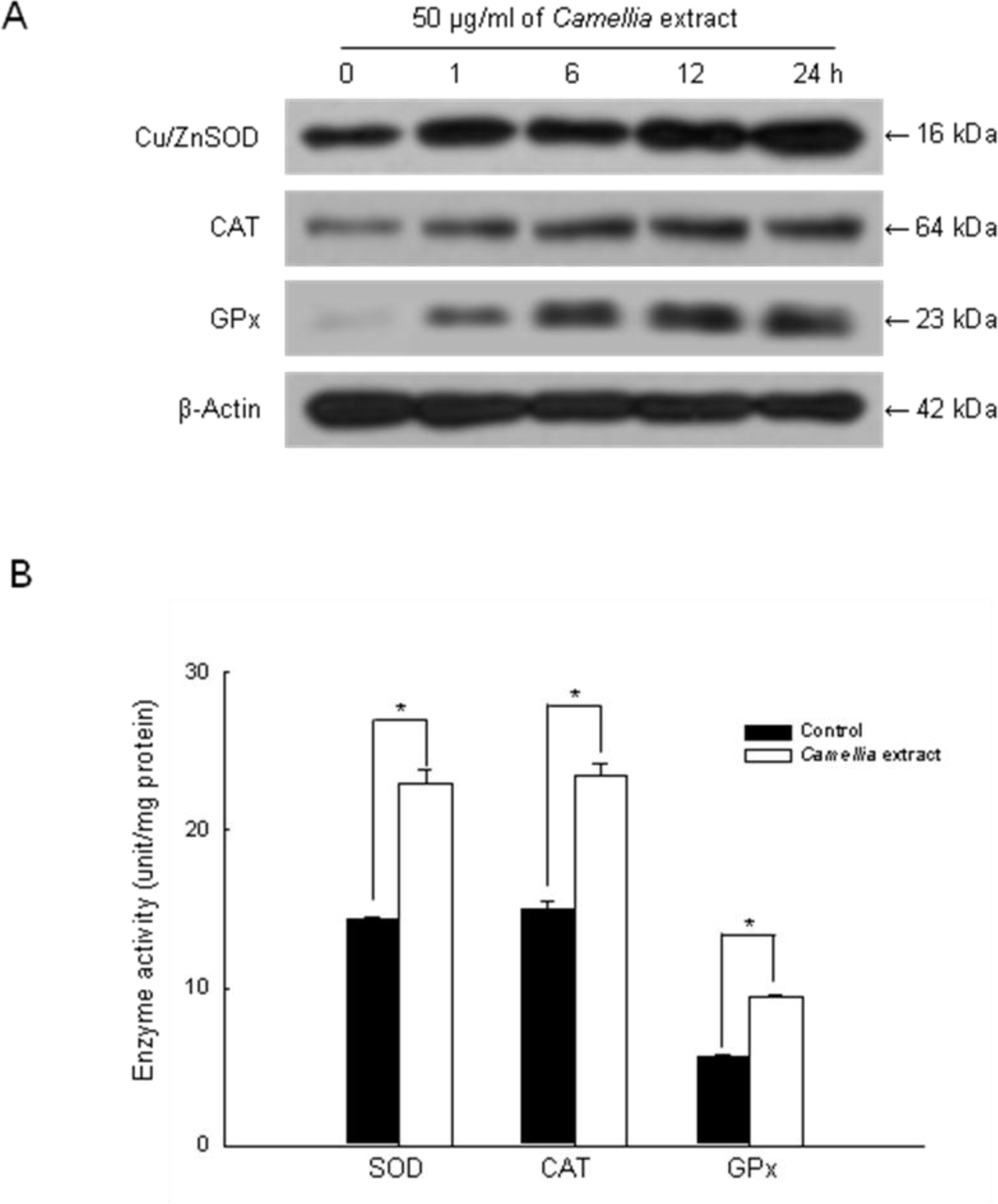
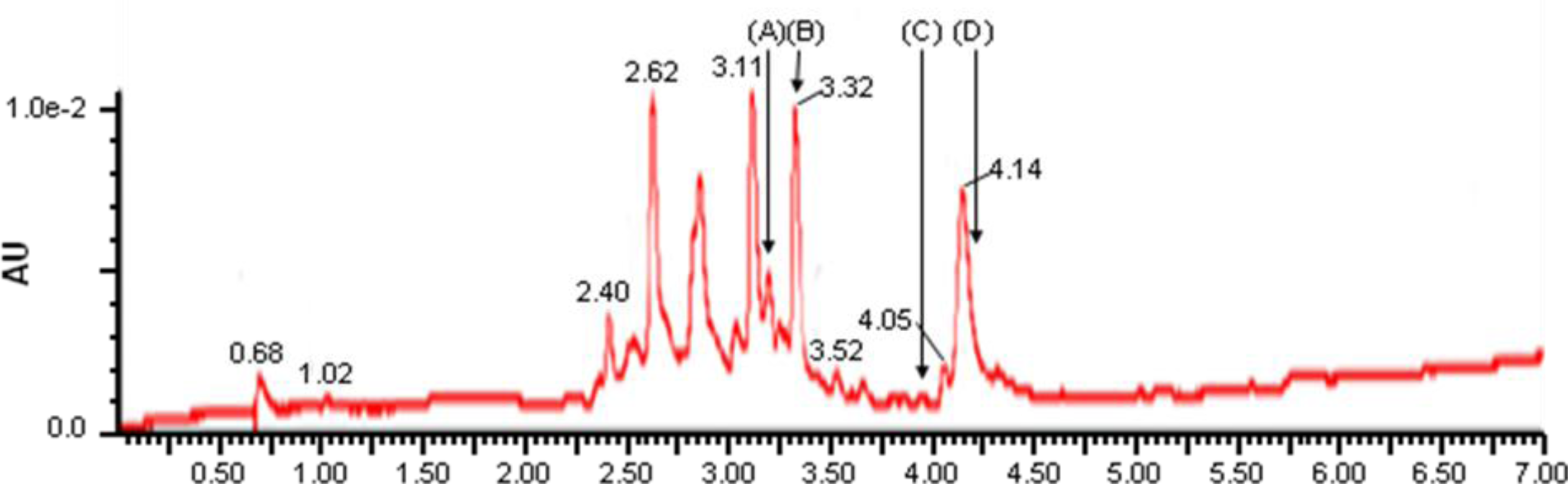
2. Results
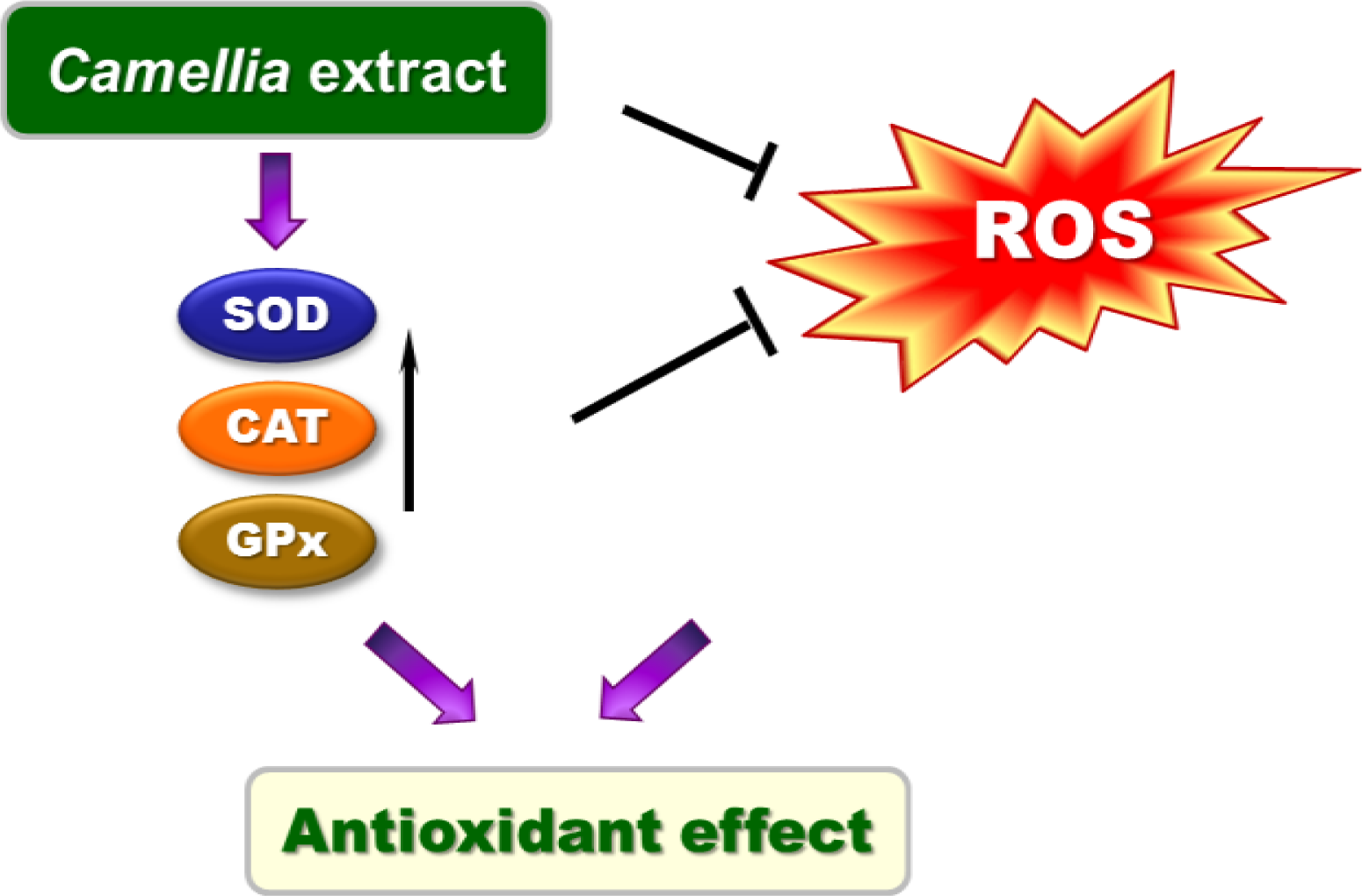
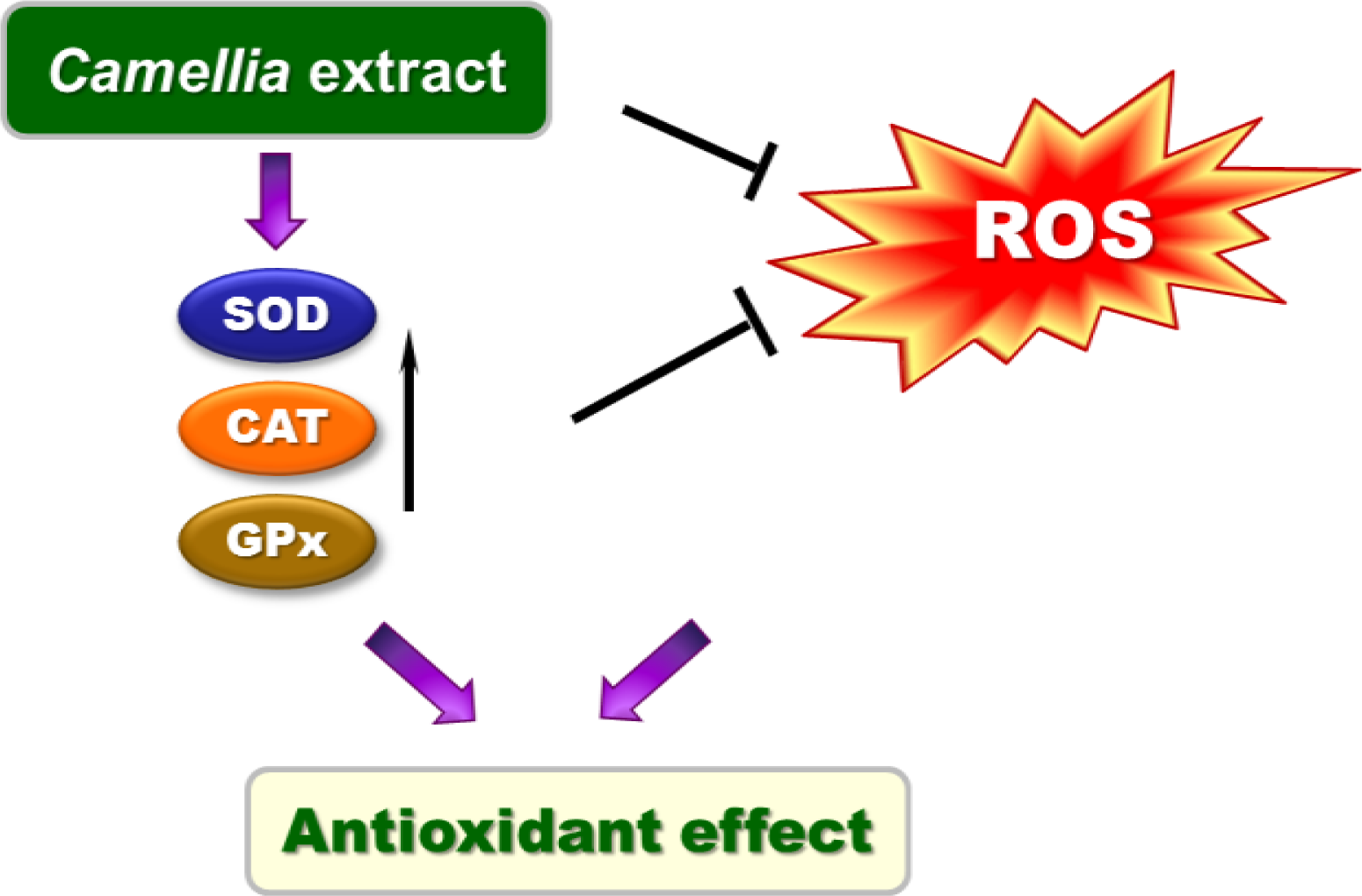
3. Discussion
4. Experimental Section
4.1. Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. DPPH Radical Scavenging Activity
4.5. Intracellular ROS Scavenging Activity
4.6. Detection of Superoxide Anion
4.7. Detection of Hydroxyl Radical
4.8. Western Blot Analysis
4.9. Measurement of SOD Activity
4.10. Measurement of CAT Activity
4.11. Measurement of GPx Activity
4.12. Statistical Analysis
Acknowledgments
References
- Ross, C; Alston, M; Bickenbach, JR; Aykin-Burns, N. Oxygen tension changes the rate of migration of human skin keratinocytes in an age-related manner. Exp. Dermatol 2011, 20, 58–63. [Google Scholar]
- Laskin, JD; Black, AT; Jan, YH; Sinko, PJ; Heindel, ND; Sunil, V; Heck, DE; Laskin, DL. Oxidants and antioxidants in sulfur mustard-induced injury. Ann. NY Acad. Sci 2010, 1203, 92–100. [Google Scholar]
- Vileno, B; Jeney, S; Sienkiewicz, A; Marcoux, PR; Miller, LM; Forró, L. Evidence of lipid peroxidation and protein phosphorylation in cells upon oxidative stress photo-generated by fullerols. Biophys. Chem 2010, 152, 164–169. [Google Scholar]
- Dreger, H; Westphal, K; Weller, A; Baumann, G; Stangl, V; Meiners, S; Stangl, K. Nrf2-dependent upregulation of antioxidative enzymes: A novel pathway for proteasome inhibitor-mediated cardioprotection. Cardiovasc. Res 2009, 83, 354–361. [Google Scholar]
- Vertuani, S; Angusti, A; Manfredini, S. The antioxidants and pro-antioxidants network: An overview. Curr. Pharm. Des 2004, 10, 1677–1694. [Google Scholar]
- Habdous, M; Herbeth, B; Vincent-Viry, M; Lamont, JV; Fitzgerald, PS; Visvikis, S; Siest, G. Serum total antioxidant status, erythrocyte superoxide dismutase and whole-blood glutathione peroxidase activities in the Stanislas cohort: Influencing factors and reference intervals. Clin. Chem. Lab. Med 2003, 41, 209–215. [Google Scholar]
- Yoshikawa, M; Morikawa, T; Asao, Y; Fujiwara, E; Nakamura, S; Matsuda, H. Medicinal flowers. XV. The structures of noroleanane- and oleanane-type triterpene oligoglycosides with gastroprotective and platelet aggregation activities from flower buds of Camellia japonica L. Chem. Pharm. Bull 2007, 55, 606–612. [Google Scholar]
- Itokawa, H; Sawada, N; Murakami, T. Structures of camelliagenins A, B, and C obtained from Camellia japonica. Chem. Pharm. Bull 1969, 17, 474–480. [Google Scholar]
- Yoshikawa, M; Harada, E; Murakami, T; Matsuda, H; Yamahara, J; Murakami, N. Camelliasaponins B1, B2, C1 and C2, new type inhibitors of ethanol absorption in rats from the seeds of Camellia japonica L. Chem. Pharm. Bull 1994, 42, 742–744. [Google Scholar]
- Onodera, K; Hanashiro, K; Yasumoto, T. Camellianoside, a novel antioxidant glycoside from the leaves of Camellia japonica. Biosci. Biotech. Biochem 2006, 70, 1995–1998. [Google Scholar]
- Itokawa, H; Nakajima, H; Ikuta, A; Iitaka, Y. Two triterpenes from the flowers of Camellia japonica. Phytochemistry 1981, 20, 2539–2542. [Google Scholar]
- Nakajima, H; Itokawa, H; Ikuta, A. Studies on the constituents of the flower of Camellia japonica (2). Yakugaku Zasshi 1984, 104, 157–161. (in Japanese).. [Google Scholar]
- Afanasev, IB; Dorozhko, AI; Brodskii, AV; Kostyuk, VA; Potapovitch, AI. Chelating and free radical scavenging mechanisms of inhibitory action of rutin and quercetin in lipid peroxidation. Biochem. Pharmacol 1989, 38, 1763–1769. [Google Scholar]
- Van Acker, SA; van Balen, GP; van den Berg, DJ; Bast, A; van der Vijgh, WJ. Influence of iron chelation on the antioxidant activity of flavonoids. Biochem. Pharmacol 1998, 56, 935–943. [Google Scholar]
- Saija, A; Scalese, M; Lanza, M; Marzullo, D; Bonina, F; Castelli, F. Flavonoids as antioxidant agents: Importance of their interaction with biomembranes. Free Radic. Biol. Med 1995, 19, 481–486. [Google Scholar]
- Jung, HA; Jung, MJ; Kim, JY; Chung, HY; Choi, JS. Inhibitory activity of flavonoids from Prunus davidiana and other flavonoids on total ROS and hydroxyl radical generation. Arch. Pharm. Res 2003, 26, 809–815. [Google Scholar]
- Samhan-Arias, AK; Martín-Romero, FJ; Gutiérrez-Merino, C. Kaempferol blocks oxidative stress in cerebellar granule cells and reveals a key role for reactive oxygen species production at the plasma membrane in the commitment to apoptosis. Free Radic. Biol. Med 2004, 37, 48–61. [Google Scholar]
- Emerit, I; Filipe, P; Freitas, J; Vassy, J. Protective effect of superoxide dismutase against hair graying in a mouse model. Photochem. Photobiol 2004, 80, 579–582. [Google Scholar]
- Alscher, RG; Erturk, N; Heath, LS. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot 2002, 53, 1331–1341. [Google Scholar]
- Bose Girigoswami, K; Bhaumik, G; Ghosh, R. Induced resistance in cells exposed to repeated low doses of H2O2 involves enhanced activity of antioxidant enzymes. Cell Biol. Int 2005, 29, 761–767. [Google Scholar]
- Rajeshkumar, NV; Ramadasan, K. Modulation of carcinogenic response and antioxidant enzymes of rats administered with 1, 2-dimethyl hydrazine by Picroliv. Cancer Lett 2003, 191, 137–143. [Google Scholar]
- Thu, VT; Kim, HK; Ha, SH; Yoo, JY; Park, WS; Kim, N; Oh, GT; Han, J. Glutathione peroxidase 1 protects mitochondria against hypoxia/reoxygenation damage in mouse hearts. Pflugers. Arch 2010, 460, 55–68. [Google Scholar]
- Erden, IM; Kahraman, A. The protective effect of flavonol quercetin against ultraviolet a induced oxidative stress in rats. Toxicology 2000, 154, 21–29. [Google Scholar]
- Inal, M; Altinişik, M; Bilgin, MD. The effect of quercetin on renal ischemia and reperfusion injury in the rat. Cell. Biochem. Funct 2002, 20, 291–296. [Google Scholar]
- Kameoka, S; Leavitt, P; Chang, C; Kuo, SM. Expression of antioxidant proteins in human intestinal Caco-2 cells treated with dietary flavonoids. Cancer Lett 1999, 146, 161–167. [Google Scholar]
- Jung, CH; Jun, CY; Lee, S; Park, CH; Cho, K; Ko, SG. Rhus verniciflua stokes extract: Radical scavenging activities and protective effects on H2O2-induced cytotoxicity in macrophage RAW 264.7 cell lines. Biol. Pharm. Bull 2006, 29, 1603–1607. [Google Scholar]
- Söhretoğlu, D; Sabuncuoğlu, SA; Sakar, MK; Ozgüneş, H; Sterner, O. Antioxidant effects of secondary metabolites from Geranium psilostemon. Nat. Prod. Commun 2010, 5, 899–902. [Google Scholar]
- Kandaswami, C; Middleton, E, Jr. Free radical scavenging and antioxidant activity of plant flavonoids. Adv. Exp. Med. Biol 1994, 366, 351–376. [Google Scholar]
- Rosenkranz, AR; Schmaldienst, S; Stuhlmeier, KM; Chen, W; Knapp, W; Zlabinger, GJ. A microplate assay for the detection of oxidative products using 2’,7’-dichlorofluorescin-diacetate. J. Immunol. Method 1992, 156, 39–45. [Google Scholar]
- Rimbach, G; Weinberg, PD; de Pascual-Teresa, S; Alonso, MG; Ewins, BA; Turner, R; Minihane, AM; Botting, N; Fairley, B; Matsugo, S; Uchida, Y; Cassidy, A. Sulfation of genistein alters its antioxidant properties and its effect on platelet aggregation and monocyte and endothelial function. Biochim. Biophys. Acta 2004, 1670, 229–237. [Google Scholar]

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Piao, M.J.; Yoo, E.S.; Koh, Y.S.; Kang, H.K.; Kim, J.; Kim, Y.J.; Kang, H.H.; Hyun, J.W. Antioxidant Effects of the Ethanol Extract from Flower of Camellia japonica via Scavenging of Reactive Oxygen Species and Induction of Antioxidant Enzymes. Int. J. Mol. Sci. 2011, 12, 2618-2630. https://doi.org/10.3390/ijms12042618
Piao MJ, Yoo ES, Koh YS, Kang HK, Kim J, Kim YJ, Kang HH, Hyun JW. Antioxidant Effects of the Ethanol Extract from Flower of Camellia japonica via Scavenging of Reactive Oxygen Species and Induction of Antioxidant Enzymes. International Journal of Molecular Sciences. 2011; 12(4):2618-2630. https://doi.org/10.3390/ijms12042618
Chicago/Turabian StylePiao, Mei Jing, Eun Sook Yoo, Young Sang Koh, Hee Kyoung Kang, Junoh Kim, Yong Jin Kim, Hak Hee Kang, and Jin Won Hyun. 2011. "Antioxidant Effects of the Ethanol Extract from Flower of Camellia japonica via Scavenging of Reactive Oxygen Species and Induction of Antioxidant Enzymes" International Journal of Molecular Sciences 12, no. 4: 2618-2630. https://doi.org/10.3390/ijms12042618
APA StylePiao, M. J., Yoo, E. S., Koh, Y. S., Kang, H. K., Kim, J., Kim, Y. J., Kang, H. H., & Hyun, J. W. (2011). Antioxidant Effects of the Ethanol Extract from Flower of Camellia japonica via Scavenging of Reactive Oxygen Species and Induction of Antioxidant Enzymes. International Journal of Molecular Sciences, 12(4), 2618-2630. https://doi.org/10.3390/ijms12042618