Genetic Diversity and Phylogeny of Antagonistic Bacteria against Phytophthora nicotianae Isolated from Tobacco Rhizosphere

Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation of Bacteria
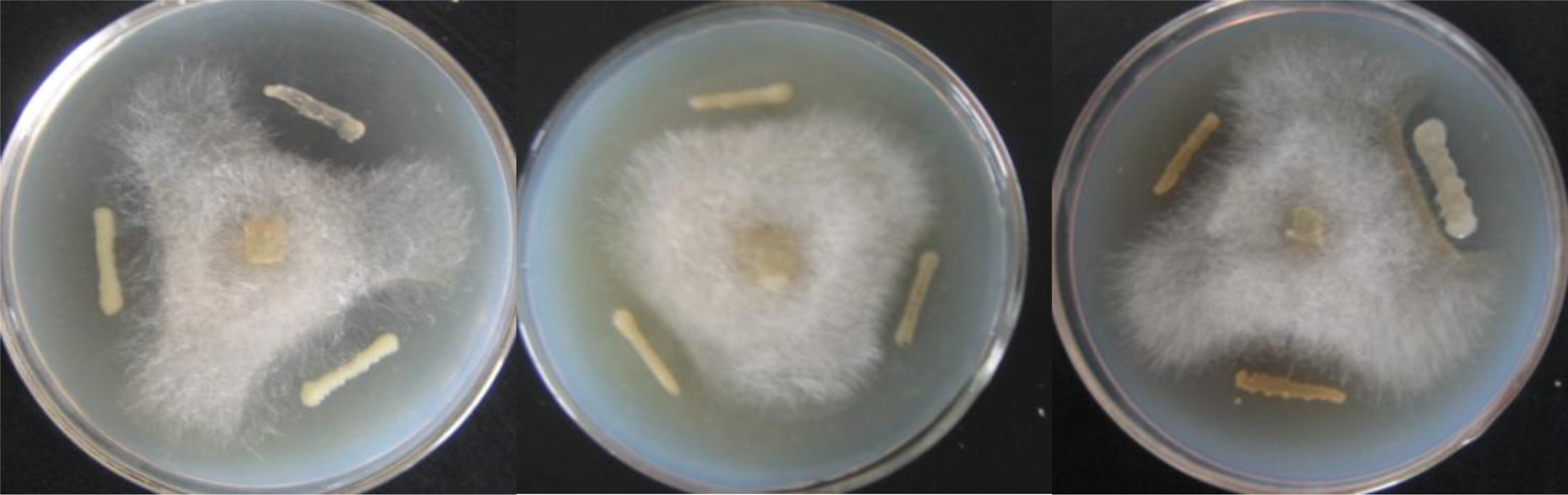
2.2. Screening of Antagonistic Bacteria
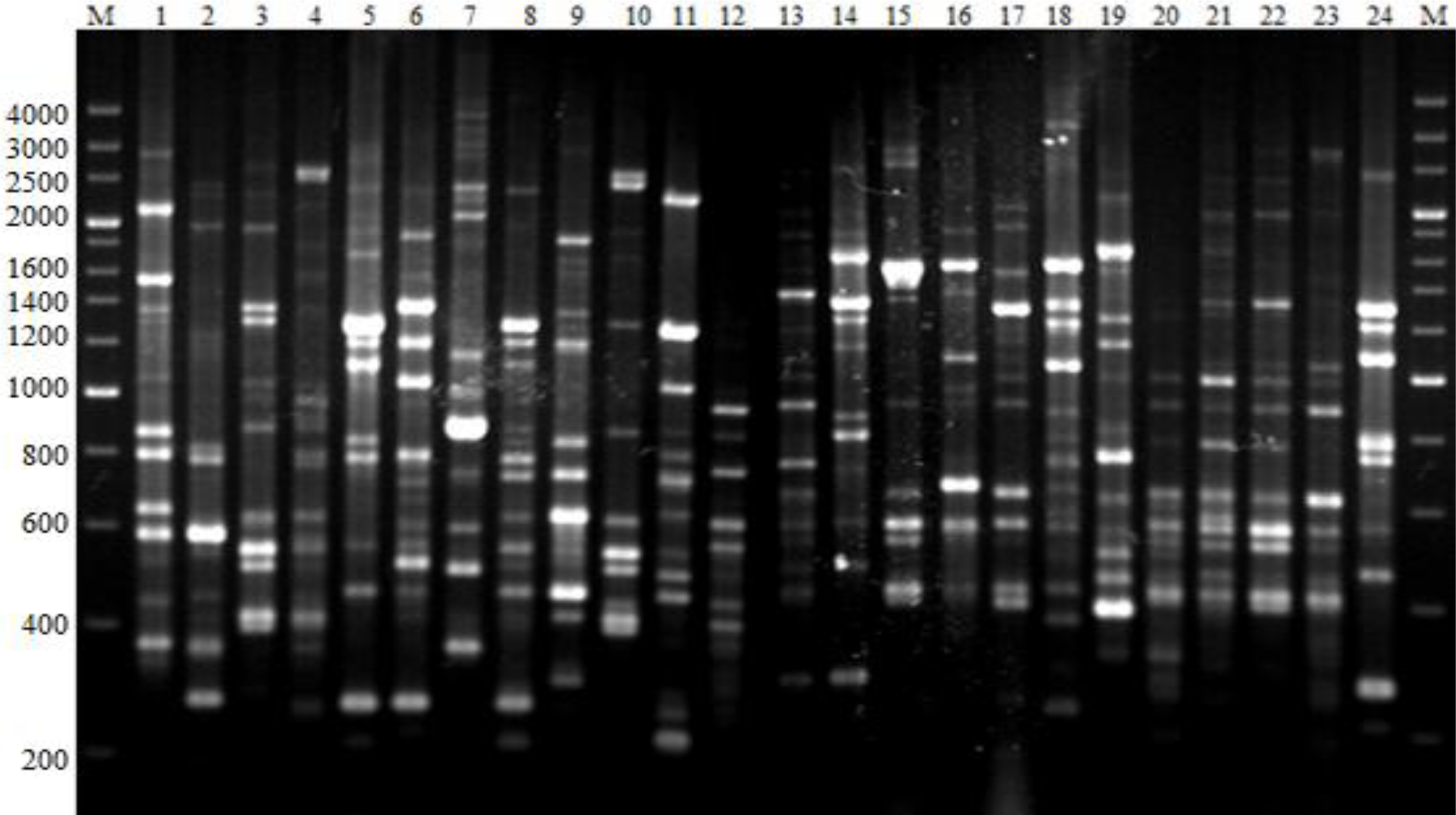
2.3. BOXAIR-PCR Genomic Fingerprints
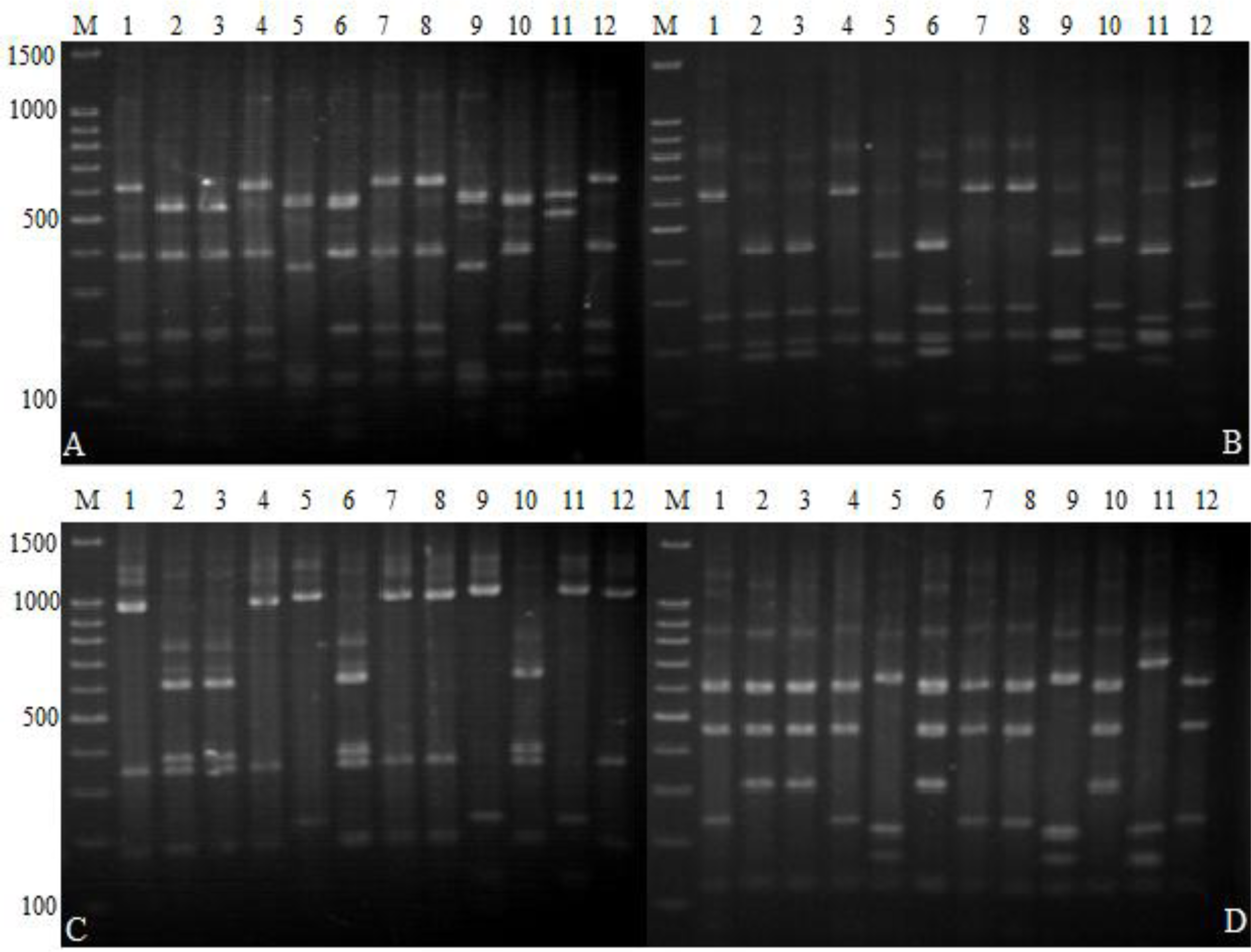
2.4. 16S-RFLP Analysis
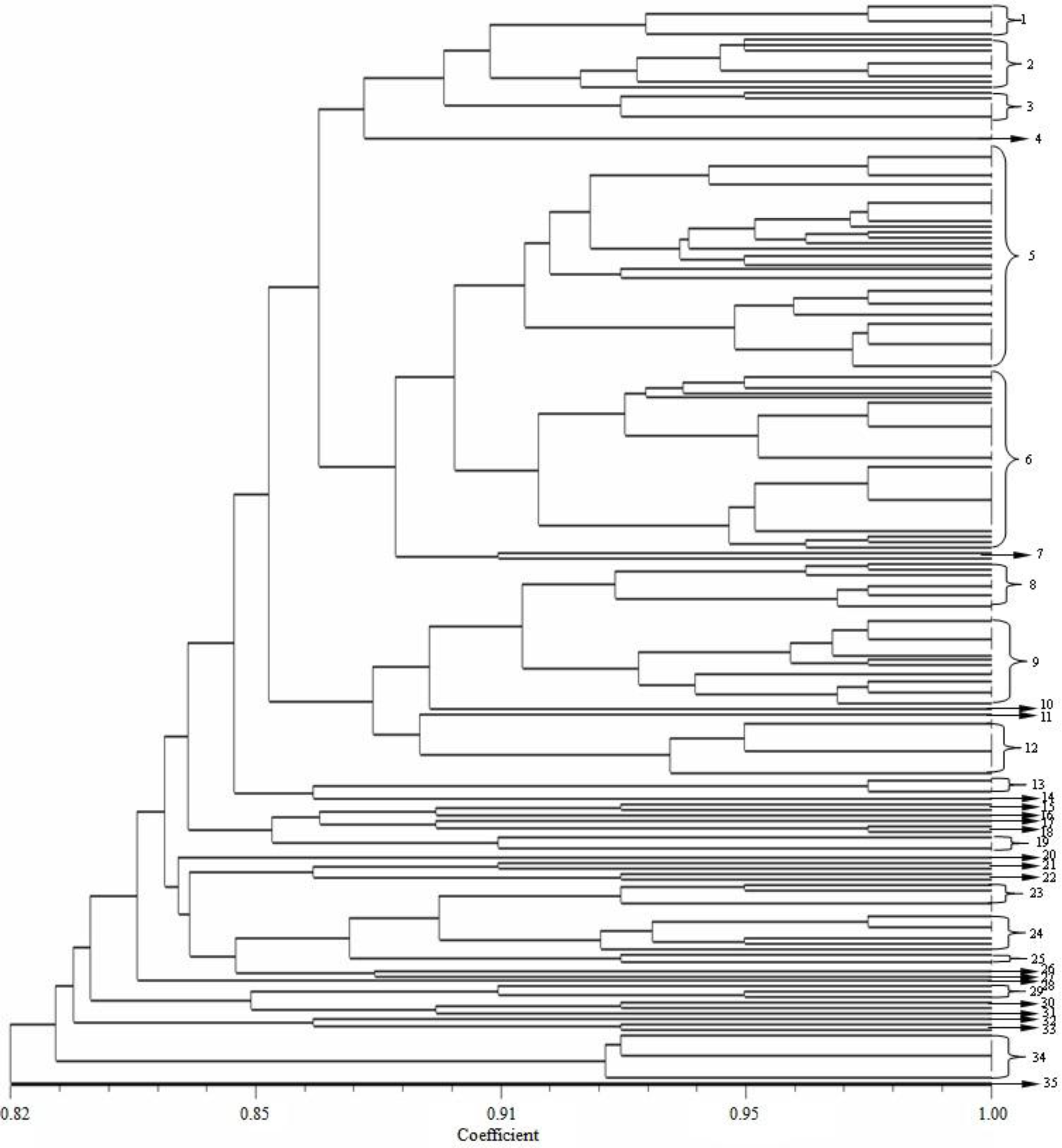
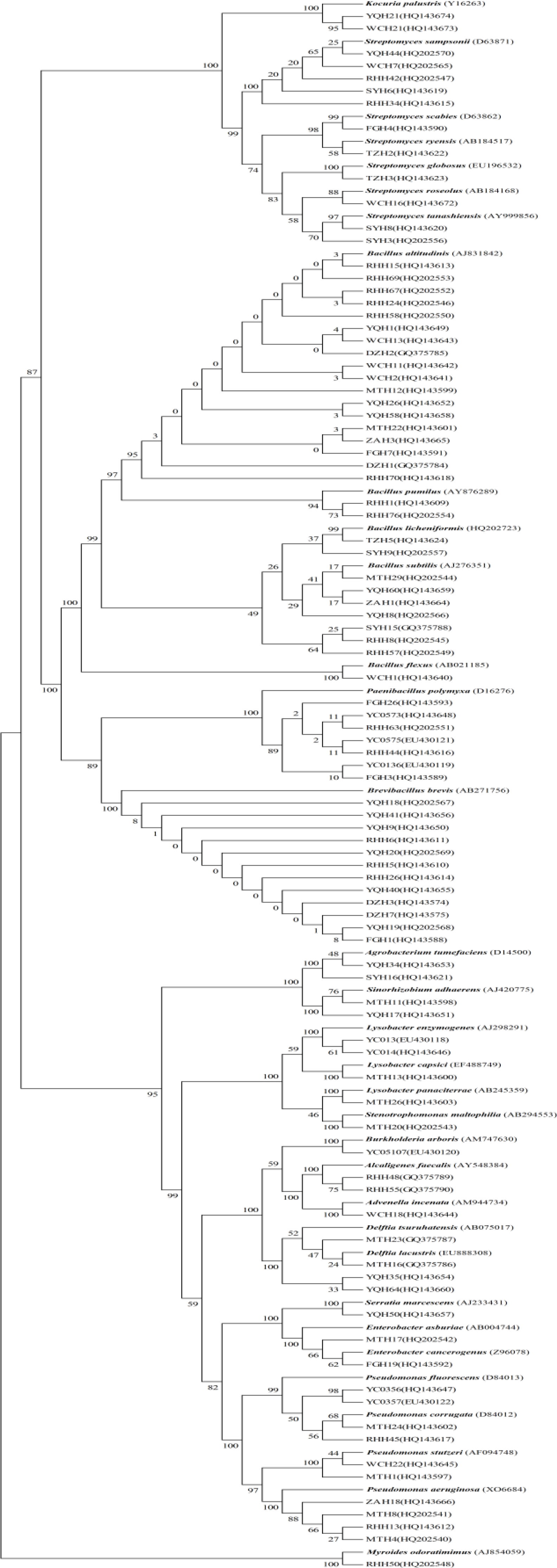
2.5. Sequencing and Phylogeny Analysis
2.6. Species Diversity of Antagonistic Bacteria
3. Experimental Section
3.1. Rhizosphere Soil
3.2. Isolation of Bacteria
3.3. Screening of Antagonistic Bacteria
3.4. Total DNA Extraction and 16S rDNA PCR Amplification
3.5. BOXAIR-PCR Genomic Fingerprints
3.6. 16S-RFLP Analysis
3.7. Sequencing and Phylogeny Analysis
4. Conclusions
Acknowledgments
References
- Tian, F; Ding, Y; Du, B. Genetic Diversity of Siderophore-producing Bacteria of Tobacco Rhizosphere. Braz. J. Microbiol 2009, 40, 276–284. [Google Scholar]
- Erwin, DC; Ribeiro, OK. Phytophthora disease worldwide; American Phytopathology Society Press: St Paul, MN, USA, 1996; pp. 98–138. [Google Scholar]
- Cartwright, DK; Spurr, HW, Jr. Biological control of Phytophthora parasitica var. Nicotianae on tobacco seedlings with non-pathogenic binucleate Rhizoctonia fungi. Soil Biol. Biochem 1998, 30, 1879–1884. [Google Scholar]
- Zhang, XG; Sun, WX; Chang, CJ. Genetic and pathogenic Variation Among Tobacco Black Shank Strains of Phytophthora parasitica var. Nicotianae from the Main Tobacco Growing in China. J. Phytopathol 2003, 151, 259–266. [Google Scholar]
- Melton, TA; Broadwell, A; Wilson, J. Disease management; Flue-Cured Tobacco Information; North Carolina State University: Raleigh, NC, USA, 1998; pp. 92–171. [Google Scholar]
- Wei, G; Kloepper, JW; Tuzun, S. Induction of systemic resistance of cucumber to Colletotrichum orbiculare by select strains of plant growth-promoting rhizobacteria. Phytopathology 1991, 81, 1508–1512. [Google Scholar]
- Van Peer, R; Niemann, R; Schippers, B. Induced resistance and phytoalexin accumulation in biological control of Fusarium wilt of carnation by Pseudomonas sp. strain WCS 417r. Phytopathology 1991, 81, 728–734. [Google Scholar]
- Maurhofer, M. Induction of systemic resistance of tobacco to Tobacco Necrosis Virus by the root-colonizing Pseudomonas fluorescens strain CHAO: Influence of the gacA gene and of pyoverdine production. Phytopathology 1994, 84, 139–146. [Google Scholar]
- Defago, G; Berling, CH; Wurthrich, B. Suppression of black root rot of tobacco and other root diseases by strains of Pseudomonas fluorescens: Potential applications and mechanisms. In Biological Control of Soil-Borne Plant Pathogens; Hornby, D, Cook, RJ, Henis, Y, Ko, WH, Rovira, AD, Schippers, B, Scott, PR, Eds.; CAB International: Wallingford, UK, 1990; pp. 93–108. [Google Scholar]
- Weller, DM. Biological control of soilborne plant pathogens in the rhizosphere with bacteria. Annu. Rev. Phytopathol 1998, 26, 379–407. [Google Scholar]
- Bloemberg, GV; Lugtenberg, BJ. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol 2001, 4, 343–350. [Google Scholar]
- Kurze, S. Biological Control of Fungal Strawberry Diseases by Serratia plymuthica HRO-C48. Plant Dis 2001, 85, 529–534. [Google Scholar]
- Emmert, EAB; Handelsman, J. Biocontrol of plant disease: A (gram-) positive perspective. FEMS Microbiol. Lett 1999, 171, 1–9. [Google Scholar]
- Whipps, JM. Ecological considerations involved in commercial development of biological control agents for soil-borne diseases. In Modern Soil Microbiology; Van Elsas, JD, Trevors, JT, Wellington, EHM, Eds.; Marcel Dekker: New York, NY, USA, 1997; pp. 525–546. [Google Scholar]
- Amadioha, AC. Controlling rice blast in vitro and in vivo with extracts of Azadirachta indica. Crop Prot 2000, 19, 287–290. [Google Scholar]
- Ansari, MM. Control of sheath blight of rice by plant extracts. Indian Phytopathol 1995, 48, 268–270. [Google Scholar]
- Fridlender, M; Inbar, J; Chet, I. Biological control of soilborne plant pathogens by a β-1,3 glucanase-producing Pseudomonas cepacia. Soil Biol. Biochem 1993, 25, 1211–1221. [Google Scholar]
- Ramamoorthy, V; Raguchander, T; Samiyappan, R. Induction of defenserelated proteins in tomato roots treated with Pseudomonas fluorescens Pf1 and Fusarium oxysporum f. sp. lycopersici. Plant Soil 2002, 239, 55–68. [Google Scholar]
- Saravanakumar, D. PGPR-induced defense responses in the tea plant against blister blight disease. Crop Prot 2007, 26, 556–565. [Google Scholar]
- Sateesh, K. Antimicrobial activity and induction of systemic resistance in rice by leaf extract of Datura metel against Rhizoctonia solani and Xanthomonas oryzae pv. oryzae. Physiol. Mol. Plant Pathol 2004, 65, 91–100. [Google Scholar]
- Schneider, S; Ullrich, WR. Differential induction of resistance and enhanced enzyme activities in cucumber and tobacco caused by treatment with various abiotic and biotic inducers. Physiol. Mol. Plant Pathol 1994, 45, 291–304. [Google Scholar]
- Kloepper, JW; Lifshitz, R; Zablotowicz, RM. Free-living bacterial inocula for enhancing crop productivity. Trends Biotechnol 1989, 7, 39–43. [Google Scholar]
- Glick, BR. The enhancement of plant growth by free living bacteria. Can. J. Microbiol 1995, 41, 109–114. [Google Scholar]
- Okon, Y; Labandera-Gonzalez, CA. Agronomic Applications of Azospirillum. In Improving Plant Productivity with Rhizosphere Bacteria; Commonwealth Scientific and Industrial Research Organization: Adelaide, Australia, 1994; pp. 274–278. [Google Scholar]
- Fuhrmann, JJ; Wollum, AG, II. Nodulation competition among Bradyrhizobium japonicum strains as influenced by rhizosphere bacteria and iron availability. Biol. Fert. Soils 1989, 7, 108–112. [Google Scholar]
- Mahaffee, WF; Kloepper, JW. Applications of plant growth-promoting rhizobacteria in sustainable agriculture. In Soil Biota, Management in Sustainable Farming Systems; Pankhurst, CE, Doube, BM, Gupta, VVSR, Grace, PR, Eds.; CSIRO Information Services: Victoria, Australia, 1994; pp. 23–31. [Google Scholar]
- Alstrom, S. Induction of disease resistance in common bean susceptible to halo blight bacterial pathogen after seed bacterization with rhizosphere pseudomonads. J. Gen. Appl. Microbiol 1991, 37, 495–501. [Google Scholar]
- Graham, TL; Sequeira, L; Huang, TR. Bacterial lipopolysaccharides as inducers of disease resistance in tobacco. Appl. Environ. Microbiol 1977, 34, 424–432. [Google Scholar]
- Van Peer, R; Schippers, B. Lipopolysaccharides of plant-growth promoting Pseudomonas spp. strain WCS 417r induce resistance in carnation to Fusarium wilt. Neth. J. Plant Pathol 1992, 98, 129–139. [Google Scholar]
- Cattelan, AJ; Hartel, PG; Fuhrmann, JJ. Screening of plant growth promoting rhizobacteria to promote early soybean growth. Soil Sci. Soc. Am. J 1999, 63, 1670–1680. [Google Scholar]
- Chanway, CP; Holl, FB. First year yield performance of spruce seedlings inoculated with plant growth promoting rhizobacteria. Can. J. Microbiol 1993, 39, 1084–1088. [Google Scholar]
- Zhender, GW. Microbe-induced resistance against pathogens and herbivores. evidence of effectiveness in agriculture. In Induced Plant Defenses Against Pathogens and Herbivores: Biochemistry, Ecology and Agriculture; Agarwal, AA, Tuzun, S, Bent, E, Eds.; APS Press: St Paul, MN, USA, 1999. [Google Scholar]
- Bent, E. Alterations in plant growth and in root hormone levels of lodgepole pines inoculated with rhizobacteria. Can. J. Microbiol 2001, 47, 793–800. [Google Scholar]
- Ahmad, F; Ahmad, I; Khan, MS. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res 2008, 163, 173–181. [Google Scholar]
- Liu, XL. Studies on tobacco rhizosphere microbes. Acta Ecologica Sinica 2006, 26, 552–557. [Google Scholar]
- Reche, MHLR; Fiuza, LM. Bacterial diversity in Ricefield water in Rio Grande do Sul. Braz. J. Microbiol 2005, 36, 253–257. [Google Scholar]
- Persing, DH. Diagnostic Molecular Microbiology Principles and Applications; ASM Press: Washington DC, USA, 1993. [Google Scholar]
- Marchesi, JR. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol 1998, 64, 795–799. [Google Scholar]
- Moyer, CL. A computer-simulated restriction fragment length polymorphism analysis of bacterial small-subunit rRNA genes: Efficacy of selected tetrameric restriction enzymes for studies of microbial diversity in nature. Appl. Environ. Microbiol 1996, 62, 2501–2507. [Google Scholar]
- Massol-Deya, AA; Odelson, DA; Hickey, RF; Tiedje, JM. Bacterial community fingerprinting of amplified 16S and 16–23S ribosomal DNA gene sequences and restriction endonuclease analysis (ARDRA). In Molecular Microbial Ecology Manual; Cluger Publishing Co: Boston, MA, USA, 1995; pp. 1–8. [Google Scholar]
- Kloepper, JW; John, L; Teintze, M. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar]
- Neilands, JB; Leong, SA. Siderophores in relation to plant growth and disease. Annu. Rev. Plant Physiol 1986, 37, 187–208. [Google Scholar]
- Berg, G. Rhizobacteria of oil seed rape antagonistic to Verticillium dahliae. J. Plant Dis. Prot 1996, 103, 20–30. [Google Scholar]
- Murray, MG; Thormpson, WF. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res 1980, 8, 4321–4326. [Google Scholar]
- Rademaker, JLW; Bruijn, FJD. Characterization and Classification of Microbes by REP-PCR Genomic Fingerprinting and Computer-Assisted Pattern Analysis. In DNA Markers: Protocols, Applications and Overviews; Caetano-Anolle’s, G, Gresshoff, PM, Eds.; J. Wiley and Sons, Inc: New York, NY, USA, 1997; pp. 151–171. [Google Scholar]
- Rohlf, F. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System; Version 1.80; Owner’s manual, Exter Software: New York, NY, USA, 1993. [Google Scholar]
- Laguerre, G. Rapid identification of Rhizobia by restriction fragment length polymorphism analysis of PCR-amplified 16S rRNA genes. Appl. Environ. Microbiol 1994, 60, 56–63. [Google Scholar]
- Tamura, K. MEGA4: Molecular evolutionary genetics analysis(MEGA)software, version 4.0. Mol. Biol. Evol 2007, 24, 1596–1599. [Google Scholar]
- Raaijmakers, JM; Weller, DM. Exploiting genotypic diversity of 2,4-diacetylphloroglucinol-producing Pseudomonas spp.: Characterization of superior root-colonizing P. fluorescens strain Q8r1–96. Appl. Environ. Microbiol 2001, 67, 2545–2554. [Google Scholar]
- Berg, G; Roskot, N; Smalla, K. Plant-Dependent Genotypic and Phenotypic Diversity of Antagonistic Rhizobacteria Isolated from Different Verticillium Host Plants. Appl. Environ. Microbiol 2002, 68, 3328–3338. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. | Isolate | Accession No. | Closest NCBI Strain and Accession No. | Similarity | |
|---|---|---|---|---|---|---|
| 1 | 1 | WCH1 | HQ143640 | Bacillus flexus GU339233 | 99% | |
| 2 | 12 | RHH1 | HQ143609 | Bacillus pumilus FJ763645 | 99% | |
| 3 | 21 | ZAH1 | HQ143664 | Bacillus subtilis GQ861470 | 99% | |
| 4 | 2 | TZH5 | HQ143624 | Bacillus licheniformis GQ375247 | 99% | |
| 5 | 57 | DZH2 | GQ375785 | Bacillus altitudinis HM582688 | 100% | |
| YQH26 | HQ143652 | Bacillus sp. HQ236080 | 99% | |||
| 6 | 46 | FGH1 | HQ143588 | Brevibacillus brevis AY591911 | 99% | |
| 7 | 13 | FGH26 | HQ143593 | Paenibacillus polymyxa AM062684 | 99% | |
| 8 | 18 | WCH7 | HQ202565 | Streptomyces sp. GU722183 | 99% | |
| TZH2 | HQ143622 | Streptomyces ryensis AB184517 | 99% | |||
| 9 | 13 | FGH4 | HQ143590 | Streptomyces scabiei HM018077 | 99% | |
| TZH3 | HQ143623 | Streptomyces globosus EU196532 | 99% | |||
| 10 | 15 | WCH16 | HQ143672 | Streptomyces roseolus AB184168 | 99% | |
| SYH8 | HQ143620 | Streptomyces tanashiensis FJ481625 | 99% | |||
| 11 | 4 | YQH21 | HQ143674 | Kocuria palustris EU379293 | 98% | |
| 12 | 17 | RHH45 | HQ143617 | Pseudomonas fluorescens HM439651 | 99% | |
| MTH24 | HQ143602 | Pseudomonas corrugata AF348508 | 99% | |||
| 13 | 8 | ZAH18 | HQ143666 | Pseudomonas aeruginosa HJ472851 | 100% | |
| 14 | 7 | WCH22 | HQ143645 | Pseudomonas stutzeri FJ959391 | 99% | |
| 15 | 4 | FGH19 | HQ143592 | Enterobacter cancerogenus HQ154134 | 99% | |
| 16 | 1 | YQH50 | HQ143657 | Serratia marcescens FJ360759 | 100% | |
| 17 | 7 | MTH23 | GQ375787 | Delftia tsuruhatensis HM003215 | 100% | |
| 18 | 4 | MTH11 | HQ143598 | Sinorhizobium adhaerens AJ420775 | 100% | |
| 19 | 3 | SYH16 | HQ143621 | Agrobacterium tumefaciens FN433082 | 99% | |
| 20 | 1 | YC05107 | EU430120 | Burkholderia arboris AB458219 | 99% | |
| 21 | 2 | RHH55 | GQ375790 | Alcaligenes faecalis HM145896 | 99% | |
| 22 | 1 | WCH18 | HQ143644 | Advenella incenata AM944735 | 100% | |
| 23 | 8 | MTH13 | HQ143600 | Lysobacter capsici FN357198 | 99% | |
| MTH26 | HQ143603 | Lysobacter panaciterrae AB245359 | 99% | |||
| 24 | 1 | MTH20 | HQ202543 | Stenotrophomonas maltophilia FN645727 | 99% | |
| 25 | 1 | RHH50 | HQ202548 | Myroides odoratimimus EU373431 | 99% | |
| Total | 267 | |||||
| Genus | Species | Numbers of Strains | Percentage1 | Percentage2 |
|---|---|---|---|---|
| Bacillus | Bacillus altitudinis | 36 | 13.48% | 34.83% |
| Bacillus sp. | 21 | 7.87% | ||
| Bacillus subtilis | 21 | 7.87% | ||
| Bacillus pumilus | 12 | 4.49% | ||
| Bacillus flexus | 1 | 0.37% | ||
| Bacillus licheniformis | 2 | 0.75% | ||
| Brevibacillus | Brevibacillus brevis | 46 | 17.23% | 17.23% |
| Paenibacillus | Paenibacillus polymyxa | 13 | 4.87% | 4.87% |
| Streptomyces | Streptomyces sampsonii | 11 | 4.12% | 17.23% |
| Streptomyces roseolus | 3 | 1.12% | ||
| Streptomyces globosus | 6 | 2.25% | ||
| Streptomyces ryensis | 2 | 0.75% | ||
| Streptomyces scabies | 5 | 1.87% | ||
| Streptomyces tanashiensis | 6 | 2.25% | ||
| Streptomyces sp. | 13 | 4.87% | ||
| Kocuria | Kocuria palustris | 4 | 1.50% | 1.50% |
| Pseudomonas | Pseudomonas aeruginosa | 9 | 3.37% | 11.99% |
| Pseudomonas stutzeri | 7 | 2.62% | ||
| Pseudomonas corrugate | 6 | 2.25% | ||
| Pseudomonas fluorescens | 5 | 1.87% | ||
| Pseudomonas sp. | 5 | 1.87% | ||
| Enterobacter | Enterobacter cancerogenus | 2 | 0.75% | 1.50% |
| Enterobacter asburiae | 2 | 0.75% | ||
| Serratia | Serratia marcescens | 1 | 0.37% | 0.37% |
| Delftia | Delftia tsuruhatensis | 3 | 1.12% | 2.62% |
| Delftia lacustris | 1 | 0.37% | ||
| Delftia sp. | 3 | 1.12% | ||
| Burkholderia | Burkholderia arboris | 1 | 0.37% | 0.37% |
| Alcaligenes | Alcaligenes faecalis | 2 | 0.75% | 0.75% |
| Advenella | Advenella incenata | 1 | 0.37% | 0.37% |
| Lysobacter | Lysobacter enzymogenes | 3 | 1.12% | 3.00% |
| Lysobacter capsici | 2 | 0.75% | ||
| Lysobacter panaciterrae | 3 | 1.12% | ||
| Stenotrophomonas | Stenotrophomonas maltophilia | 1 | 0.37% | 0.37% |
| Sinorhizobium | Sinorhizobium adhaerens | 4 | 1.50% | 1.50% |
| Agrobacterium | Agrobacterium tumefaciens | 3 | 1.12% | 1.12% |
| Myroides | Myroides odoratimimus | 1 | 0.37% | 0.37% |
| Total | 267 | 100% | 100% |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jin, F.; Ding, Y.; Ding, W.; Reddy, M.S.; Fernando, W.G.D.; Du, B. Genetic Diversity and Phylogeny of Antagonistic Bacteria against Phytophthora nicotianae Isolated from Tobacco Rhizosphere. Int. J. Mol. Sci. 2011, 12, 3055-3071. https://doi.org/10.3390/ijms12053055
Jin F, Ding Y, Ding W, Reddy MS, Fernando WGD, Du B. Genetic Diversity and Phylogeny of Antagonistic Bacteria against Phytophthora nicotianae Isolated from Tobacco Rhizosphere. International Journal of Molecular Sciences. 2011; 12(5):3055-3071. https://doi.org/10.3390/ijms12053055
Chicago/Turabian StyleJin, Fengli, Yanqin Ding, Wei Ding, M.S. Reddy, W.G. Dilantha Fernando, and Binghai Du. 2011. "Genetic Diversity and Phylogeny of Antagonistic Bacteria against Phytophthora nicotianae Isolated from Tobacco Rhizosphere" International Journal of Molecular Sciences 12, no. 5: 3055-3071. https://doi.org/10.3390/ijms12053055
APA StyleJin, F., Ding, Y., Ding, W., Reddy, M. S., Fernando, W. G. D., & Du, B. (2011). Genetic Diversity and Phylogeny of Antagonistic Bacteria against Phytophthora nicotianae Isolated from Tobacco Rhizosphere. International Journal of Molecular Sciences, 12(5), 3055-3071. https://doi.org/10.3390/ijms12053055