Actinopyga lecanora Hydrolysates as Natural Antibacterial Agents

Abstract
:1. Introduction
2. Results and Discussion
2.1. Proximate Composition
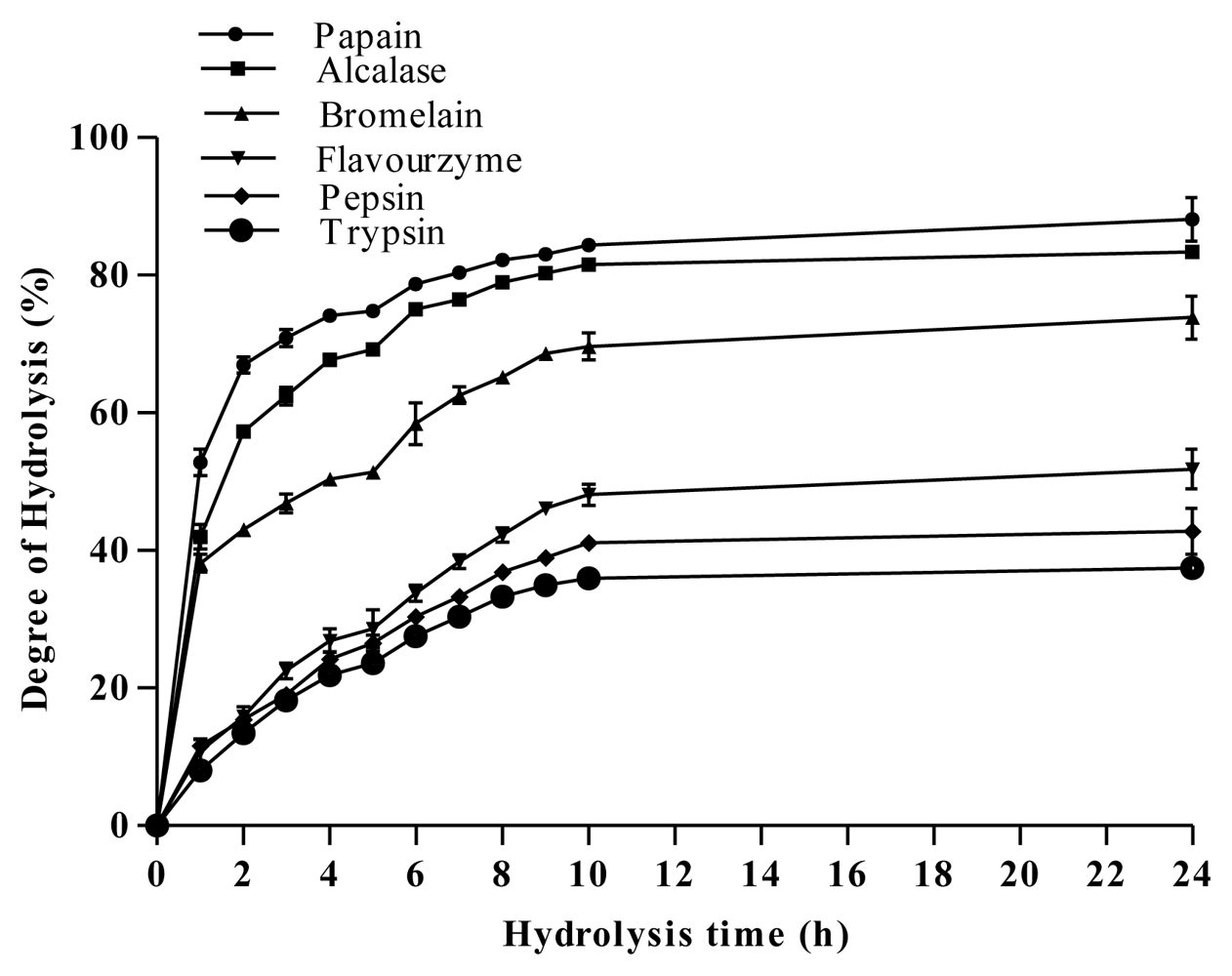
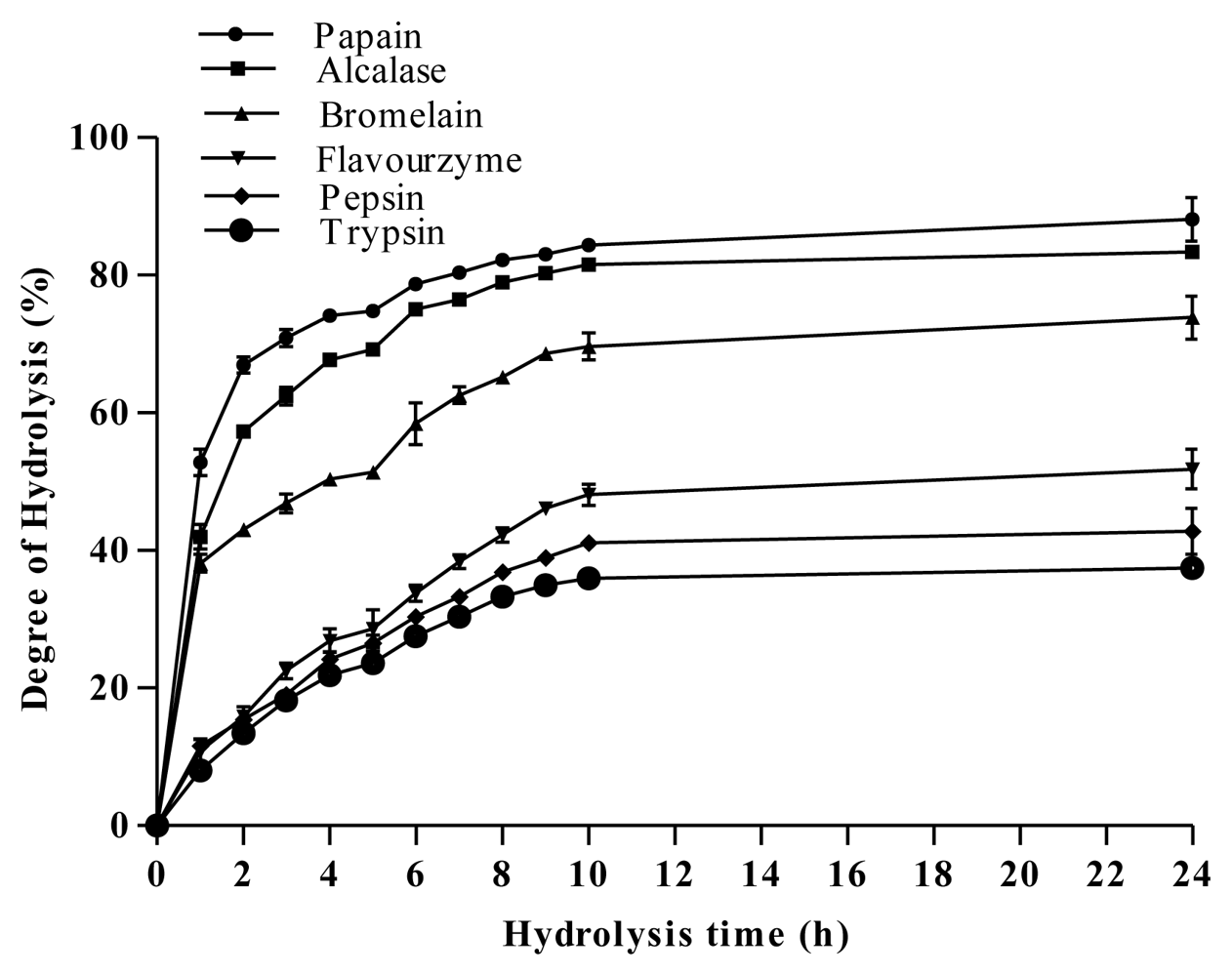
2.2. Degree of Hydrolysis
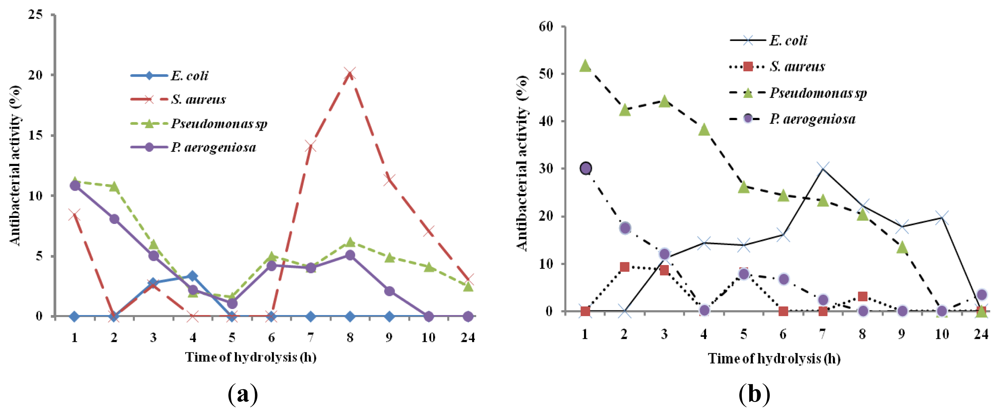
2.3. Antibacterial Activity of Hydrolysates
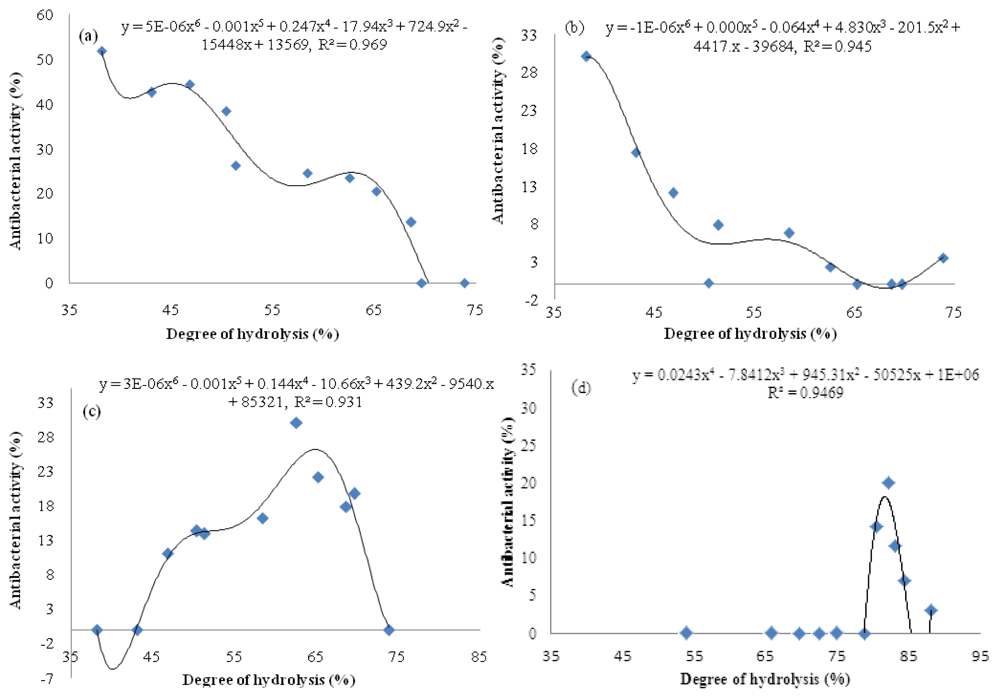
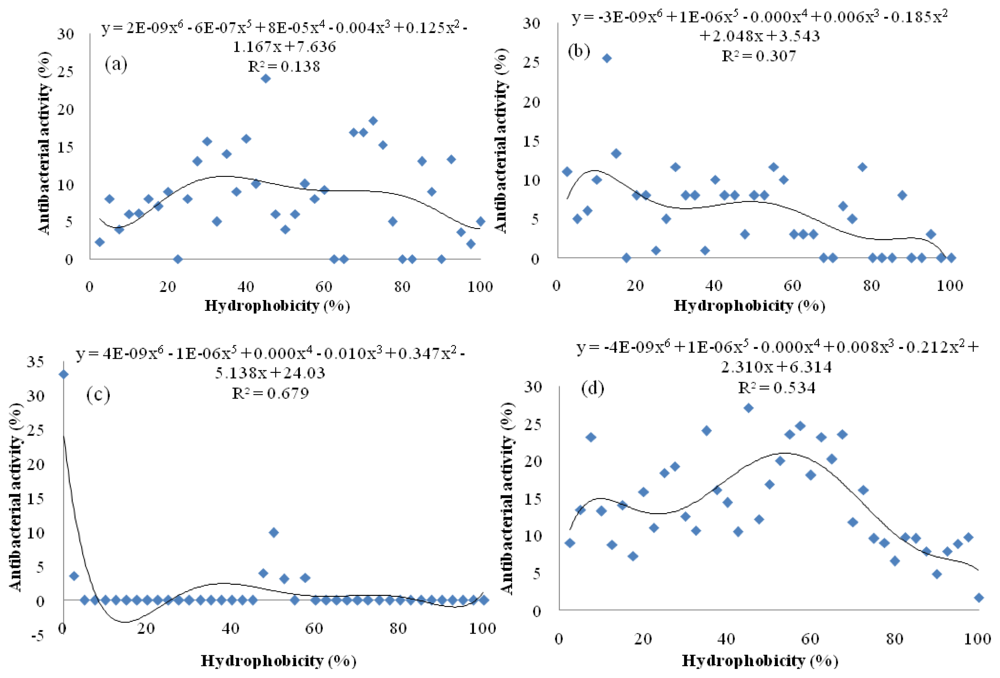
2.4. Relationship between Degree of Hydrolysis and Antibacterial Activity
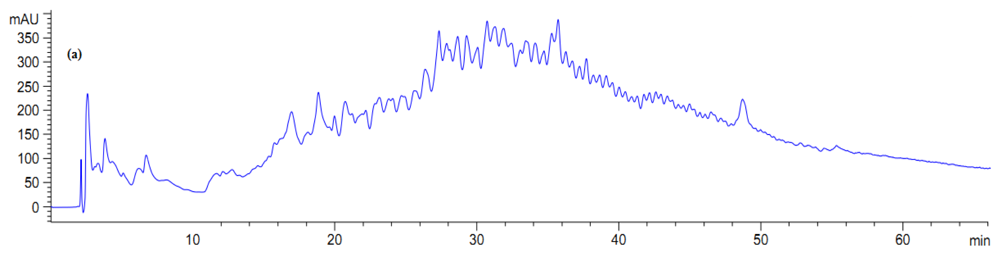
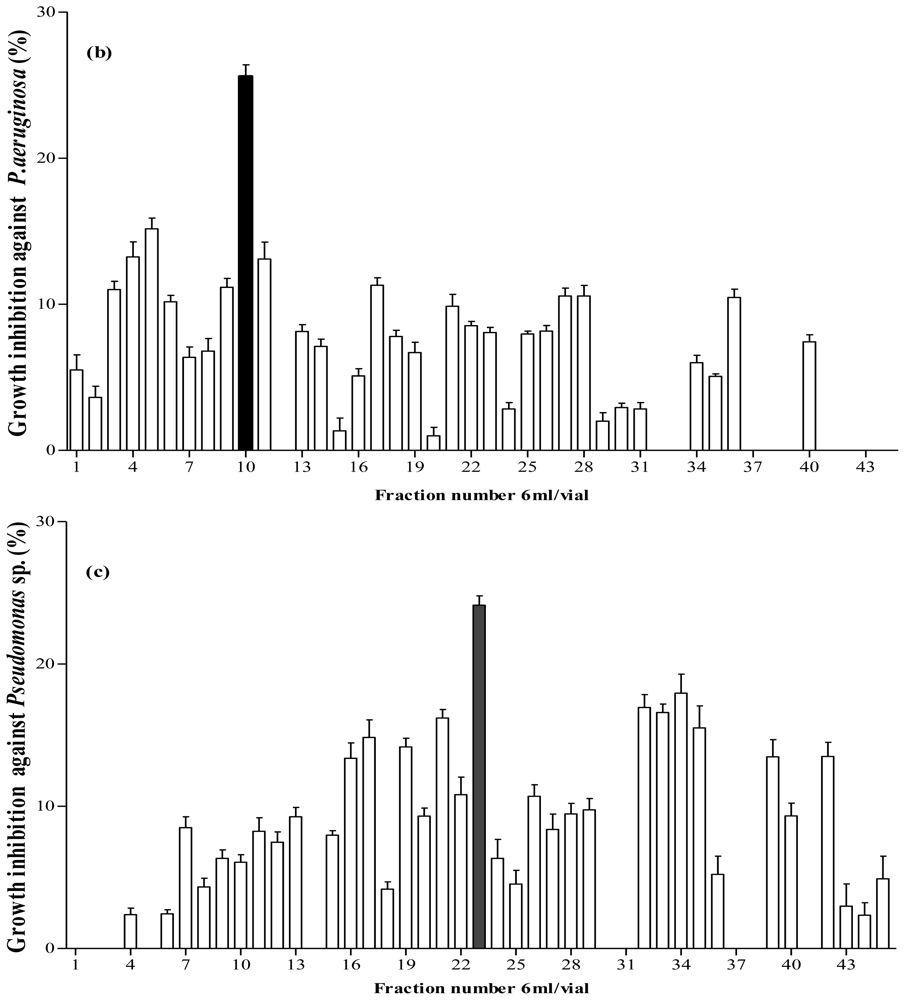
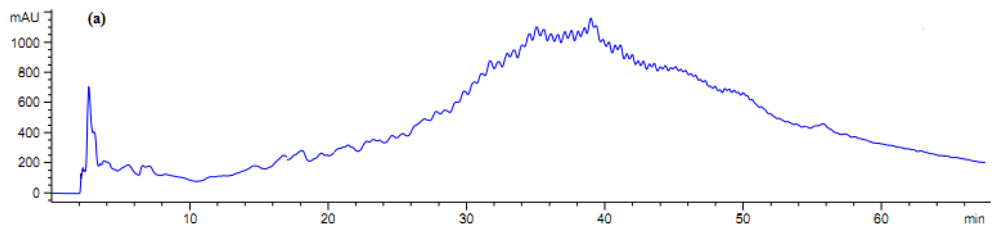
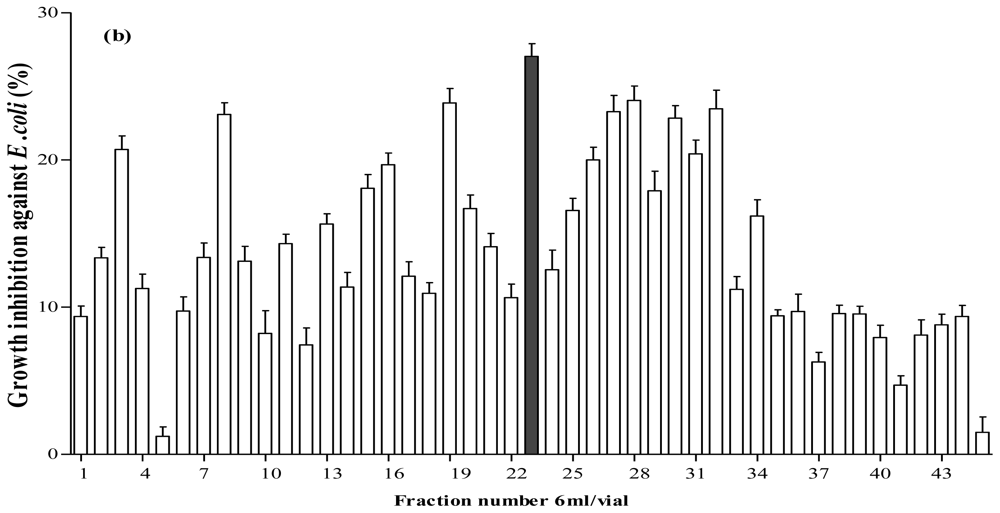
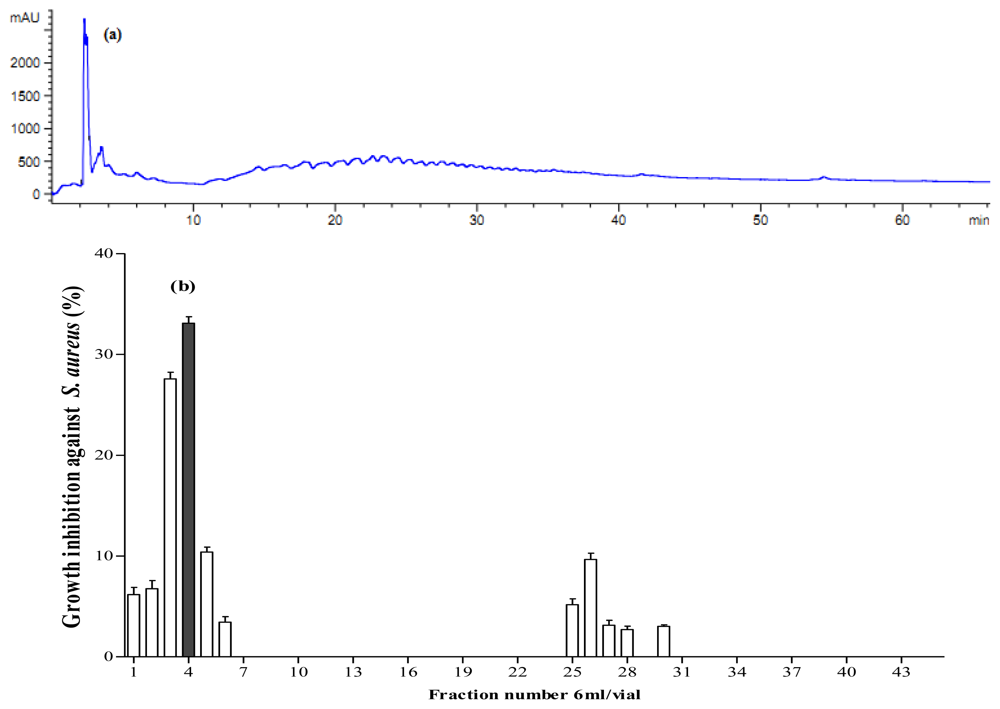
2.5. Antibacterial Activity of Fractionated Peptides
3. Experimental Section
3.1. Materials and Chemicals
3.2. Samples
3.3. Proximate Analysis
3.4. Preparation of A. lecanora Protein Hydrolysate
3.5. Determination of the Degree of Hydrolysis
3.6. Cultivation and Bacterial Inoculum Perpetration
3.7. Determination of Antibacterial Activity
3.8. Fractionation of A. lecanora Peptide
4. Statistical Analysis
5. Conclusion
Acknowledgements
References
- Hong, S.Y.; Park, T.G.; Lee, K.-H. The effect of charge increase on the specificity and activity of a short antimicrobial peptide. Peptides 2001, 22, 1669–1674. [Google Scholar]
- Vercruysse, L.; van Camp, J.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem 2005, 53, 8106–8115. [Google Scholar]
- Peña-Ramos, E.A.; Xiong, Y.L. Antioxidant activity of soy protein hydrolysates in a liposomal system. J. Food Sci 2002, 67, 2952–2956. [Google Scholar]
- Davalos, A.; Miguel, M.; Bartolome, B.; Lopez-Fandino, R. Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis. J. Food Prot 2004, 67, 1939–1944. [Google Scholar]
- Divakaramenon, V.; David, K.; Antoine, H.S.; Jayendra, B.B.; Richard, B.; Allen, M.; Amanda, L.R.; Brittany, C.W.; Rickey, P.H. Novel antimicrobial peptides that exhibit activity against select agents and other drug resistant bacteria. Bioorg. Med. Chem 2010, 18, 5137–5147. [Google Scholar]
- Arenas, G.; Guzman, F.; Cardenas, C.; Mercado, L.; Marshall, S.H. A novel antifungal peptide designed from the primary structure of a natural antimicrobial peptide purified from Argopecten purpuratus hemocytes. Peptides 2009, 30, 1405–1411. [Google Scholar]
- Battison, A.L.; Summerfield, R.; Patrzykat, A. Isolation and characterisation of two antimicrobial peptides from haemocytes of the American lobster Homarus americanus. Fish Shellfish Immun 2008, 25, 181–187. [Google Scholar]
- Liu, Z.; Dong, S.; Xu, J.; Zeng, M.; Song, H.; Zhao, Y. Production of cysteine-rich antimicrobial peptide by digestion of oyster (Crassostrea gigas) with alcalase and bromelin. Food Control 2008, 19, 231–235. [Google Scholar]
- Bartlett, T.C.; Cuthbertson, B.J.; Shepard, E.F.; Chapman, R.W.; Gross, P.S.; Warr, G.W. Crustins, homologues of an 11.5-kDa antibacterial peptide, from two species of penaeid shrimp, Litopenaeus vannamei and Litopenaeus setiferus. Mar. Biol 2002, 4, 278–293. [Google Scholar]
- Chun, L.; Tor, H.; Olaf, B.S.; Trond, O.J.; Klara, S. Strongylocins, novel antimicrobial peptides from the green sea urchin. Dev. Comp. Immunol 2008, 32, 1430–1440. [Google Scholar]
- Mingyong, Z.; Wenxuan, C.; Yuanhui, Z.; Zunying, L.; Shiyuan, D.; Yao, G. Antiviral active peptide from oyster. Chin. J. Oceanol. Limnol 2008, 23, 307–312. [Google Scholar]
- Zhong, Y.; Ahmad Khan, M.; Shahidi, F. Compositional characteristics and antioxidant properties of fresh and processed sea cucumber (Cucumaria frondosa). J. Agric. Food Chem 2007, 55, 1188–1192. [Google Scholar]
- Liu, H.M.; Cai, C.G.; Zhan, J.M. Using sea cucumber to treat 10 cases of anaemia. Guangxi Chin. Trad. Med 1984, 7, 18. [Google Scholar]
- Fredalina, B.D.; Ridzwan, B.H.; Zainal Abdin, A.A.; Kaswandi, M.A.; Zaiton, H.; Zali, I.; Kittakoop, P.; Jais, A.M. Fatty acid composition in local sea cucumber, Stichopus chloronotus for wound healing. Gen. Pharmacol 1999, 33, 337–340. [Google Scholar]
- Althunibat, O.Y.; Hashim, R.; Taher, M.; Daud, J.M.; Ikeda, M.-A.; Zali, B.I. In vitro antioxidant and antiproliferative activities of three malaysian sea cucumber species. Eur. J. Sci. Res 2009, 37, 376–387. [Google Scholar]
- Ridzwan, B.H.; Kaswandi, M.A.; Azman, Y.; Fuad, M. Screen for antibacterial agents in three species of sea cucumber from coastal areas of sabah. Gen. Pharmacol 1995, 26, 1539–1543. [Google Scholar]
- Ridzwan, B.H.; Zarina, M.Z.; Nadirah, M.; Kaswandi, M.A.; Shmsuddin, A.F. The antinociceptive effects of extracts from Stichopus chloronotus Brandt. Pakistan J. Biol. Sci 2001, 4, 244–246. [Google Scholar]
- Hing, H.L.; Kaswandi, M.A.; Azraul-Mumtazah, R.; Hamidah, S.A.; Sahalan, A.Z.; Normalawati, S.; Samsudin, M.W.; Ridzwan, B.H. Effect of methanol extracts from sea cucumbers Holothuria edulis and Stichopus chloronotus on candida albicans. Microsc. Microanal 2007, 13, 270–275. [Google Scholar]
- Kerr, R.; Chen, Z. In vivo and in vitro biosynthesis of saponins in sea cucumbers (Holothuroidea). J. Nat. Prod 1995, 58, 172–176. [Google Scholar]
- Miyamoto, T.; Togawa, K.; Higuchi, R.; Komori, T.; Sasaki, T. Constituents of Holothuroidea, II. Six newly identified biologically active triterpenoid glycoside sulfates from the sea cucumber Cucumaria Echinata. Eur. J. Org. Chem 1990, 1990, 453–460. [Google Scholar]
- Aminin, D.L.; Chaykina, E.L.; Agafonova, I.G.; Avilov, S.A.; Kalinin, V.I.; Stonik, V.A. Antitumor activity of the immunomodulatory lead Cumaside. Int. Immunopharmacol 2010, 10, 648–654. [Google Scholar]
- Vieira, R.P.; Mulloy, B.; Mourão, P.A. Structure of a fucose-branched chondroitin sulphate from sea cucumber. Evidence for the presence of 3-O-sulfo-β-d-glucuronosyl residues. J. Biol. Chem 1991, 266, 13530–13536. [Google Scholar]
- Nagase, H.; Enjyoji, K.; Minamiguchi, K.; Kitazato, K.T.; Kitazato, K.; Saito, H.; Kato, H. Depolymerized holothurian glycosaminoglycan with novel anticoagulant actions: Antithrombin III and heparin cofactor II-independent inhibition of factor X activation by factor IXa-factor VIIIa complex and heparin cofactor II-dependent inhibition of thrombin. Blood 1995, 85, 1527–1534. [Google Scholar]
- Pacheco, R.G.; Vicente, C.P.; Zancan, P.; Mourão, P.A.S. Different antithrombotic mechanisms among glycosaminoglycans revealed with a new fucosylated chondroitin sulfate from an Echinoderm. Blood Coagul. Fibrinolysis 2000, 11, 563–573. [Google Scholar]
- Mourao, P.A.S.; Pereira, M.S. Searching for alternatives to heparin: Sulfated fucans from marine invertebrates. Trends Cardiovasc. Med 1999, 9, 225–232. [Google Scholar]
- Goad, L.J.; Garneau, F.X.; Simard, J.L.; Apsimon, J.W.; Girard, M. Isolation of Δ9(11)-sterols from the sea cucumber, Implications for holothurin biosynthesis. Tetrahedron Lett 1985, 26, 3513–3516. [Google Scholar]
- Mamelona, J.; Pelletier, E.M.; Lalancette, K.G.; Legault, J.; Karboune, S.; Kermasha, S. Quantification of phenolic contents and antioxidant capacity of atlantic sea cucumber, Cucumaria frondosa. Food Chem 2007, 104, 1040–1047. [Google Scholar]
- Rafiuddin Ahmed, M.; Venkateshwarlu, U.; Jayakumar, R. Multilayered peptide incorporated collagen tubules for peripheral nerve repair. Biomaterials 2004, 25, 85–94. [Google Scholar]
- Sugawara, T.; Zaima, N.; Yamamoto, A.; Sakai, S.; Noguchi, R.; Hirata, T. Isolation of sphingoid bases of sea cucumber cerberosides and their cytotoxicity against human colon cancer cells. Biosci. Biotechnol. Biochem 2006, 70, 2906–2912. [Google Scholar]
- Mojica, E.R.E.; Merca, F.E. Lectin from the body walls of black sea cucumber (Holothuria atra Jaeger). Philippine J. Sci 2004, 133, 77–85. [Google Scholar]
- Mojica, E.R.E.; Merca, F.E. Biological properties of lectin from sea cucumber (Holothuria scabra Jaeger). J. Biol. Sci 2005, 5, 472–477. [Google Scholar]
- Jiaxin, C. Overview of sea cucumber farming and sea ranching practices in China. SPC Beche-de-mer Inf. Bull 2003, 18, 18–23. [Google Scholar]
- Forghani, B.; Ebrahimpour, A.; Bakar, J.; Abdul-Hamid, A.; Hassan, Z.; Saari, N. Enzyme hydrolysates from Stichopus horrens as a new source for angiotensin converting enzyme inhibitory peptides. J. Evid. Based Complement. Altern. Med. 2012. [Google Scholar] [CrossRef]
- Change-Lee, M.V.; Price, R.J.; Lampila, L.E. Effect of processing on proximate composition and mineral content of sea cucumbers (Parastichopus spp.). J. Food Sci 1989, 54, 567–572. [Google Scholar]
- Zarei, M.; Ebrahimpour, A.; Abdul-Hamid, A.; Anwar, F.; Saari, N. Production of defatted palm kernel cake protein hydrolysate as a valuable source of natural antioxidants. Int. J. Mol. Sci 2012, 13, 8097–8111. [Google Scholar]
- Sarita D’Avila dos, S.; Vilasia, G.M.; Myriam, S.-M.; Carlos, P. Evaluation of functional properties in protein hydrolysates from bluewing searobin (Prionotus punctatus) obtained with different microbial enzymes. Food Bioprocess Technol 2011, 4, 1399–1406. [Google Scholar]
- Song, R.; Wei, R.; Zhang, B.; Wang, D. Optimization of the antibacterial activity of Half-Fin Anchovy (Setipinna taty) hydrolysates. Food Bioprocess Technol 2012, 5, 1979–1989. [Google Scholar]
- Salampessy, J.; Phillips, M.; Seneweera, S.; Kailasapathy, K. Release of antimicrobial peptides through bromelain hydrolysis of leatherjacket (Meuchenia sp.) insoluble proteins. Food Chem 2010, 120, 556–560. [Google Scholar]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar]
- Jang, A.; Cheorun, J.; Kyung-Sun, K.; Mooha, L. Antimicrobial and human cancer cell cytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory peptides. Food Chem 2008, 107, 327–336. [Google Scholar]
- Association of Official Analytical Chemists (AOAC), Official Methods of Analysis, 15th ed; Helrich, K. (Ed.) AOAC: Arlington, VA, USA, 1990.
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci 1983, 66, 1219–1227. [Google Scholar]
- Penasa, E.; Prestamoa, G.; Gomezb, R. High pressure and the enzymatic hydrolysis of soybean whey proteins. Food Chem 2004, 85, 641–648. [Google Scholar]
- Patgaonkar, M.; Aranha, C.; Bhonde, G.; Reddy, K.V.R. Identification and characterization of anti-microbial peptides from rabbit vaginal fluid. Vet. Immunol. Immunop 2011, 139, 176–186. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Amount (%) | |
|---|---|---|
| aA. lecanora | Other species | |
| Moisture | 87.96 ± 0.49 | 82–92.6 |
| Protein | 7.03 ± 0.16 | 2.5–13.8 |
| Fat | 0.65 ± 0.08 | 0.l–0.9 |
| Ash | 2.93 ± 0.07 | l.5–4.3 |
| Carbohydrate | 1.43 ± 0.09 | 0–2.2 |
| Enzyme | Buffer (50 mM) | pH | Temperature (°C ) | Agitation rate (rpm) |
|---|---|---|---|---|
| Papain | Phosphat | 7 | 60 | 150 |
| Pepsin | KCl-HCl | 1.5 | 37 | 150 |
| Trypsin | Borate | 8 | 37 | 150 |
| Alcalase | Borate | 8 | 55 | 150 |
| Bromelain | Acetate | 5.5 | 55 | 150 |
| Flavourzyme | Phosphat | 6.5 | 55 | 150 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ghanbari, R.; Ebrahimpour, A.; Abdul-Hamid, A.; Ismail, A.; Saari, N. Actinopyga lecanora Hydrolysates as Natural Antibacterial Agents. Int. J. Mol. Sci. 2012, 13, 16796-16811. https://doi.org/10.3390/ijms131216796
Ghanbari R, Ebrahimpour A, Abdul-Hamid A, Ismail A, Saari N. Actinopyga lecanora Hydrolysates as Natural Antibacterial Agents. International Journal of Molecular Sciences. 2012; 13(12):16796-16811. https://doi.org/10.3390/ijms131216796
Chicago/Turabian StyleGhanbari, Raheleh, Afshin Ebrahimpour, Azizah Abdul-Hamid, Amin Ismail, and Nazamid Saari. 2012. "Actinopyga lecanora Hydrolysates as Natural Antibacterial Agents" International Journal of Molecular Sciences 13, no. 12: 16796-16811. https://doi.org/10.3390/ijms131216796
APA StyleGhanbari, R., Ebrahimpour, A., Abdul-Hamid, A., Ismail, A., & Saari, N. (2012). Actinopyga lecanora Hydrolysates as Natural Antibacterial Agents. International Journal of Molecular Sciences, 13(12), 16796-16811. https://doi.org/10.3390/ijms131216796