Mitogen-Activated Protein (MAP) Kinases in Plant Metal Stress: Regulation and Responses in Comparison to Other Biotic and Abiotic Stresses


Abstract
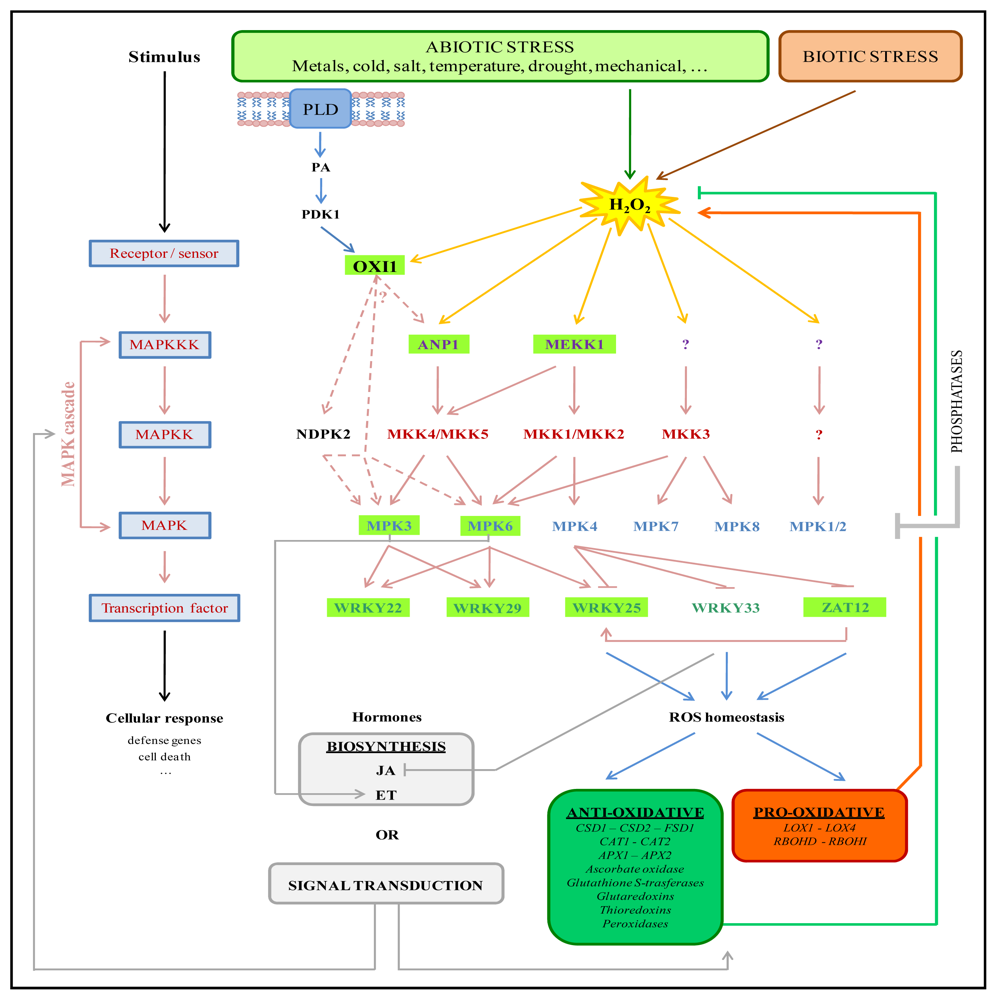
:1. Introduction
2. MAPKinases: Nomenclature and Classification
3. Regulation of MAPKinases
3.1. Scaffolding
3.2. Phosphatases
3.3. Lipid Signaling
4. Role of MAPK Cascades in Stress Response Signaling
4.1. MAPK Cascades Are Involved in Metal Stress
4.2. Comparison to MAPK Pathways Involved in Other Stress Responses
5. MAPK Cascades Regulate Stress Responses by Activation of Gene Transcription
5.1. Transcription Factors
5.2. Regulation of the Cellular Redox Status by MAPK Cascades
5.3. MAPK Cascades Interfere with Hormone Signaling
5.3.1. Ethylene
5.3.2. Jasmonate
6. Conclusions
Acknowledgments
References
- Nriagu, J.O.; Pacyna, J.M. Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature 1988, 333, 134–139. [Google Scholar]
- Ruttens, A.; Boulet, J.; Weyens, N.; Smeets, K.; Adriaensen, K.; Meers, E.; van Slycken, S.; Tack, F.; Meiresonne, L.; Thewys, T.; Witters, N.; Carleer, R.; Dupae, J.; Vangronsveld, J. Short rotation coppice culture of willow and poplar as energy crops on metal contaminated agricultural soils. Int. J. Phytoremediat 2011, 13, 194–207. [Google Scholar]
- Kirkham, M.B. Cadmium in plants on polluted soils: Effects of soil factors, hyperaccumulation, and amendments. Geoderma 2006, 137, 19–32. [Google Scholar]
- Chary, N.S.; Kamala, C.T.; Raj, D.S.S. Assessing risk of heavy metals from consuming food grown on sewage irrigated soils and food chain transfer. Ecotoxicol. Environ. Saf 2008, 69, 513–524. [Google Scholar]
- Marschner, H. Functions of Mineral Nutrients: Micronutrients. In Mineral Nutrition of Higher Plants, 2nd ed; Marschner, H., Ed.; Academic Press: London, UK, 1995; pp. 313–404. [Google Scholar]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol 2007, 173, 677–702. [Google Scholar]
- Cuypers, A.; Smeets, K.; Vangronsveld, J. Heavy Metal Stress in Plants. In Plant Stress Biology: From Genomics to Systems Biology, 1st ed; Hirt, H., Ed.; Wiley-VCH Verlag: Weinheim, Germany, 2009; pp. 161–178. [Google Scholar]
- Leonard, S.S.; Bower, J.J.; Shi, X. Metal-induced toxicity, carcinogenesis, mechanisms and cellular responses. Mol. Cell. Biochem 2004, 255, 3–10. [Google Scholar]
- Hogervorst, J.; Plusquin, M.; Vangronsveld, J.; Nawrot, T.; Cuypers, A.; van Hecke, E.; Roels, H.A.; Carleer, R.; Staessen, J.A. House dust as possible route of environmental exposure to cadmium and lead in the adult general population. Environ. Res 2007, 103, 30–37. [Google Scholar]
- Nawrot, T.; Plusquin, M.; Hogervorst, J.; Roels, H.A.; Celis, H.; Thijs, L.; Vangronsveld, J.; van Hecke, E.; Staessen, J.A. Environmental exposure to cadmium and risk of cancer: A prospective population-based study. Lancet Oncol 2006, 7, 119–226. [Google Scholar]
- Nawrot, T.; Staessen, J.A.; Roels, H.; Munters, E.; Cuypers, A.; Richart, T.; Ruttens, A.; Smeets, K.; Clijsters, H.; Vangronsveld, J. Cadmium exposure in the population: from risks to measures of prevention. Biometals 2010, 23, 769–782. [Google Scholar]
- Thijssen, S.; Cuypers, A.; Maringwa, J.; Smeets, K.; Horemans, N.; Lambrichts, I.; van Kerkhove, E. Low cadmium exposure triggers a biphasic oxidative stress response in mice kidneys. Toxicology 2007, 236, 29–41. [Google Scholar]
- DalCorso, G.; Farinati, S.; Maistri, S.; Furini, A. How plants cope with cadmium: Staking all on metabolism and gene expression. J. Integr. Plant Biol 2008, 50, 1268–1280. [Google Scholar]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol 2009, 36, 409–430. [Google Scholar]
- Smeets, K.; Opdenakker, K.; Remans, T.; van Sanden, S.; van Belleghem, F.; Semane, B.; Horemans, N.; Guisez, Y.; Vangronsveld, J.; Cuypers, A. Oxidative stress-related responses at transcriptional and enzymatic levels after exposure to Cd or Cu in a multipollution context. J. Plant Physiol 2009, 166, 1982–1992. [Google Scholar]
- Cuypers, A.; Smeets, K.; Ruytinx, J.; Opdenakker, K.; Keunen, E.; Remans, T.; Horemans, N.; Vanhoudt, N.; van Sanden, S.; van Belleghem, F.; Guisez, Y.; Colpaert, J.; Vangronsveld, J. The cellular redox state as a modulator in cadmium and copper responses in Arabidopsis thaliana seedlings. J. Plant Physiol 2011, 168, 309–316. [Google Scholar]
- Cuypers, A.; Keunen, E.; Bohler, S.; Jozefczak, M.; Opdenakker, K.; Gielen, H.; Vercampt, H.; Bielen, A.; Schellingen, K.; Vangronsveld, J.; Remans, T. Cadmium and Copper Stress Induce a Cellular Oxidative Challenge Leading to Damage versus Signalling. In Metal Toxicity in Plants: Perception, Signalling and Remediation, 1st ed; Gupta, D.K.G., Sandalio, L.M., Eds.; Springer-Verlag GmbH: Berlin, Germany, 2011; pp. 65–90. [Google Scholar]
- Gratao, P.L.; Polle, A.; Lea, P.J.; Azevedo, R.A. Making the life of heavy metal-stressed plants a little easier. Funct. Plant Biol 2005, 32, 481–494. [Google Scholar]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci 2004, 9, 490–498. [Google Scholar]
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive oxygen species signaling in plants. Antioxid. Redox Sign 2006, 8, 1757–1764. [Google Scholar]
- Opdenakker, K.; Remans, T.; Keunen, E.; Vangronsveld, J.; Cuypers, A. Exposure of Arabidopsis thaliana to Cd or Cu excess leads to oxidative stress mediated alterations in MAPKinase transcript levels. Environ. Exp. Bot 2012, 83, 53–61. [Google Scholar]
- Kalbina, I.; Strid, A. The role of NADPH oxidase and MAP kinase phosphatase in UV-B-dependent gene expression in Arabidopsis. Plant Cell Environ 2006, 29, 1783–1793. [Google Scholar]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ 2010, 33, 453–467. [Google Scholar]
- Vandenabeele, S.; Vanderauwera, S.; Vuylsteke, M.; Rombauts, S.; Langebartels, C.; Seidlitz, H.K.; Zabeau, M.; van Montagu, M.; Inze, D.; van Breusegem, F. Catalase deficiency drastically affects gene expression induced by high light in Arabidopsis thaliana. Plant J 2004, 39, 45–58. [Google Scholar]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant 2006, 126, 45–51. [Google Scholar]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol 2006, 141, 373–378. [Google Scholar]
- Rogers, H.J. Is there an important role for reactive oxygen species and redox regulation during floral senescence? Plant Cell Environ 2012, 35, 217–233. [Google Scholar]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J 2006, 45, 113–122. [Google Scholar]
- De Pinto, M.C.; Locato, V.; de Gara, L. Redox regulation in plant programmed cell death. Plant Cell Environ 2012, 35, 234–244. [Google Scholar]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; Davies, J.M.; Dolan, L. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Miller, G.; Shulaev, V.; Mittler, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant 2008, 133, 481–489. [Google Scholar]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen peroxide in plants: A versatile molecule of the reactive oxygen species network. J. Integr. Plant Biol 2008, 50, 2–18. [Google Scholar]
- Mishra, N.S.; Tuteja, R.; Tuteja, N. Signaling through MAP kinase networks in plants. Arch. Biochem. Biophys 2006, 452, 55–68. [Google Scholar]
- Ichimura, K.; Shinozaki, K.; Tena, G.; Sheen, J.; Henry, Y.; Champion, A.; Kreis, M.; Zhang, S.Q.; Hirt, H.; Wilson, C.; Heberle-Bors, E.; Ellis, B.E.; Morris, P.C.; Innes, R.W.; Ecker, J.R.; Scheel, D.; Klessig, D.F.; Machida, Y.; Mundy, J.; Ohashi, Y.; Walker, J.C. Mitogen-activated protein kinase cascades in plants: a new nomenclature. Trends Plant Sci 2002, 7, 301–308. [Google Scholar]
- Jonak, C.; Okresz, L.; Bogre, L.; Hirt, H. Complexity, cross talk and integration of plant MAP kinase signalling. Curr. Opin. Plant Biol 2002, 5, 415–424. [Google Scholar]
- Morrison, D.K.; Davis, R.J. Regulation of map kinase signaling modules by scaffold proteins in mammals. Annu. Rev. Cell Dev. Biol 2003, 19, 91–118. [Google Scholar]
- Nakagami, H.; Kiegerl, S.; Hirt, H. OMTK1, a novel MAPKKK, channels oxidative stress signaling through direct MAPK interaction. J. Biol. Chem 2004, 279, 26959–26966. [Google Scholar]
- Bartels, S.; Besteiro, M.A.G.; Lang, D.; Ulm, R. Emerging functions for plant MAP kinase phosphatases. Trends Plant Sci 2010, 15, 322–329. [Google Scholar]
- Ulm, R.; Revenkova, E.; di Sansebastiano, G.P.; Bechtold, N.; Paszkowski, J. Mitogen-activated protein kinase phosphatase is required for genotoxic stress relief in Arabidopsis. Gene Dev 2001, 15, 699–709. [Google Scholar]
- Ulm, R.; Ichimura, K.; Mizoguchi, T.; Peck, S.C.; Zhu, T.; Wang, X.; Shinozaki, K.; Paszkowski, J. Distinct regulation of salinity and genotoxic stress responses by Arabidopsis MAP kinase phosphatase 1. EMBO J 2002, 21, 6483–6493. [Google Scholar]
- Anderson, J.C.; Bartels, S.; Besteiro, M.A.G.; Shahollari, B.; Ulm, R.; Peck, S.C. Arabidopsis MAP kinase phosphatase 1 (AtMKP1) negatively regulates MPK6-mediated PAMP responses and resistance against bacteria. Plant J 2011, 67, 258–268. [Google Scholar]
- Besteiro, M.A.G.; Bartels, S.; Albert, A.; Ulm, R. Arabidopsis MAP kinase phosphatase 1 and its target MAP kinases 3 and 6 antagonistically determine UV-B stress tolerance, independent of the UVR8 photoreceptor pathway. Plant J 2011, 68, 727–737. [Google Scholar]
- Lee, K.; Song, E.H.; Kim, H.S.; Yoo, J.H.; Han, H.J.; Jung, M.S.; Lee, S.M.; Kim, K.E.; Kim, M.C.; Cho, M.J.; Chung, W.S. Regulation of MAPK phosphatase 1 (AtMKP1) by calmodulin in Arabidopsis. J. Biol. Chem 2008, 283, 23581–23588. [Google Scholar]
- Bartels, S.; Anderson, J.C.; Besteiro, M.A.G.; Carreri, A.; Hirt, H.; Buchala, A.; Metraux, J.P.; Peck, S.C.; Ulm, R. MAP kinase phosphatase1 and protein tyrosine phosphatase1 are repressors of salicylic acid synthesis and SNC1-mediated responses in Arabidopsis. Plant Cell 2009, 21, 2884–2897. [Google Scholar]
- Lee, J.S.; Ellis, B.E. Arabidopsis MAPK phosphatase 2 (MKP2) positively regulates oxidative stress tolerance and inactivates the MPK3 and MPK6 MAPKs. J. Biol. Chem 2007, 282, 25020–25029. [Google Scholar]
- Lumbreras, V.; Vilela, B.; Irar, S.; Sole, M.; Capellades, M.; Valls, M.; Coca, M.; Pages, M. MAPK phosphatase MKP2 mediates disease responses in Arabidopsis and functionally interacts with MPK3 and MPK6. Plant J 2010, 63, 1017–1030. [Google Scholar]
- Bargmann, B.O.; Munnik, T. The role of phospholipase D in plant stress responses. Curr. Opin. Plant Biol 2006, 9, 515–522. [Google Scholar]
- Yu, L.J.; Nie, J.N.; Cao, C.Y.; Jin, Y.K.; Yan, M.; Wang, F.Z.; Liu, J.; Xiao, Y.; Liang, Y.H.; Zhang, W.H. Phosphatidic acid mediates salt stress response by regulation of MPK6 in Arabidopsis thaliana. New Phytol 2010, 188, 762–773. [Google Scholar]
- Anthony, R.G.; Henriques, R.; Helfer, A.; Meszaros, T.; Rios, G.; Testerink, C.; Munnik, T.; Deak, M.; Koncz, C.; Bogre, L. A protein kinase target of a PDK1 signalling pathway is involved in root hair growth in Arabidopsis. EMBO J 2004, 23, 572–581. [Google Scholar]
- Rentel, M.C.; Lecourieux, D.; Ouaked, F.; Usher, S.L.; Petersen, L.; Okamoto, H.; Knight, H.; Peck, S.C.; Grierson, C.S.; Hirt, H.; Knight, M.R. OXI1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis. Nature 2004, 427, 858–861. [Google Scholar]
- Camehl, I.; Drzewiecki, C.; Vadassery, J.; Shahollari, B.; Sherameti, I.; Forzani, C.; Munnik, T.; Hirt, H.; Oelmuller, R. The OXI1 kinase pathway mediates Piriformospora indica-induced growth promotion in Arabidopsis. PLoS Pathog 2011, 7. [Google Scholar] [CrossRef]
- Anthony, R.G.; Khan, S.; Costa, J.; Pais, M.S.; Bogre, L. The Arabidopsis protein kinase PTI1–2 is activated by convergent phosphatidic acid and oxidative stress signaling pathways downstream of PDK1 and OXI1. J. Biol. Chem 2006, 281, 37536–37546. [Google Scholar]
- Forzani, C.; Carreri, A.; van Bentem, S.D.; Lecourieux, D.; Lecourieux, F.; Hirt, H. The Arabidopsis protein kinase Pto-interacting 1–4 is a common target of the oxidative signal-inducible 1 and mitogen-activated protein kinases. Febs J 2011, 278, 1126–1136. [Google Scholar]
- Moon, H.; Lee, B.; Choi, G.; Shin, S.; Prasad, D.T.; Lee, O.; Kwak, S.S.; Kim, D.H.; Nam, J.; Bahk, J.; Hong, J.C.; Lee, S.Y.; Cho, M.J.; Lim, C.O.; Yun, D.J. NDP kinase 2 interacts with two oxidative stress-activated MAPKs to regulate cellular redox state and enhances multiple stress tolerance in transgenic plants. Proc. Natl. Acad. Sci. USA 2003, 100, 358–363. [Google Scholar]
- Zhang, T.; Liu, Y.; Yang, T.; Zhang, L.; Xu, S.; Xue, L.; An, L. Diverse signals converge at MAPK cascades in plant. Plant Physiol. Biochem 2006, 44, 274–283. [Google Scholar]
- Pitzschke, A.; Hirt, H. Disentangling the Complexity of Mitogen-activated protein kinases and reactive oxygen species signaling. Plant Physiol 2009, 149, 606–615. [Google Scholar]
- Zhang, S.Q.; Klessig, D.F. MAPK cascades in plant defense signaling. Trends Plant Sci 2001, 6, 520–527. [Google Scholar]
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: A complex signalling network involved in multiple biological processes. Biochem. J 2008, 413, 217–226. [Google Scholar]
- Liu, X.M.; Kim, K.E.; Kim, K.C.; Nguyen, X.C.; Han, H.J.; Jung, M.S.; Kim, H.S.; Kim, S.H.; Park, H.C.; Yun, D.J.; Chung, W.S. Cadmium activates Arabidopsis MPK3 and MPK6 via accumulation of reactive oxygen species. Phytochemistry 2010, 71, 614–618. [Google Scholar]
- Rao, K.P.; Vani, G.; Kumar, K.; Wankhede, D.P.; Misra, M.; Gupta, M.; Sinha, A.K. Arsenic stress activates MAP kinase in rice roots and leaves. Arch. Biochem. Biophys 2011, 506, 73–82. [Google Scholar]
- Jonak, C.; Nakagami, H.; Hirt, H. Heavy metal stress. Activation of distinct mitogen-activated protein kinase pathways by copper and cadmium. Plant Physiol 2004, 136, 3276–3283. [Google Scholar]
- Kovtun, Y.; Chiu, W.L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. USA 2000, 97, 2940–2945. [Google Scholar]
- Opdenakker, K. Centre for Environmental Sciences, Hasselt University, Diepenbeek, Belgium. Arabidopsis thaliana oxi1 and mpk6 knockout seedlings are less sensitive to Cu exposure. Unpublished work. 2012. [Google Scholar]
- Roelofs, D.; Aarts, M.G.M.; Schat, H.; van Straalen, N.M. Functional ecological genomics to demonstrate general and specific responses to abiotic stress. Funct. Ecol 2008, 22, 8–18. [Google Scholar]
- Suzuki, N.; Koizumi, N.; Sano, H. Screening of cadmium-responsive genes in Arabidopsis thaliana. Plant Cell Environ 2001, 24, 1177–1188. [Google Scholar]
- Gupta, M.; Sharma, P.; Sarin, N.B.; Sinha, A.K. Differential response of arsenic stress in two varieties of Brassica juncea L. Chemosphere 2009, 74, 1201–1208. [Google Scholar]
- Agrawal, G.K.; Rakwal, R.; Iwahashi, H. Isolation of novel rice (Oryza sativa L.) multiple stress responsive MAP kinase gene, OsMSRMK2, whose mRNA accumulates rapidly in response to environmental cues. Biochem. Bioph. Res. Commun 2002, 294, 1009–1016. [Google Scholar]
- Agrawal, G.K.; Agrawal, S.K.; Shibato, J.; Iwahashi, H.; Rakwal, R. Novel rice MAP kinases OsMSRMK3 and OsWJUMK1 involved in encountering diverse environmental stresses and developmental regulation. Biochem. Bioph. Res. Commun 2003, 300, 775–783. [Google Scholar]
- Yeh, C.M.; Hsiao, L.J.; Huang, H.J. Cadmium activates a mitogen-activated protein kinase gene and MBP kinases in rice. Plant Cell Physiol 2004, 45, 1306–1312. [Google Scholar]
- Yeh, C.M.; Hung, W.C.; Huang, H.J. Copper treatment activates mitogen-activated protein kinase signalling in rice. Physiol. Plant 2003, 119, 392–399. [Google Scholar]
- Hung, W.C.; Huang, D.D.; Yeh, C.M.; Huang, H.J. Reactive oxygen species, calcium and serine/threonine phosphatase are required for copper-induced MAP kinase gene OsMAPK2, expression in rice. Plant Growth Regul 2005, 45, 233–241. [Google Scholar]
- Yeh, C.M.; Chien, P.S.; Huang, H.J. Distinct signalling pathways for induction of MAP kinase activities by cadmium and copper in rice roots. J. Exp. Bot 2007, 58, 659–671. [Google Scholar]
- Lin, C.W.; Chang, H.B.; Huang, H.J. Zinc induces mitogen-activated protein kinase activation mediated by reactive oxygen species in rice roots. Plant Physiol. Biochem 2005, 43, 963–968. [Google Scholar]
- Huang, T.L.; Huang, H.J. ROS and CDPK-like kinase-mediated activation of MAP kinase in rice roots exposed to lead. Chemosphere 2008, 71, 1377–1385. [Google Scholar]
- Tsai, T.M.; Huang, H.J. Effects of iron excess on cell viability and mitogen-activated protein kinase activation in rice roots. Physiol. Plant 2006, 127, 583–592. [Google Scholar]
- Wang, J.X.; Ding, H.D.; Zhang, A.; Ma, F.F.; Cao, J.M.; Jiang, M.Y. A novel mitogen-activated protein kinase gene in maize (Zea mays), ZmMPK3, is involved in response to diverse environmental cues. J. Integr. Plant Biol 2010, 52, 442–452. [Google Scholar]
- Ding, H.D.; Tan, M.P.; Zhang, C.; Zhang, Z.W.; Zhang, A.Y.; Kang, Y.J. Hexavalent chromium (VI) stress induces mitogen-activated protein kinase activation mediated by distinct signal molecules in roots of Zea mays L. Environ. Exp. Bot 2009, 67, 328–334. [Google Scholar]
- Ren, D.T.; Yang, H.P.; Zhang, S.Q. Cell death mediated by MAPK is associated with hydrogen peroxide production in Arabidopsis. J. Biol. Chem 2002, 277, 559–565. [Google Scholar]
- Nakagami, H.; Soukupova, H.; Schikora, A.; Zarsky, V.; Hirt, H. A mitogen-activated protein kinase kinase kinase mediates reactive oxygen species homeostasis in Arabidopsis. J. Biol. Chem 2006, 281, 38697–38704. [Google Scholar]
- Ichimura, K.; Mizoguchi, T.; Irie, K.; Morris, P.; Giraudat, J.; Matsumoto, K.; Shinozaki, K. Isolation of ATMEKK1 (a MAP kinase kinase kinase)—Interacting proteins and analysis of a MAP kinase cascade in Arabidopsis. Biochem. Bioph. Res. Commun 1998, 253, 532–543. [Google Scholar]
- Mizoguchi, T.; Ichimura, K.; Irie, K.; Morris, P.; Giraudat, J.; Matsumoto, K.; Shinozaki, K. Identification of a possible MAP kinase cascade in Arabidopsis thaliana based on pairwise yeast two-hybrid analysis and functional complementation tests of yeast mutants. FEBS Lett 1998, 437, 56–60. [Google Scholar]
- Ichimura, K.; Mizoguchi, T.; Yoshida, R.; Yuasa, T.; Shinozaki, K. Various abiotic stresses rapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6. Plant J 2000, 24, 655–665. [Google Scholar]
- Huang, Y.F.; Li, H.; Gupta, R.; Morris, P.C.; Luan, S.; Kieber, J.J. ATMPK4, an Arabidopsis homolog of mitogen-activated protein kinase, is activated in vitro by AtMEK1 through threonine phosphorylation. Plant Physiol 2000, 122, 1301–1310. [Google Scholar]
- Matsuoka, D.; Nanmori, T.; Sato, K.; Fukami, Y.; Kikkawa, U.; Yasuda, T. Activation of AtMEK1, an Arabidopsis mitogen-activated protein kinase kinase, in vitro and in vivo: Analysis of active mutants expressed in E-coli and generation of the active form in stress response in seedlings. Plant J 2002, 29, 637–647. [Google Scholar]
- Pitzschke, A.; Djamei, A.; Bitton, F.; Hirt, H. A major role of the MEKK1-MKK1/2-MPK4 pathway in ROS signalling. Mol. Plant 2009, 2, 120–137. [Google Scholar]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar]
- Ding, Y.F.; Chen, Z.; Zhu, C. Microarray-based analysis of cadmium-responsive microRNAs in rice (Oryza sativa). J. Exp. Bot 2011, 62, 3563–3573. [Google Scholar]
- Kim, S.H.; Woo, D.H.; Kim, J.M.; Lee, S.Y.; Chung, W.S.; Moon, Y.H. Arabidopsis MKK4 mediates osmotic-stress response via its regulation of MPK3 activity. Biochem. Bioph. Res. Commun 2011, 412, 150–154. [Google Scholar]
- Suarez-Rodriguez, M.C.; Adams-Phillips, L.; Liu, Y.D.; Wang, H.C.; Su, S.H.; Jester, P.J.; Zhang, S.Q.; Bent, A.F.; Krysan, P.J. MEKK1 is required for flg22-induced MPK4 activation in Arabidopsis plants. Plant Physiol 2007, 143, 661–669. [Google Scholar]
- Teige, M.; Scheikl, E.; Eulgem, T.; Doczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol. Cell 2004, 15, 141–152. [Google Scholar]
- Doczi, R.; Brader, G.; Pettko-Szandtner, A.; Rajh, I.; Djamei, A.; Pitzschke, A.; Teige, M.; Hirt, H. The Arabidopsis mitogen-activated protein kinase kinase MKK3 is upstream of group C mitogen-activated protein kinases and participates in pathogen signaling. Plant Cell 2007, 19, 3266–3279. [Google Scholar]
- Takahashi, F.; Yoshida, R.; Ichimura, K.; Mizoguchi, T.; Seo, S.; Yonezawa, M.; Maruyama, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The mitogen-activated protein kinase cascade MKK3-MPK6 is an important part of the jasmonate signal transduction pathway in Arabidopsis. Plant Cell 2007, 19, 805–818. [Google Scholar]
- Takahashi, F.; Mizoguchi, T.; Yoshida, R.; Ichimura, K.; Shinozaki, K. Calmodulin-dependent activation of MAP kinase for ROS homeostasis in Arabidopsis. Mol. Cell 2011, 41, 649–660. [Google Scholar]
- Ortiz-Masia, D.; Perez-Amador, M.A.; Carbonell, J.; Marcote, M.J. Diverse stress signals activate the C1 subgroup MAP kinases of Arabidopsis. FEBS Lett 2007, 581, 1834–1840. [Google Scholar]
- Popescu, S.C.; Popescu, G.V.; Bachan, S.; Zhang, Z.M.; Gerstein, M.; Snyder, M.; Dinesh-Kumar, S.P. MAPK target networks in Arabidopsis thaliana revealed using functional protein microarrays. Gene Dev 2009, 23, 80–92. [Google Scholar]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci 2000, 5, 199–206. [Google Scholar]
- Gadjev, I.; Vanderauwera, S.; Gechev, T.S.; Laloi, C.; Minkov, I.N.; Shulaev, V.; Apel, K.; Inze, D.; Mittler, R.; van Breusegem, F. Transcriptomic footprints disclose specificity of reactive oxygen species signaling in Arabidopsis. Plant Physiol 2006, 141, 436–445. [Google Scholar]
- Zhou, X.; Jiang, Y.J.; Yu, D.Q. WRKY22 transcription factor mediates dark-induced leaf senescence in Arabidopsis. Mol. Cells 2011, 31, 303–313. [Google Scholar]
- Andreasson, E.; Jenkins, T.; Brodersen, P.; Thorgrimsen, S.; Petersen, N.H.T.; Zhu, S.; Qiu, J.L.; Micheelsen, P.; Rocher, A.; Petersen, M.; Newman, M.A.; Nielsen, H.B.; Hirt, H.; Somssich, I.; Mattsson, O.; Mundy, J. The MAP kinase substrate MKS1 is a regulator of plant defense responses. EMBO J 2005, 24, 2579–2589. [Google Scholar]
- Zheng, Z.Y.; Abu Qamar, S.; Chen, Z.X.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J 2006, 48, 592–605. [Google Scholar]
- Li, S.J.; Fu, Q.T.; Huang, W.D.; Yu, D.Q. Functional analysis of an Arabidopsis transcription factor WRKY25 in heat stress. Plant Cell Rep 2009, 28, 683–693. [Google Scholar]
- Jiang, Y.Q.; Deyholos, M.K. Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses. Plant Mol. Biol 2009, 69, 91–105. [Google Scholar]
- Vanderauwera, S.; Zimmermann, P.; Rombauts, S.; Vandenabeele, S.; Langebartels, C.; Gruissem, W.; Inze, D.; van Breusegem, F. Genome-wide analysis of hydrogen peroxide-regulated gene expression in Arabidopsis reveals a high light-induced transcriptional cluster involved in anthocyanin biosynthesis. Plant Physiol 2005, 139, 806–821. [Google Scholar]
- Desikan, R.; Mackerness, S.A.H.; Hancock, J.T.; Neill, S.J. Regulation of the Arabidopsis transcriptome by oxidative stress. Plant Physiol 2001, 127, 159–172. [Google Scholar]
- Cheong, Y.H.; Chang, H.S.; Gupta, R.; Wang, X.; Zhu, T.; Luan, S. Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in Arabidopsis. Plant Physiol 2002, 129, 661–677. [Google Scholar]
- Davletova, S.; Schlauch, K.; Coutu, J.; Mittler, R. The zinc-finger protein Zat12 plays a central role in reactive oxygen and abiotic stress signaling in Arabidopsis. Plant Physiol 2005, 139, 847–856. [Google Scholar]
- Li, S.J.; Fu, Q.T.; Chen, L.G.; Huang, W.D.; Yu, D.Q. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 coordinate induction of plant thermotolerance. Planta 2011, 233, 1237–1252. [Google Scholar]
- Rizhsky, L.; Davletova, S.; Liang, H.J.; Mittler, R. The zinc finger protein Zat12 is required for cytosolic ascorbate peroxidase 1 expression during oxidative stress in Arabidopsis. J. Biol. Chem 2004, 279, 11736–11743. [Google Scholar]
- Davletova, S.; Rizhsky, L.; Liang, H.J.; Zhong, S.Q.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar]
- Vogel, J.T.; Zarka, D.G.; Van Buskirk, H.A.; Fowler, S.G.; Thomashow, M.F. Roles of the CBF2 and ZAT12 transcription factors in configuring the low temperature transcriptome of Arabidopsis. Plant J 2005, 41, 195–211. [Google Scholar]
- Xing, Y.; Jia, W.S.; Zhang, J.H. AtMEK1 mediates stress-induced gene expression of CAT1 catalase by triggering H2O2 production in Arabidopsis. J. Exp. Bot 2007, 58, 2969–2981. [Google Scholar]
- Xing, Y.; Jia, W.S.; Zhangl, J.H. AtMKK1 mediates ABA-induced CAT1 expression and H2O2 production via AtMPK6-coupled signaling in Arabidopsis. Plant J 2008, 54, 440–451. [Google Scholar]
- Kendrick, M.D.; Chang, C. Ethylene signaling: New levels of complexity and regulation. Curr. Opin. Plant Biol 2008, 11, 479–485. [Google Scholar]
- Wasternack, C.; Kombrink, E. Jasmonates: Structural requirements for lipid-derived signals active in plant stress responses and development. ACS Chem. Biol 2010, 5, 63–77. [Google Scholar]
- Liu, Y.D.; Zhang, S.Q. Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in Arabidopsis. Plant Cell 2004, 16, 3386–3399. [Google Scholar]
- Xu, J.; Li, Y.; Wang, Y.; Liu, H.X.; Lei, L.; Yang, H.L.; Liu, G.Q.; Ren, D.T. Activation of MAPK kinase 9 induces ethylene and camalexin biosynthesis and enhances sensitivity to salt stress in Arabidopsis. J. Biol. Chem 2008, 283, 26996–27006. [Google Scholar]
- Shan, X.Y.; Yan, J.B.; Xie, D.X. Comparison of phytohormone signaling mechanisms. Curr. Opin. Plant Biol 2012, 15, 84–91. [Google Scholar]
- Kieber, J.J.; Rothenberg, M.; Roman, G.; Feldmann, K.A.; Ecker, J.R. Ctr1, a negative regulator of the ethylene response pathway in Arabidopsis, Encodes a member of the raf family of protein-kinases. Cell 1993, 72, 427–441. [Google Scholar]
- Novikova, G.V.; Moshkov, I.E.; Smith, A.R.; Hall, M.A. The effect of ethylene on MAPKinase-like activity in Arabidopsis thaliana. FEBS Lett 2000, 474, 29–32. [Google Scholar]
- Ouaked, F.; Rozhon, W.; Lecourieux, D.; Hirt, H. A MAPK pathway mediates ethylene signaling in plants. EMBO J 2003, 22, 1282–1288. [Google Scholar]
- Yoo, S.D.; Cho, Y.H.; Tena, G.; Xiong, Y.; Sheen, J. Dual control of nuclear EIN3 by bifurcate MAPK cascades in C2H4 signalling. Nature 2008, 451, 789–796. [Google Scholar]
- Bethke, G.; Unthan, T.; Uhrig, J.F.; Poschl, Y.; Gust, A.A.; Scheel, D.; Lee, J. Flg22 regulates the release of an ethylene response factor substrate from MAP kinase 6 in Arabidopsis thaliana via ethylene signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 8067–8072. [Google Scholar]
- Remans, T.; Opdenakker, K.; Smeets, K.; Mathijsen, D.; Vangronsveld, J.; Cuypers, A. Metal-specific and NADPH oxidase dependent changes in lipoxygenase and NADPH oxidase gene expression in Arabidopsis thaliana exposed to cadmium or excess copper. Funct. Plant Biol 2010, 37, 532–544. [Google Scholar]
- Opdenakker, K. Centre for Environmental Sciences, Hasselt University, Diepenbeek, Belgium. MPK6 regulates transcription of lipoxygenases in Arabidopsis thaliana. Unpublished work. 2012. [Google Scholar]
- Bell, E.; Creelman, R.A.; Mullet, J.E. A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 8675–8679. [Google Scholar]
- Leon, J.; Rojo, E.; Sanchez-Serrano, J.J. Wound signalling in plants. J. Exp. Bot 2001, 52, 1–9. [Google Scholar]
- Petersen, M.; Brodersen, P.; Naested, H.; Andreasson, E.; Lindhart, U.; Johansen, B.; Nielsen, H.B.; Lacy, M.; Austin, M.J.; Parker, J.E.; Sharma, S.B.; Klessig, D.F.; Martienssen, R.; Mattsson, O.; Jensen, A.B.; Mundy, J. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell 2000, 103, 1111–1120. [Google Scholar]
- Van Bentem, S.D.; Anrather, D.; Dohnal, I.; Roitinger, E.; Csaszar, E.; Joore, J.; Buijnink, J.; Carreri, A.; Forzani, C.; Lorkovic, Z.J.; Barta, A.; Lecourieux, D.; Verhounig, A.; Jonak, C.; Hirt, H. Site-specific phosphorylation profiling of Arabidopsis proteins by mass spectrometry and peptide chip analysis. J. Proteome Res 2008, 7, 2458–2470. [Google Scholar]


{kind=link}
| Classification of MAPKinases | |||||
|---|---|---|---|---|---|
| MAPKinase | Amino acids phosphorylated by the kinase | Phosphorylated amino acid motif | Group | Examples | Response to/in |
| MAPKKK | Serine/threonine | MEKK1-type | A | MEKK1, MEKK2, MEKK3, MEKK4 | Redox control–oxidative stress; Abiotic stresses: drought, salt, mechanical; Biotic stress: bacterial pathogens; Hormones: salicylic acid. |
| ANP1, ANP2, ANP3 | Redox control–oxidative stress; Biotic stress: bacterial pathogens; Hormones: auxin; Cytokinesis. | ||||
| Raf-like | B | EDR1, CTR1 | Hormones: ethylene; Disease resistance signling. | ||
| C | ATN1, MRK1 | Unknown. | |||
| MAPKK | Threonine/tyrosine | S/T-XXXXX-S/T | A | MKK1, MKK2, MKK6 | Redox control–oxidative stress; Abiotic stresses: cold, salt, low humidity, mechanical; Biotic stresses: bacterial pathogens; Hormones: salicylic acid; Cell division. |
| B | MKK3 | Oxidative stress; Abiotic stresses: mechanical; Biotic stresses: bacterial pathogens; Hormones: jasmonic acid. | |||
| MAPKKK | Threonine/tyrosine | S/T-XXXXX-S/T | C | MKK4, MKK5 | Redox control–oxidative stress; Abiotic stresses: salt; Biotic stress: bacterial pathogens; Hormones. |
| D | MKK7, MKK8, MKK9, MKK10 | Oxidative stress; Biotic stress: bacterial pathogens. | |||
| MAPK | Serine/threonine/tyrosine | TEY | A | MPK3, MPK6, MPK10 | Redox control–oxidative stress; Abiotic stresses: salt, cold; Biotic stress: bacterial pathogens; Hormones: jasmonic acid. |
| B | MPK4, MPK5, MPK11, MPK12, MPK13 | Redox control–oxidative stresses; Abiotic stresses: salt, cold, low humidity, mechanical; Hormones: salicylic acid; Cell division. | |||
| C | MPK1, MPK2, MPK7, MPK14 | Oxidative stress; Abiotic stresses: mechanical; Biotic stresses: bacterial pathogens; Hormones: jasmonic acid, abscisic acid; Circadian-rhythm-regulated. | |||
| TDY | D | MPK8, MPK9, MPK15/16/17/18/19/20 | Oxidative stress; Abiotic stresses: mechanical; Biotic: blast fungus; Hormones: jasmonic acid. | ||
| Metal-Induced MAPKinases | |||||||
|---|---|---|---|---|---|---|---|
| Plant | Component of MAPK cascade | Metal | Concentration | Exposure Time | Observations | Reference | |
| Arabidopsis thaliana | MAPKKK | MEKK1 | Cd | 500 μM CdCl2 | 1–3 h | ↑ mRNA levels | [66] |
| MAPK | MPK3/MPK6 | Cd | 1 μM CdCl2 | 10 min | ↑ activity | [60] | |
| Cu/Cd | 2 μM CuSO4/5 μM CdSO4 | 2–24 h | ↑ mRNA levels | [21] | |||
| Brassica juncea | MAPK | 46 kDa MAPK | As(III) | 50 μM As(III) | 15 min–1 h | ↑ activity | [67] |
| Medicago sativa | MAPKK | SIMKK | Cu | 100 μM CuCl2 | 30 min | induces SAMK and SIMK | [62] |
| MAPK | SAMK | Cu/Cd | 100 μM CuCl2/CdCl2 | 10 min–1 h (Cu)/30 min–3 h (Cd) | ↑ activity | [62] | |
| SIMK | Cu/Cd | 100 μM CuCl2 /CdCl2 | 5 min–6 h (Cu)/10 min–6 h (Cd) | ↑ activity | [62] | ||
| MMK2 | Cu/Cd | 100 μM CuCl2/CdCl2 | 10 min–1 h (Cu)/10 min–3 h (Cd) | ↑ activity | [62] | ||
| MMK3 | Cu/Cd | 100 μM CuCl2/CdCl2 | 10 min–1 h (Cu)/10 min–1 h (Cd) | ↑ activity | [62] | ||
| Oryza sativa | MAPKK | OsMKK4 | As | 50 μM As(III) | 3–9 h | ↑ mRNA levels | [61] |
| MAPK | OsMSRMK2 | Cu/Cd/Hg | 100 μM CuSO4/CdCl2/HgClO3 | 30 min | ↑ mRNA levels | [68] | |
| OsMSRMK3 | Cu/Cd/Hg | 100 μM CuSO4/CdCl2/HgClO3 | 15 min–2 h | ↑ mRNA levels | [69] | ||
| OsWJUMK | Cu/Cd/Hg | 100 μM CuSO4/CdCl2/HgClO3 | 15 min–2 h | ↑ mRNA levels | [69] | ||
| OsMPK2 | Cd | 400 μM CdCl2 | 3–12 h | ↑ mRNA levels | [70] | ||
| Cu | 100 μM CuCl2 | 3–12 h | ↑ mRNA levels | [71,72] | |||
| OsMPK3 | Cu/Cd | 50 μM CuCl2/100 μM CdCl2 | 1 h | ↑ activity | [73] | ||
| As | 50 μM As(III) | 30 min–9 h | ↑ mRNA and activity levels | [61] | |||
| OsMPK4 | As | 50 μM As(III) | 30 min–9 h | ↑ mRNA and activity levels | [61] | ||
| OsMPK6 | Cu/Cd | 50 μM CuCl2/100 μM CdCl2 | 1 h | ↑ activity | [73] | ||
| 40 kDa MAPK | Zn | 1 mM ZnCl2 | 15 min–8 h | ↑ activity | [74] | ||
| Pb | 10 mM Pb(NO3)2 | 30 min–8 h | ↑ activity | [75] | |||
| 42 kDa MAPK | Zn | 1 mM ZnCl2 | 15 min–8 h | ↑ activity | [74] | ||
| I | 500 μM FeSO4 | 15–30 min | ↑ activity | [76] | |||
| Pb | 10 mM Pb(NO3)2 | 15 min–8 h | ↑ activity | [75] | |||
| Zea mays | MAPK | ZmMPK3 | Cd | 500 CdCl2 | 30 min–1 h | ↑ mRNA levels | [77] |
| ZmMPK5 | Cr(VI) | 250 μM Cr(VI) | 30 min | ↑ activity | [78] | ||
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Opdenakker, K.; Remans, T.; Vangronsveld, J.; Cuypers, A. Mitogen-Activated Protein (MAP) Kinases in Plant Metal Stress: Regulation and Responses in Comparison to Other Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2012, 13, 7828-7853. https://doi.org/10.3390/ijms13067828
Opdenakker K, Remans T, Vangronsveld J, Cuypers A. Mitogen-Activated Protein (MAP) Kinases in Plant Metal Stress: Regulation and Responses in Comparison to Other Biotic and Abiotic Stresses. International Journal of Molecular Sciences. 2012; 13(6):7828-7853. https://doi.org/10.3390/ijms13067828
Chicago/Turabian StyleOpdenakker, Kelly, Tony Remans, Jaco Vangronsveld, and Ann Cuypers. 2012. "Mitogen-Activated Protein (MAP) Kinases in Plant Metal Stress: Regulation and Responses in Comparison to Other Biotic and Abiotic Stresses" International Journal of Molecular Sciences 13, no. 6: 7828-7853. https://doi.org/10.3390/ijms13067828