HR4 Gene Is Induced in the Arabidopsis-Trichoderma atroviride Beneficial Interaction


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
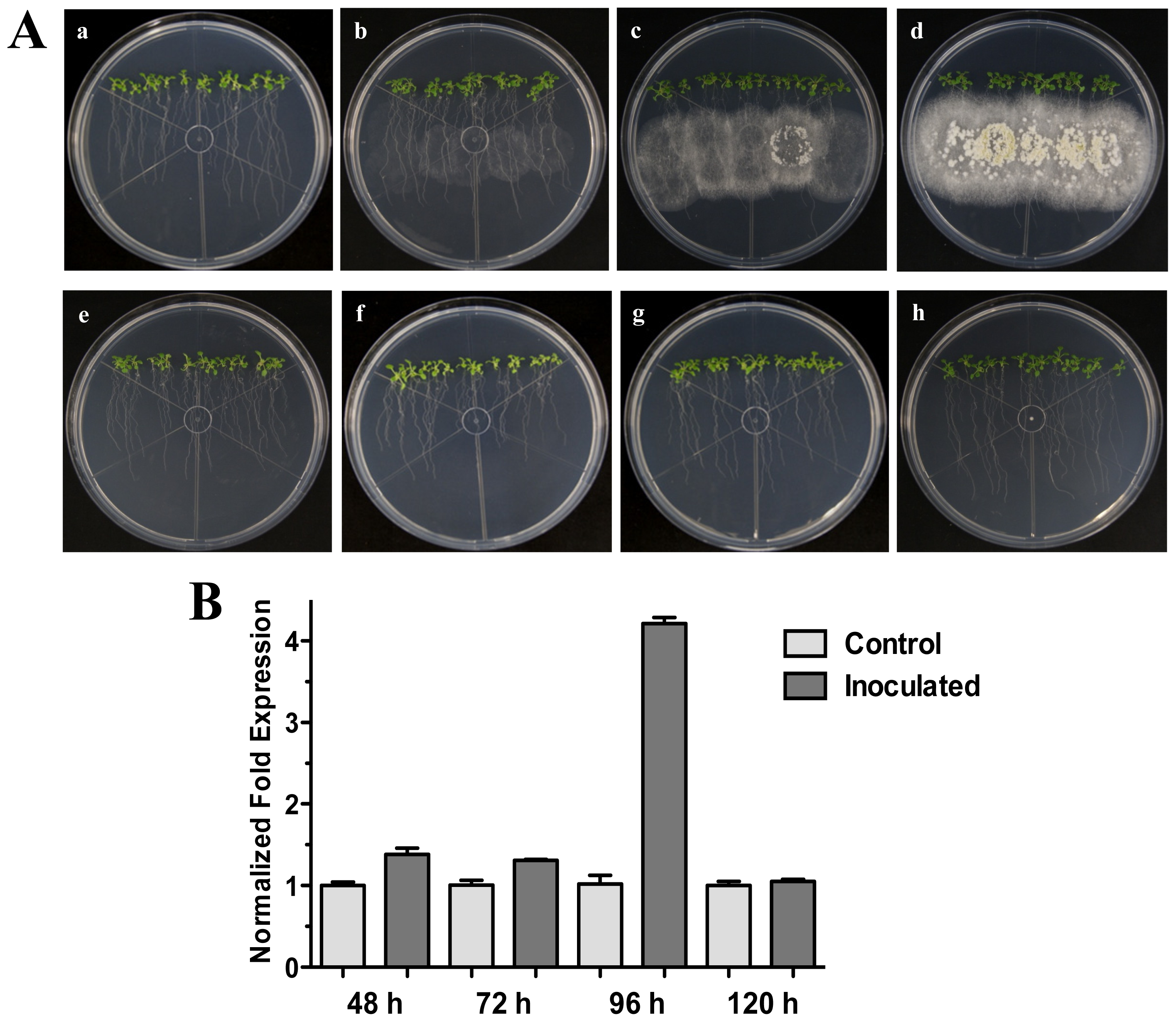
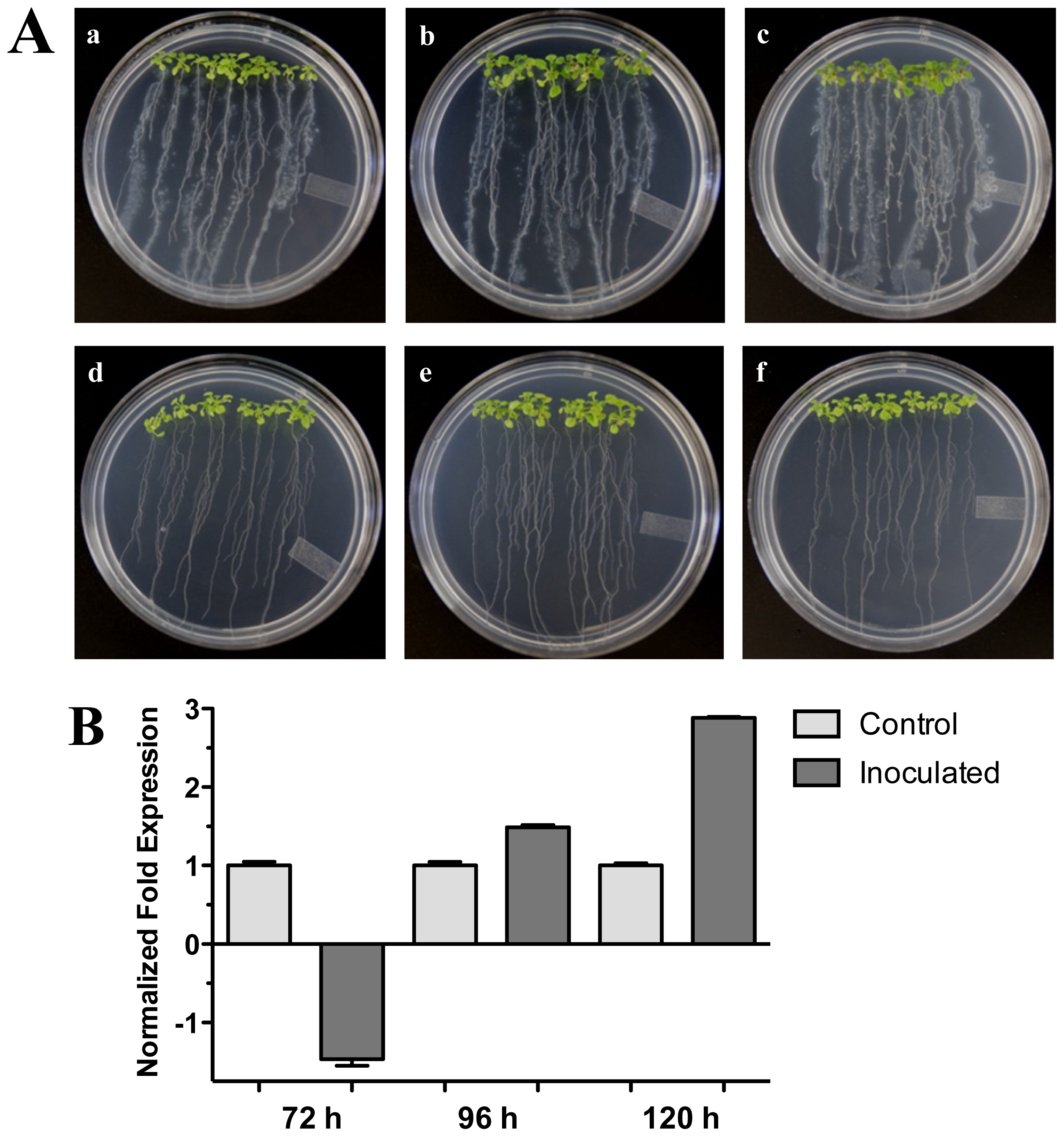
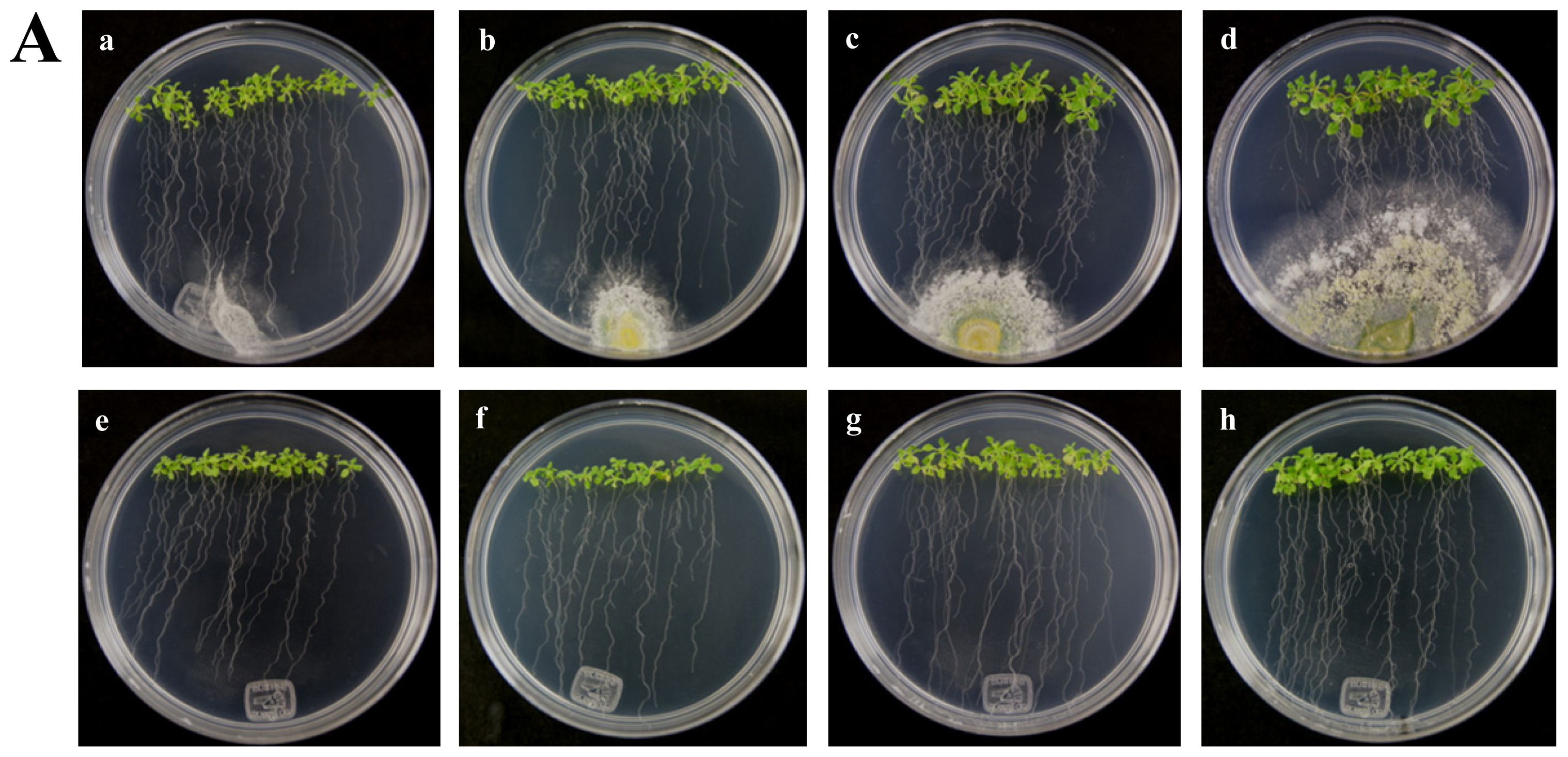
2.1. The HR4 Gene (Ecotype Col-0) Expression in Interaction with Trichoderma
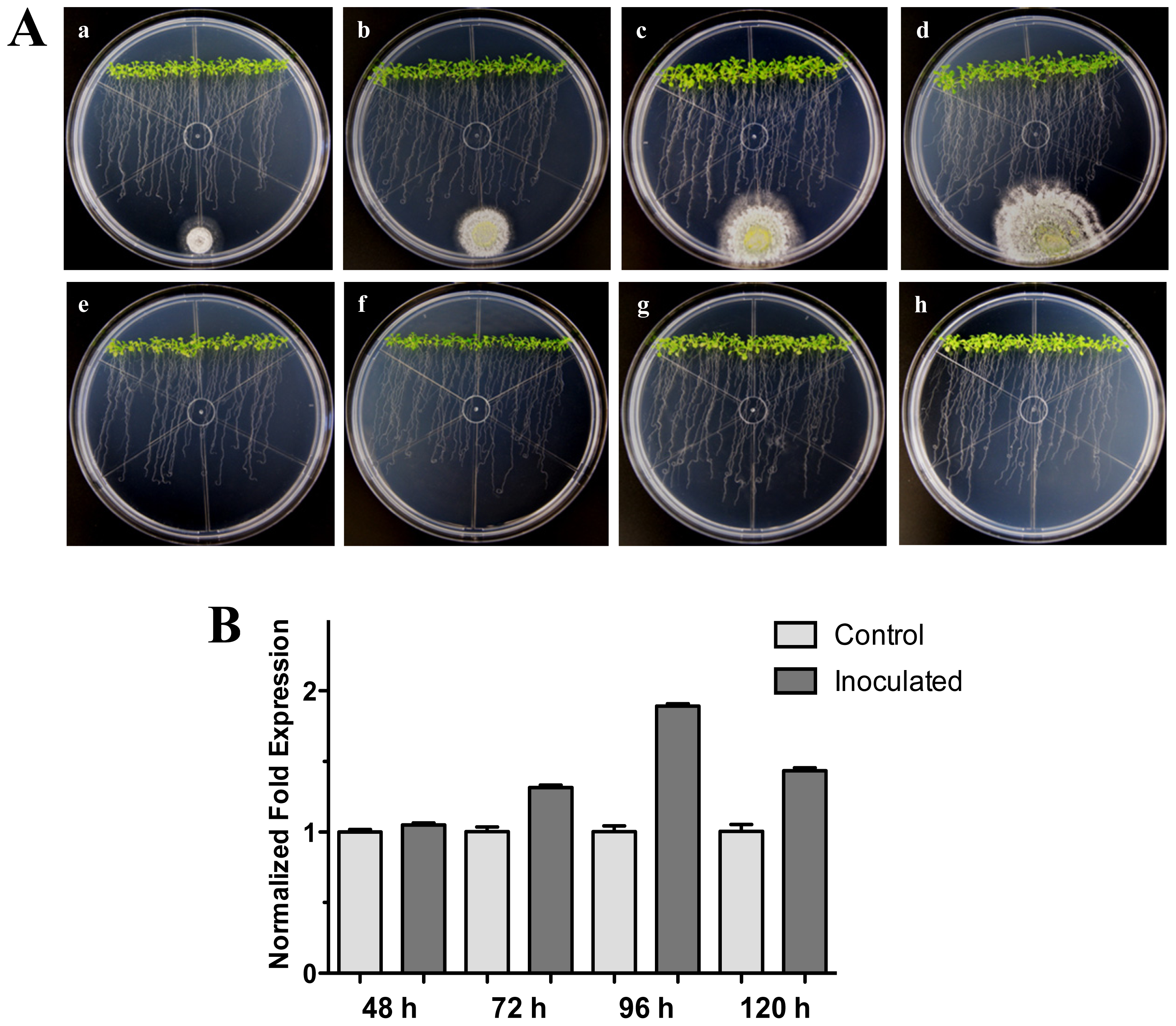
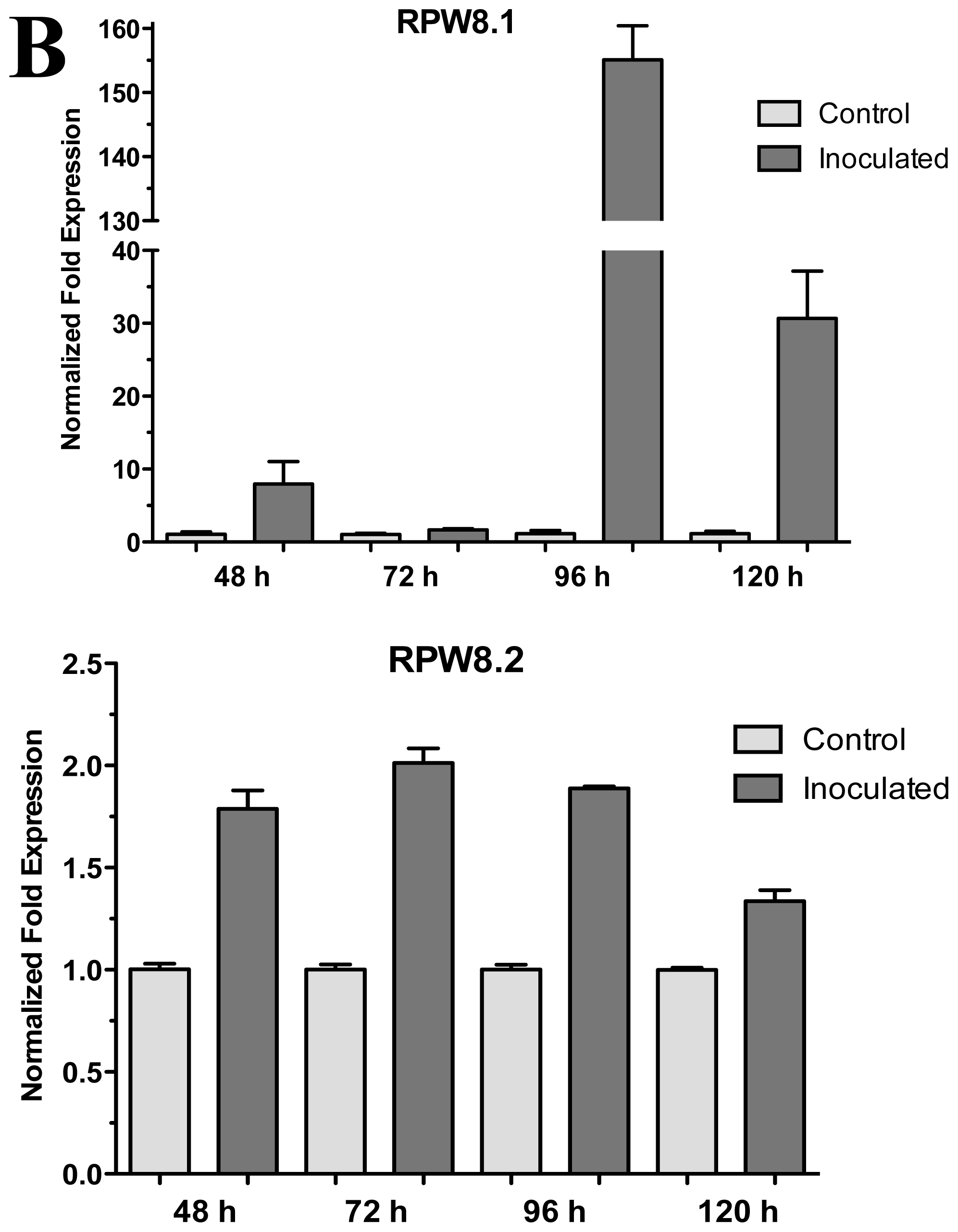
2.2. The RPW8.1 and RPW8.2 Genes (Ecotype Ms-0) Expression in Interaction with Trichoderma atroviride
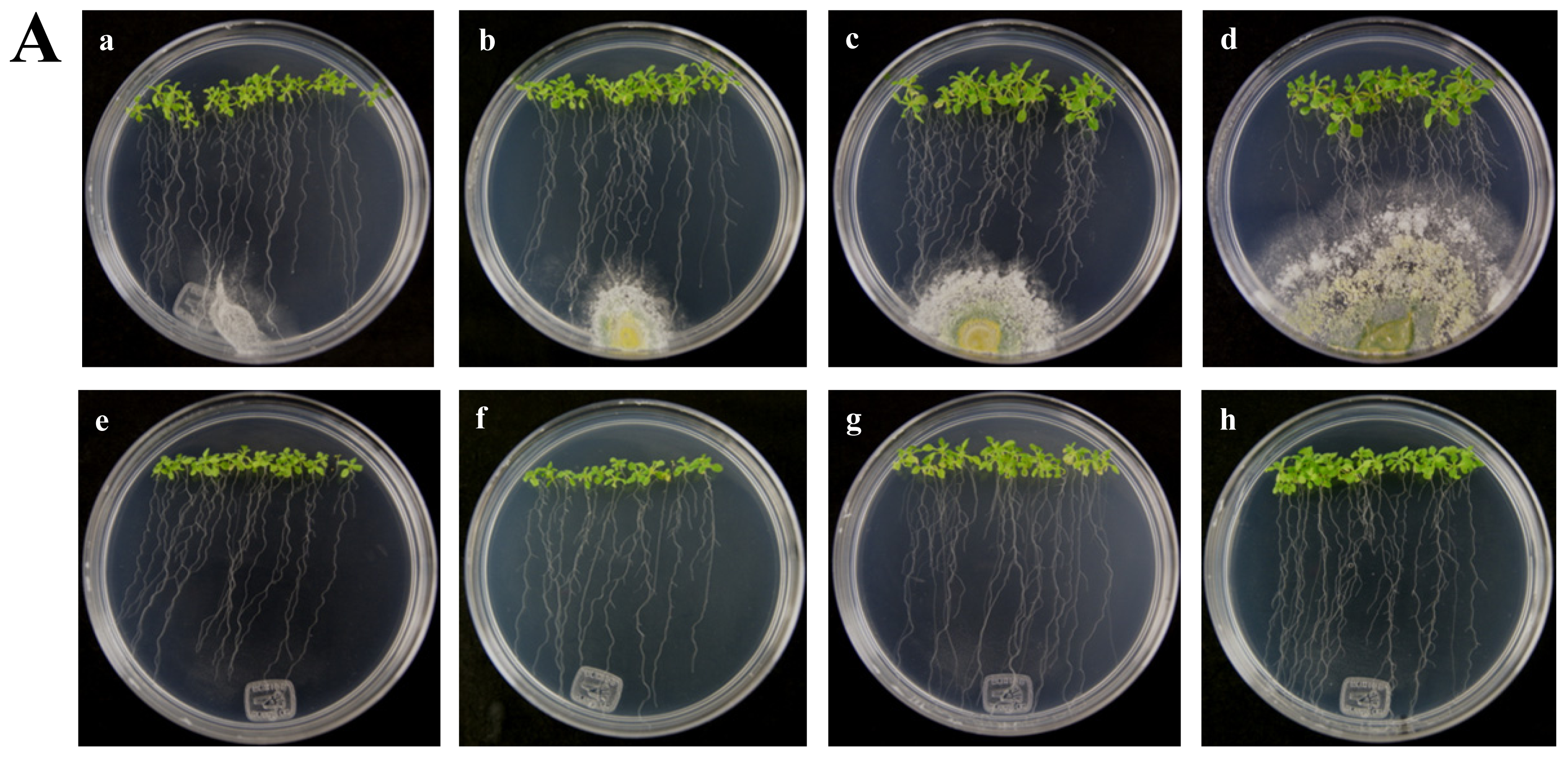
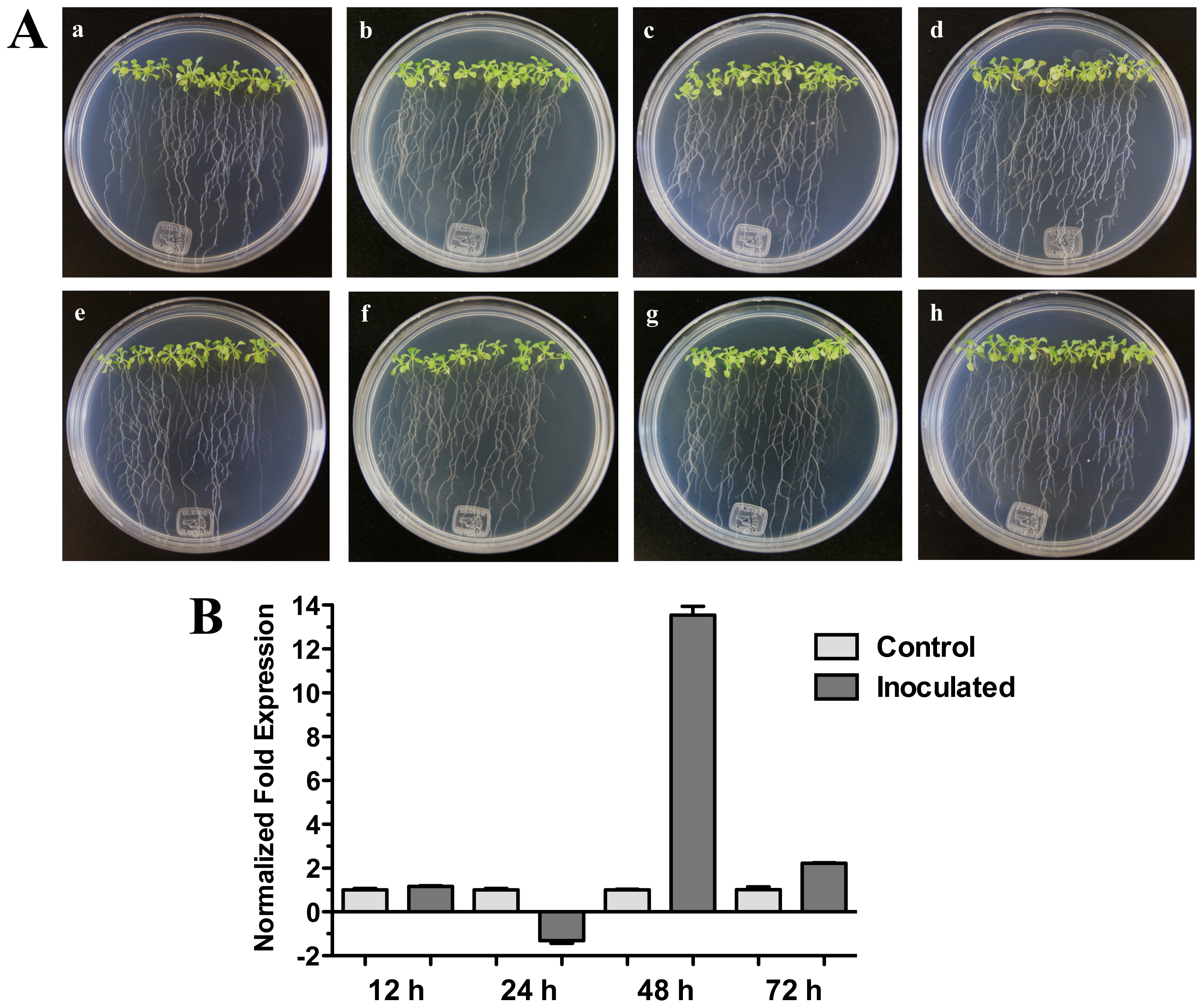
2.3. HR4 Gene Is Induced by Bacterial Interactions
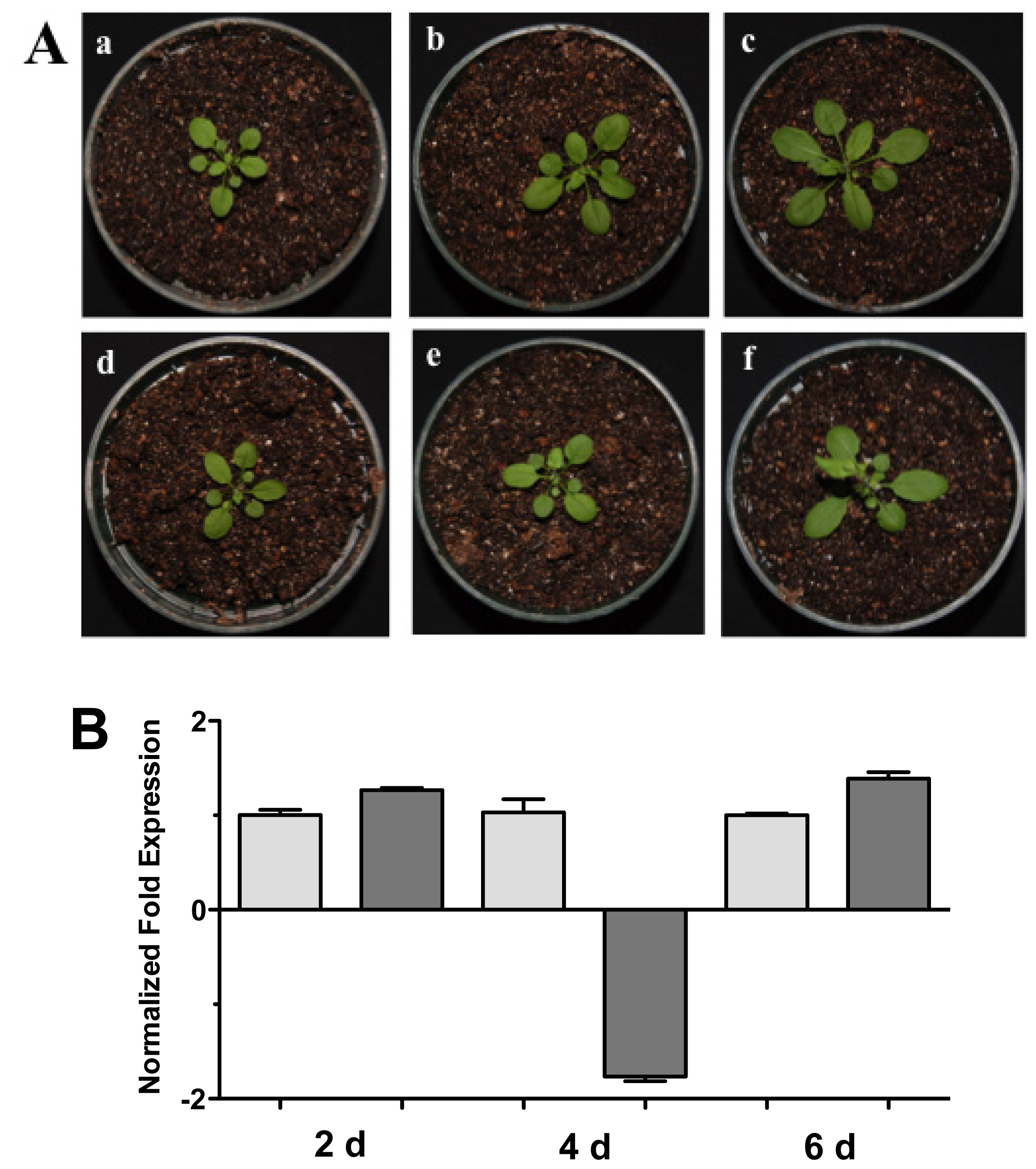
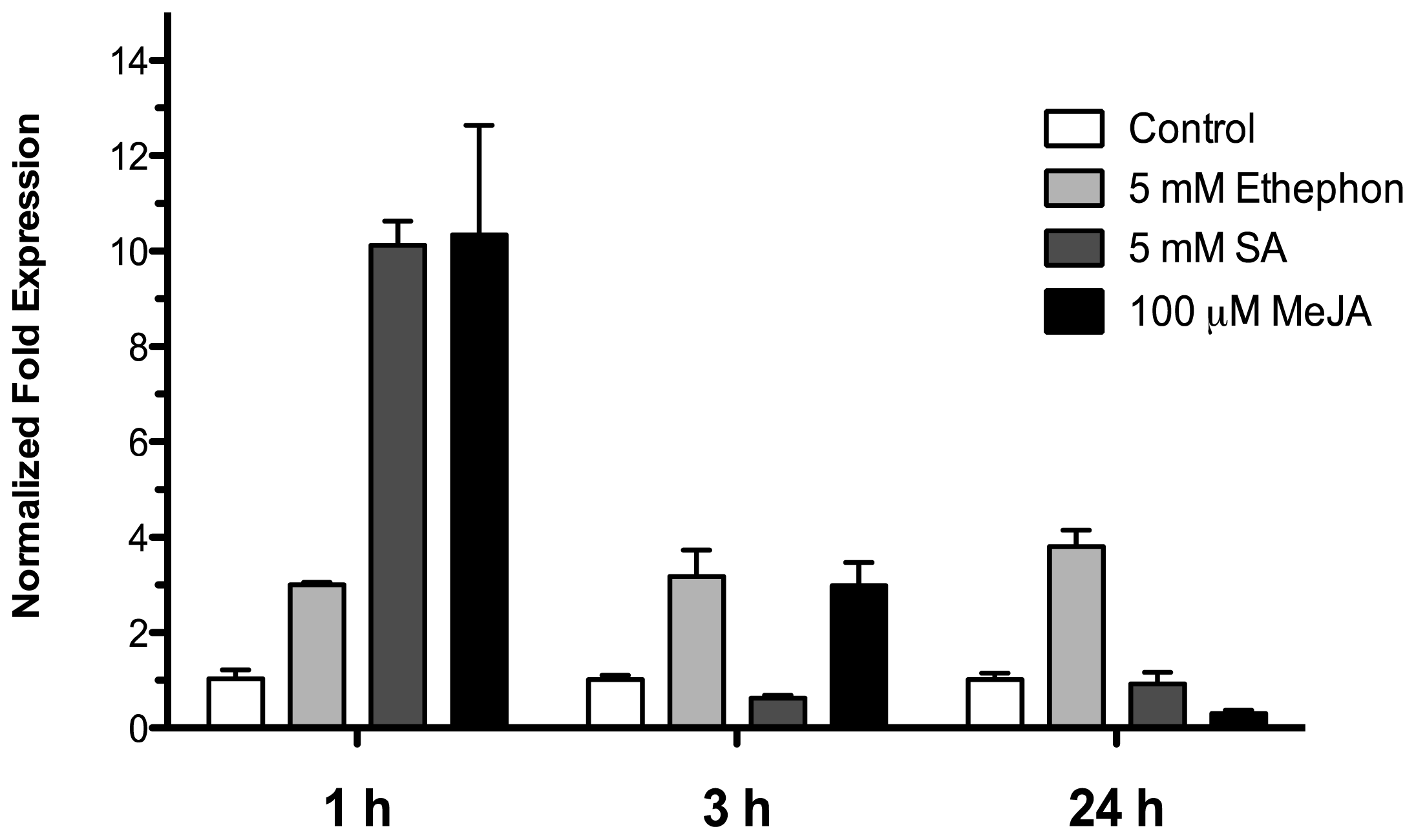
2.4. Effect of Phytohormones on the Expression of HR4 Gene
3. Discussion
4. Experimental Section
4.1. Plant Material and Growth Conditions
4.2. Microorganism Growth Conditions and Seedling Inoculations
4.3. Phytohormone Application
4.4. RNA Extraction and Real-Time qRT-PCR
5. Conclusions
Acknowledgments
References
- Zhao, S.; Qi, X. Signaling in plant disease resistance and symbiosis. J. Integr. Plant Biol 2008, 7, 799–807. [Google Scholar]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar]
- Lotze, M.T.; Zeh, H.J.; Rubartelli, A.; Sparvero, L.J.; Amoscato, A.A.; Washburn, N.R.; Devera, M.E.; Liang, X.; Tör, M.; Billiar, T. The grateful dead: Damage-associated molecular pattern molecules and reduction/oxidation regulate immunity. Immunol. Rev 2007, 220, 60–81. [Google Scholar]
- Tör, M.; Lotze, M.T.; Holton, N. Receptor-mediated signalling in plants: Molecular patterns and programmes. J. Exp. Bot 2009, 60, 3645–3654. [Google Scholar]
- Zipfel, C.; Robatzek, S. Pathogen-associated molecular pattern-triggered immunity: Veni, Vidi…? Plant Physiol 2010, 154, 551–554. [Google Scholar]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet 2010, 11, 539–548. [Google Scholar]
- Zamioudis, C.; Pieterse, C.M.J. Modulation of host immunity by beneficial microbes. Mol. Plant Microbe Interact 2012, 25, 139–150. [Google Scholar]
- Gururani, M.A.; Venkatesh, J.; Upadhyaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol 2012, 78, 51–65. [Google Scholar]
- Bent, A.F.; Mackey, D. Elicitors, Effectors, and R Genes: The new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol 2007, 1, 399–436. [Google Scholar]
- Xiao, S.; Wang, W.; Yang, X. Evolution of resistance genes. Nucleic Acids Mol. Biol 2008, 21, 1–25. [Google Scholar]
- Micali, C.; Göllner, K.; Humphry, M.; Consonni, C.; Panstruga, R. The Powdery Mildew Disease of Arabidopsis: A Paradigm for the Interaction Between Plants and Biotrophic Fungi. In The Arabidopsis Book; American Society of Plant Biology: Rockville, MD, USA, 2008; Volume 6, pp. 1–19. [Google Scholar]
- Xiao, S.; Ellwood, S.; Calis, O.; Patrick, E.; Li, T.X.; Coleman, M.; Turner, J.G. Broad-spectrum mildew resistance in Arabidopsis thaliana mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar]
- Xiao, S.; Emerson, B.; Ratanasut, K.; Patrick, E.; O’Neill, C.; Bancroft, I.; Turner, J.G. Origin and maintenance of a broad-spectrum disease resistance locus in Arabidopsis. Mol. Biol. Evol 2004, 21, 1661–1672. [Google Scholar]
- Orgil, U.; Araki, H.; Tangchaiburana, S.; Berkey, R.; Xiao, S. Intraespecific genetic variations, fitness cost and benefit of RPW8, a disease resistance locus in Arabidopsis thaliana. Genetics 2007, 176, 2317–2333. [Google Scholar]
- Hoyos-Carvajal, L.; Ordua, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control 2009, 51, 409–416. [Google Scholar]
- Moran-Diez, E.; Hermosa, R.; Ambrosino, P.; Cardoza, R.E.; Gutiérrez, S.; Lorito, M.; Monte, E. The ThPG1 endopolygalacturonase is required for the Trichoderma harzianum–plant beneficial interaction. Mol. Plant Microbe Interact 2009, 22, 1021–1031. [Google Scholar]
- Tucci, M.; Ruocco, M.; de Masi, L.; de Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol 2011, 12, 341–354. [Google Scholar]
- Yoshioka, Y.; Ichikawa, H.; Naznin, H.A.; Kogure, A.; Hyakumachi, M. Systemic resistance induced in Arabidopsis thaliana by Trichoderma asperellum SKT-1, a microbial pesticide of seedborne diseases of rice. Pest Manag. Sci 2012, 68, 60–66. [Google Scholar]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.I.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol 2009, 149, 1579–1592. [Google Scholar]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar]
- Viterbo, A.; Wiest, A.; Brotman, Y.; Chet, I.; Kenerley, C. The 18mer peptaibols from Trichoderma virens elicit plant defence responses. Mol. Plant Pathol 2007, 8, 737–746. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol 2004, 2, 43–56. [Google Scholar]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational research on Trichoderma: From ‘omics to the field. Annu. Rev. Phytopathol 2010, 48, 395–417. [Google Scholar]
- Delgado-Sánchez, P.; Ortega-Amaro, M.A.; Rodríguez-Hernández, A.A.; Jiménez-Bremont, J.F.; Flores, J. Further evidence from the effect of fungi on breaking Opuntia seed dormancy. Plant Signal. Behav 2010, 5, 1229–1230. [Google Scholar]
- Delgado-Sánchez, P.; Ortega-Amaro, M.A.; Jiménez-Bremont, J.F.; Flores, J. Are fungi important for breaking seed dormancy in desert species? Experimental evidence in Opuntia streptacantha (Cactaceae). Plant Biol 2011, 13, 154–159. [Google Scholar]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar]
- Harman, G.E. Myths and dogmas of biocontrol: Changes in perceptions derived from research on Trichoderma harzianum T-22. Plant Dis 2000, 84, 377–393. [Google Scholar]
- Bae, H.; Sicher, R.C.; Kim, M.S.; Kim, S.H.; Strem, M.D.; Melnick, R.L.; Bailey, B.A. The beneficial endophyte Trichoderma hamatum isolate DIS 219b promotes growth and delays the onset of the drought response in Theobroma cacao. J. Exp. Bot 2009, 60, 3279–3295. [Google Scholar]
- Yildirim, E.; Taylor, A.G.; Spittler, T.D. Ameliorative effects of biological treatments on growth of squash plants under salt stress. Sci. Hortic. (Amst) 2006, 111, 1–6. [Google Scholar]
- Alfano, G.; Ivey, M.L.L.; Cakir, C.; Bos, J.I.B.; Miller, S.A.; Madden, L.V.; Kamoun, S.; Hoitink, H.A.J. Systemic modulation of gene expression in tomato by Trichoderma hamatum 382. Phytopathology 2007, 97, 429–437. [Google Scholar]
- Marra, R.; Ambosino, P.; Carbone, V.; Vinale, F.; Woo, S.L.; Ruocco, M.; Ciliento, R.; Lanzuise, S.; Ferraioli, S.; Soriente, I.; et al. Study of the three-way interaction between Trichoderma atroviride, plant and fungal pathogens using a proteome approach. Curr. Genet 2006, 50, 307–321. [Google Scholar]
- Segarra, G.; Casanova, E.; Bellido, D.; Odena, M.A.; Oliveira, E.; Trillas, I. Proteome, salicylic acid, and jasmonic acid changes in cucumber plants inoculated with Trichoderma asperellum strain T34. Proteomics 2007, 7, 3943–3952. [Google Scholar]
- Shoresh, M.; Harman, G.E. The molecular basis of shoot responses of maize seedlings to Trichoderma harzianum T22 inoculation of the root: A proteomic approach. Plant Physiol 2008, 147, 2147–2163. [Google Scholar]
- Moreno, C.A.; Castillo, F.; González, A.; Bernal, D.; Jaimes, Y.; Chaparro, M.; González, C.; Rodriguez, F.; Restrepo, S.; Cotes, A.M. Biological and molecular characterization of the response of tomato plants treated with Trichoderma koningiopsis. Physiol. Mol. Plant Pathol 2009, 74, 111–120. [Google Scholar]
- Morán-Diez, E.; Rubio, B.; Domínguez, S.; Hermosa, R.; Monte, E.; Nicolás, C. Transcriptional response of Arabidopsis thaliana after 24 h incubation with the biocontrol fungus Trichoderma harzianum. Plant Physiol 2012, 169, 614–620. [Google Scholar]
- Brotman, Y.; Lisec, J.; Méret, M.; Chet, I.; Willmitzer, L.; Viterbo, A. Transcript and metabolite analysis of the Trichoderma-induced systemic resistance response to Pseudomonas syringae in Arabidopsis thaliana. Microbiology 2012, 158, 139–146. [Google Scholar]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol 2010, 48, 21–43. [Google Scholar]
- Sáenz-Mata, J.; Jiménez-Bremont, J.F. Institute Potosino of Scientific and Technological Research: San Luis Potosí, SLP, México; Unpublished work; 2009. [Google Scholar]
- Mulema, J.M.K.; Denby, K.J. Spatial and temporal transcriptomic analysis of the Arabidopsis thaliana-Botrytis cinerea interaction. Mol. Biol. Rep 2012, 39, 4039–4049. [Google Scholar]
- Xiao, S.Y.; Brown, S.; Patrick, E.; Brealey, C.; Turner, J.G. Enhanced transcription of the Arabidopsis disease resistance genes RPW8.1 and RPW8.2 occurs via a salicylic acid-dependent amplification circuit and is required for hypersensitive cell death. Plant Cell 2003, 15, 33–45. [Google Scholar]
- Wang, Y.; Ohara, Y.; Nakayashiki, H.; Tosa, Y.; Mayama, S. Microarray analysis of the gene expression profile induced by the endophytic plant growth-promoting rhizobacteria, Pseudomonas fluorescens FPT9601-T5 in Arabidopsis. Mol. Plant Microbe Interact 2005, 18, 385–396. [Google Scholar]
- Verhage, A.; van Wees, S.C.M.; Pieterse, C.M.J. Plant immunity: It’s the hormones talking, but what do they say? Plant Physiol 2010, 154, 536–540. [Google Scholar]
- Pieterse, C.M.J.; Leon-Reyes, A.; van der Ent, S.; van Wees, S.C.M. Networking by small-molecules hormones in plant immunity. Nat. Chem. Biol 2009, 5, 308–316. [Google Scholar]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol 2009, 69, 473–488. [Google Scholar]
- Salas-Marina, M.A.; Silva-Flores, M.A.; Uresti-Rivera, E.E.; Castro-Longoria, E.; Herrera-Estrella, A.; Casas-Flores, S. Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acid/ethylene and salicylic acid pathways. Eur. J. Plant. Pathol 2011, 131, 15–26. [Google Scholar]
- Xiao, S.; Calis, O.; Patrick, E.; Zhang, G.; Charoenwattana, P.; Muskett, P.; Parker, J.E.; Turner, J.G. The atypical resistance gene, RPW8, recruits components of basal defence for powdery mildew resistance in Arabidopsis. Plant J 2005, 42, 95–110. [Google Scholar]
- Feys, B.J.; Moisan, L.J.; Newman, M.A.; Parker, J.E. Direct interaction between the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4. EMBO J 2001, 20, 5400–5411. [Google Scholar]
- Blanco, F.; Garreton, V.; Frey, N.; Dominguez, C.; Perez-Acle, T.; van der Straeten, D.; Jordana, X.; Holuigue, L. Identification of NPR1-dependent and independent genes early induced by salicylic acid treatment in Arabidopsis. Plant Mol. Biol 2005, 59, 927–944. [Google Scholar]
- Galon, Y.; Nave, R.; Boyce, J.M.; Nachmias, D.; Knight, M.R.; Fromm, H. Calmodulin-binding transcription activator (CAMTA) 3 mediates biotic defense responses in Arabidopsis. FEBS Lett 2008, 582, 943–948. [Google Scholar]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescein. J. Lab. Clin. Med 1954, 44, 301–307. [Google Scholar]







© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sáenz-Mata, J.; Jiménez-Bremont, J.F. HR4 Gene Is Induced in the Arabidopsis-Trichoderma atroviride Beneficial Interaction. Int. J. Mol. Sci. 2012, 13, 9110-9128. https://doi.org/10.3390/ijms13079110
Sáenz-Mata J, Jiménez-Bremont JF. HR4 Gene Is Induced in the Arabidopsis-Trichoderma atroviride Beneficial Interaction. International Journal of Molecular Sciences. 2012; 13(7):9110-9128. https://doi.org/10.3390/ijms13079110
Chicago/Turabian StyleSáenz-Mata, Jorge, and Juan Francisco Jiménez-Bremont. 2012. "HR4 Gene Is Induced in the Arabidopsis-Trichoderma atroviride Beneficial Interaction" International Journal of Molecular Sciences 13, no. 7: 9110-9128. https://doi.org/10.3390/ijms13079110
APA StyleSáenz-Mata, J., & Jiménez-Bremont, J. F. (2012). HR4 Gene Is Induced in the Arabidopsis-Trichoderma atroviride Beneficial Interaction. International Journal of Molecular Sciences, 13(7), 9110-9128. https://doi.org/10.3390/ijms13079110