Dual Targeting and Retrograde Translocation: Regulators of Plant Nuclear Gene Expression Can Be Sequestered by Plastids

Abstract
:1. Introduction: Plastids as Sensors and Efficient Communicators of Environmental Conditions
2. The Concept of Compartment-Specific Protein Targeting Requires Revision
3. A Growing Number of Genes Encode Proteins Targeted to More Than One DNA Containing Compartment
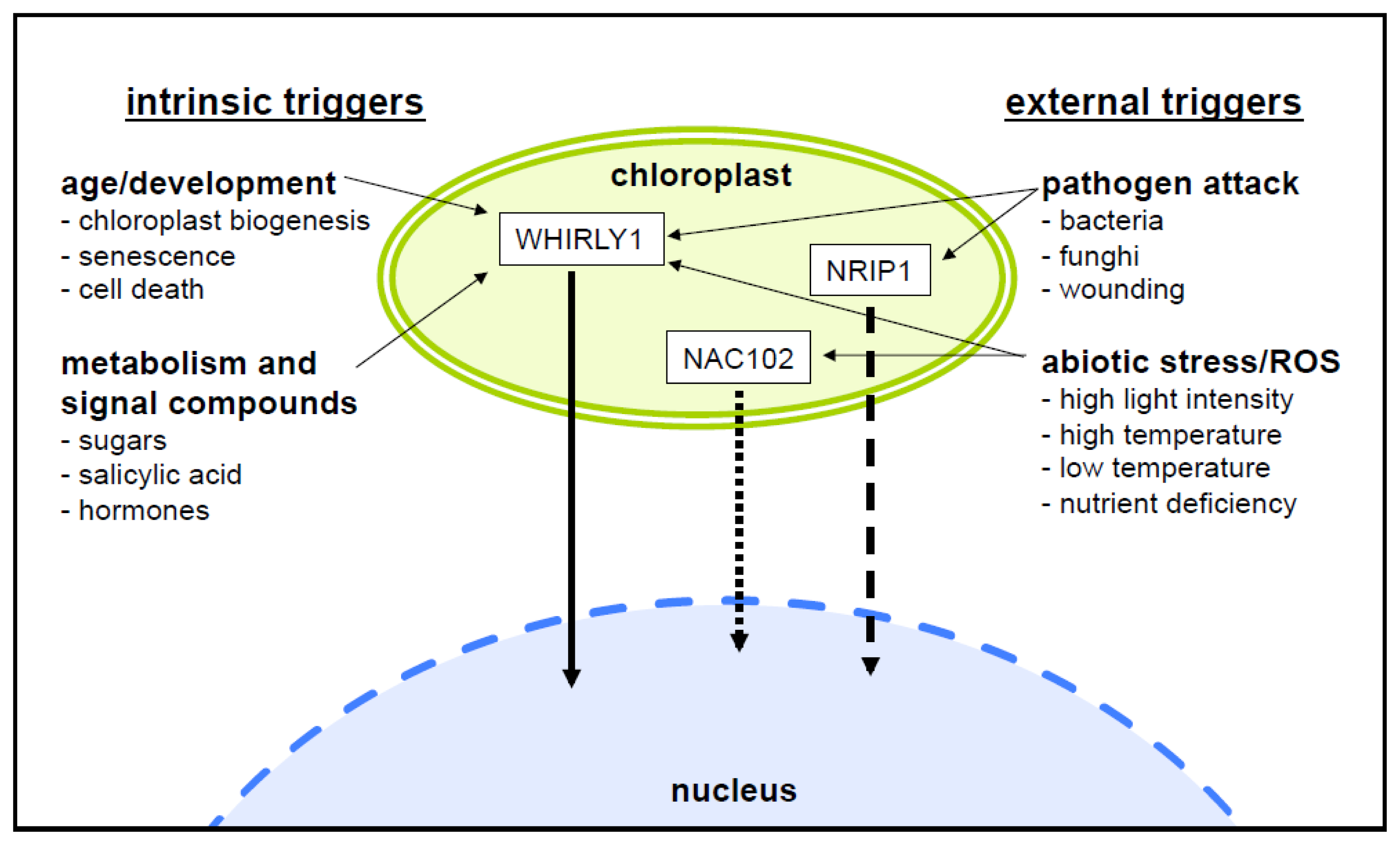
4. What is the Reason for Sequestration of Nuclear Proteins in Plastids?
5. Most Plastid/Nucleus Targeted Proteins are Involved in Gene Expression
6. What is the Evidence for Protein Release from Plastids?
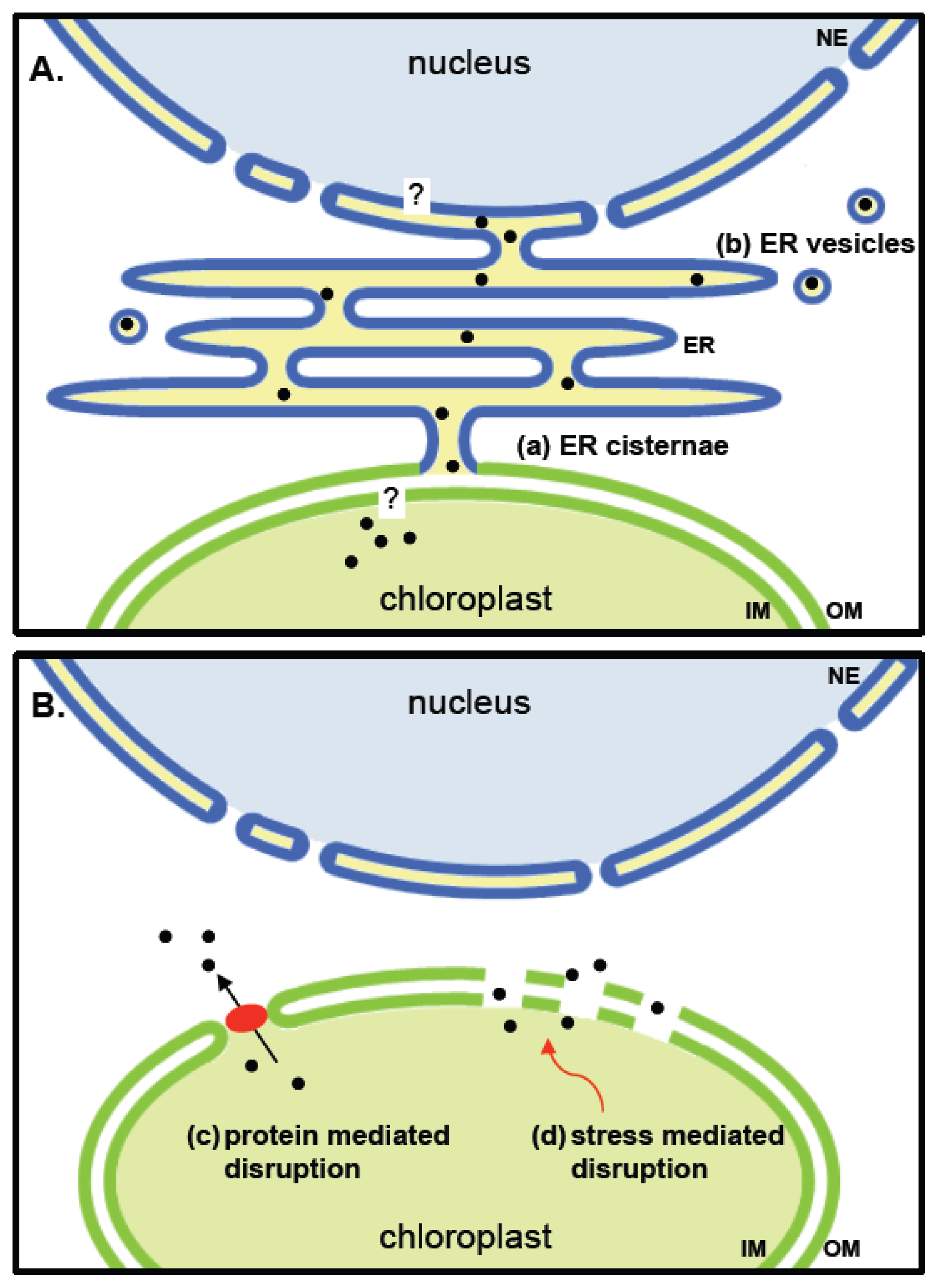
7. Putative Release Pathways for Proteins Sequestered in Plastids
7.1. Stromule Tip Shedding
7.2. Direct Membrane Contacts and Vesicle Budding from the Plastid Envelope Membrane
7.3. Channels or Retrograde Protein Transporters
7.4. Changes in the Permeability of the Plastid Envelope
8. Outlook
Acknowledgements
References
- Bouvier, F.; Mialoundama, A.S.; Camara, B. A Sentinel Role for Plastids. In The Chloroplast; Sandelius, A.S., Aronsson, H., Eds.; Springer: Heidelberg, Germany, 2009; pp. 267–292. [Google Scholar]
- Pfannschmidt, T.; Bräutigam, K.; Wagner, R.; Dietzel, L.; Schröter, Y.; Steiner, S.; Nykytenko, A. Potential regulation of gene expression in photosynthetic cells by redox and energy state: Approaches towards better understanding. Ann. Bot 2008, 103, 599–607. [Google Scholar]
- Foyer, C.H.; Noctor, G. Managing the cellular redox hub in photosynthetic organisms. Plant Cell Environ 2012, 35, 199–201. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Pogson, B.J.; Woo, N.S.; Förster, B.; Small, I.D. Plastid signalling to the nucleus and beyond. Trends Plant Sci 2008, 13, 602–609. [Google Scholar]
- Estavillo, G.M.; Crisp, P.A.; Pornsiriwong, W.; Wirtz, M.; Collinge, D.; Carrie, C.; Giraud, E.; Whelan, J.; David, P.; Javot, H.; et al. Evidence for a SAL1-PAP chloroplast retrograde pathway that functions in drought and high light signaling in Arabidopsis. Plant Cell 2011, 23, 3992–4012. [Google Scholar]
- Xiao, Y.; Savchenko, T.; Baidoo, E.E.; Chehab, W.E.; Hayden, D.M.; Tolstikov, V.; Corwin, J.A.; Kliebenstein, D.J.; Keasling, J.D.; Dehesh, K. Retrograde signaling by the plastidial metabolite MEcPP regulates expression of nuclear stress-response genes. Cell 2012, 149, 1525–1535. [Google Scholar]
- Ramel, F.; Birtic, S.; Ginies, C.; Soubigou-Taconnat, L.; Triantaphylidès, C.; Havaux, M. Carotenoid oxidation products are stress signals that mediate gene responses to singlet oxygen in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5535–5540. [Google Scholar]
- Guttman, D.S.; Vinatzer, B.A.; Sarkar, S.F.; Ranall, M.V.; Kettler, G.; Greenberg, J.T. A functional screen for the type III (Hrp) secretome of the plant pathogen Pseudomonas syringae. Science 2002, 295, 1722–1726. [Google Scholar]
- Qi, J.; Zhou, G.; Yang, L.; Erb, M.; Lu, Y.; Sun, X.; Cheng, J.; Lou, Y. The chloroplast-localized phospholipases D α4 and α5 regulate herbivore-induced direct and indirect defenses in rice. Plant Physiol 2011, 157, 1987–1999. [Google Scholar]
- Morker, K.H.; Roberts, M.R. Light exerts multiple levels of influence on the Arabidopsis wound response. Plant Cell Environ 2011, 34, 717–728. [Google Scholar] [Green Version]
- Krause, K.; Krupinska, K. Nuclear regulators with a second home in organelles. Trends Plant Sci 2009, 14, 194–199. [Google Scholar]
- Sun, X.; Feng, P.; Xu, X.; Guo, H.; Ma, J.; Chi, W.; Lin, R.; Lu, C.; Zhang, L. A chloroplast envelope-bound PHD transcription factor mediates chloroplast signals to the nucleus. Nat. Commun 2011, 2, 477. [Google Scholar]
- Silva-Filho, M.C. One ticket for multiple destinations: Dual targeting of proteins to distinct subcellular locations. Curr. Opin. Plant Biol 2003, 6, 589–595. [Google Scholar]
- Regev-Rudzki, N.; Pines, O. Eclipsed distribution: A phenomenon of dual targeting of proteins and its significance. Bioessays 2007, 29, 772–782. [Google Scholar]
- Reape, T.J.; McCabe, P.F. Apoptotic-like regulation of programmed cell death in plants. Apoptosis 2010, 15, 249–256. [Google Scholar]
- Isemer, R.; Mulisch, M.; Schäfer, A.; Kirchner, S.; Koop, H.U.; Krupinska, K. Recombinant Whirly1 translocates from transplastomic chloroplasts to the nucleus. FEBS Lett 2012, 586, 85–88. [Google Scholar]
- Small, I.; Wintz, H.; Akashi, K.; Mireau, H. Two birds with one stone: Genes that encode products targeted to two or more compartments. Plant Mol. Biol 1998, 38, 265–277. [Google Scholar]
- Luo, M.; Orsi, R.; Patrucco, E.; Pancaldi, S.; Cella, R. Multiple transcription start sites of the carrot dihydrofolate reductase-thymidylate synthase gene, and sub-cellular localization of the bifunctional protein. Plant Mol. Biol 1997, 33, 709–722. [Google Scholar]
- Duchêne, A.M.; Giritch, A.; Hoffmann, B.; Cognat, V.; Lancelin, D.; Peeters, N.M.; Zaepfel, M.; Maréchal-Drouard, L.; Small, I.D. Dual targeting is the rule for organellar aminoacyl-tRNA synthetases in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2005, 102, 16484–16489. [Google Scholar]
- Ohta, M.; Sugita, M.; Sugiura, M. Three types of nuclear genes encoding chloroplast RNA-binding proteins (cp29, cp31 and cp33) are present in Arabidopsis thaliana: Presence of cp31 in chloroplasts and its homologue in nuclei/cytoplasms. Plant Mol. Biol 1995, 27, 529–539. [Google Scholar]
- Meier, I.; Phelan, T.; Gruissem, W.; Spiker, S.; Schneider, D. MFP1, a novel plant filament-like protein with affinity for matrix attachment region DNA. Plant Cell 1996, 8, 2105–2115. [Google Scholar]
- Boyle, B.; Brisson, N. Repression of the defense gene PR-10a by the single-stranded DNA binding protein SEBF. Plant Cell 2001, 13, 2525–2537. [Google Scholar]
- Kwon, C.; Chung, I.K. Interaction of an Arabidopsis RNA-binding protein with plant single-stranded telomeric DNA modulates telomerase activity. J. Biol. Chem 2004, 279, 12812–12818. [Google Scholar]
- Gindullis, F.; Meier, I. Matrix attachment region binding protein MFP1 is localized in discrete domains at the nuclear envelope. Plant Cell 1999, 11, 1117–1128. [Google Scholar]
- Samaniego, R.; Jeong, S.Y.; Meier, I.; de la Espina, S.M. Dual location of MAR-binding, filament-like protein 1 in Arabidopsis, tobacco, and tomato. Planta 2006, 223, 1201–1206. [Google Scholar]
- Ruwe, H.; Kupsch, C.; Teubner, M.; Schmitz-Linneweber, C. The RNA-recognition motif in chloroplasts. J. Plant Physiol 2011, 168, 1361–1371. [Google Scholar]
- Yoo, H.H.; Kwon, C.; Chung, I.K. An Arabidopsis splicing RNP variant STEP1 regulates telomere length homeostasis by restricting access of nuclease and telomerase. Mol. Cells 2010, 30, 279–283. [Google Scholar]
- Waldmüller, S.; Müller, U.; Link, G. GSBF1, a seedling-specific bZIP DNA-binding protein with preference for a ‘split’ G-box-related element in Brassica napus RbcS promoters. Plant Mol. Biol 1996, 32, 631–639. [Google Scholar]
- Sato, N.; Ohshima, K.; Watanabe, A.; Ohta, N.; Nishiyama, Y.; Joyard, J.; Douce, R. Molecular characterization of the PEND protein, a novel bZIP protein present in the envelope membrane that is the site of nucleoid replication in developing plastids. Plant Cell 1998, 10, 859–872. [Google Scholar]
- Terasawa, K.; Sato, N. Plastid localization of the PEND protein is mediated by a noncanonical transit peptide. FEBS J 2009, 276, 1709–1719. [Google Scholar]
- Ma, Z.; Dooner, H.K. A mutation in the nuclear-encoded plastid ribosomal protein S9 leads to early embryo lethality in maize. Plant J 2004, 37, 92–103. [Google Scholar]
- Grabowski, E.; Miao, Y.; Mulisch, M.; Krupinska, K. Single-stranded DNA-binding protein Whirly1 in barley leaves is located in plastids and the nucleus of the same cell. Plant Physiol 2008, 147, 1800–1804. [Google Scholar]
- Krause, K.; Kilbienski, I.; Mulisch, M.; Rödiger, A.; Schäfer, A.; Krupinska, K. DNA-binding proteins of the Whirly family in Arabidopsis thaliana are targeted to the organelles. FEBS Lett 2005, 579, 3707–3712. [Google Scholar]
- Desveaux, D.; Després, C.; Joyeux, A.; Subramaniam, R.; Brisson, N. PBF-2 is a novel single-stranded DNA binding factor implicated in PR-10a gene activation in potato. Plant Cell 2000, 12, 1477–1489. [Google Scholar]
- Yoo, H.H.; Kwon, C.; Lee, M.M.; Chung, I.K. Single-stranded DNA binding factor AtWHY1 modulates telomere length homeostasis in Arabidopsis. Plant J 2007, 49, 442–451. [Google Scholar]
- Raynaud, C.; Perennes, C.; Reuzeau, C.; Catrice, O.; Brown, S.; Bergounioux, C. Cell and plastid division are coordinated through the prereplication factor AtCDT1. Proc. Natl. Acad. Sci. USA 2005, 102, 8216–8221. [Google Scholar]
- Raynaud, C.; Sozzani, R.; Glab, N.; Domenichini, S.; Perennes, C.; Cella, R.; Kondorosi, E.; Bergounioux, C. Two cell-cycle regulated SET-domain proteins interact with proliferating cell nuclear antigen (PCNA) in Arabidopsis. Plant J 2006, 47, 395–407. [Google Scholar]
- Kodama, Y.; Sano, H. Evolution of a basic helix-loop-helix protein from a transcriptional repressor to a plastid-resident regulatory factor: Involvement in hypersensitive cell death in tobacco plants. J. Biol. Chem 2006, 281, 35369–35380. [Google Scholar]
- Kodama, Y.; Sano, H. Functional diversification of a basic helix-loop-helix protein due to alternative transcription during generation of amphidiploidy in tobacco plants. Biochem. J 2007, 403, 493–499. [Google Scholar]
- Pfalz, J.; Liere, K.; Kandlbinder, A.; Dietz, K.J.; Oelmüller, R. pTAC2, -6, and -12 are components of the transcriptionally active plastid chromosome that are required for plastid gene expression. Plant Cell 2006, 18, 176–197. [Google Scholar]
- Chen, M.; Galvão, R.M.; Li, M.; Burger, B.; Bugea, J.; Bolado, J.; Chory, J. Arabidopsis HEMERA/pTAC12 initiates photomorphogenesis by phytochromes. Cell 2010, 141, 1230–1240. [Google Scholar]
- Schwacke, R.; Fischer, K.; Ketelsen, B.; Krupinska, K.; Krause, K. Comparative survey of plastid and mitochondrial targeting properties of transcription factors in Arabidopsis and rice. Mol. Genet. Genomics 2007, 277, 631–646. [Google Scholar]
- Galichet, A.; Hoyerova, K.; Kamínek, M.; Gruissem, W. Farnesylation directs AtIPT3 subcellular localization and modulates cytokinin biosynthesis in Arabidopsis. Plant Physiol 2008, 146, 1155–1164. [Google Scholar]
- Caplan, J.L.; Mamillapalli, P.; Burch-Smith, T.M.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplastic protein NRIP1 mediates innate immune receptor recognition of a viral effector. Cell 2008, 132, 449–462. [Google Scholar]
- Lai, Z.; Li, Y.; Wang, F.; Cheng, Y.; Fan, B.; Yu, J.Q.; Chen, Z. Arabidopsis sigma factor binding proteins are activators of the WRKY33 transcription factor in plant defense. Plant Cell 2011, 23, 3824–3841. [Google Scholar]
- Inze, A.; Vanderauwera, S.; Hoeberichts, F.A.; Vandorpe, M.; van Gaever, T.; van Breusegem, F. A subcellular localization compendium of hydrogen peroxide-induced proteins. Plant Cell Environ 2012, 35, 308–320. [Google Scholar]
- Melonek, J.; Matros, A.; Trösch, M.; Mock, H.P.; Krupinska, K. The core of chloroplast nucleoids contains architectural SWIB-domain proteins. Plant Cell 2012, 24, 3060–3073. [Google Scholar]
- Xiong, J.Y.; Lai, C.X.; Qu, Z.; Yang, X.Y.; Qin, X.H.; Liu, G.Q. Recruitment of AtWHY1 and AtWHY3 by a distal element upstream of the kinesin gene AtKP1 to mediate transcriptional repression. Plant Mol. Biol 2009, 71, 437–449. [Google Scholar]
- Padmanabhan, M.S.; Dinesh-Kumar, S.P. All hands on deck-the role of chloroplasts, endoplasmic reticulum, and the nucleus in driving plant innate immunity. Mol. Plant-Microbe Interact 2010, 23, 1368–1380. [Google Scholar]
- Fu, Z.Q.; Guo, M.; Jeong, B.R.; Tian, F.; Elthon, T.E.; Cerny, R.L.; Staiger, D.; Alfano, J.R. A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature 2007, 447, 284–288. [Google Scholar]
- Desveaux, D.; Maréchal, A.; Brisson, N. Whirly transcription factors: Defense gene regulation and beyond. Trends Plant Sci 2005, 10, 95–102. [Google Scholar]
- Kakizaki, T.; Yazu, F.; Nakayama, K.; Ito-Inaba, Y.; Inaba, T. Plastid signalling under multiple conditions is accompanied by a common defect in RNA editing in plastids. J. Exp. Bot 2012, 63, 251–260. [Google Scholar]
- Rapp, J.C.; Mullet, J.E. Chloroplast transcription is required to express the nuclear genes rbcS and cab. Plastid DNA copy number is regulated independently. Plant Mol. Biol 1991, 17, 813–823. [Google Scholar]
- Melonek, J.; Mulisch, M.; Schmitz-Linneweber, C.; Grabowski, E.; Hensel, G.; Krupinska, K. Whirly1 in chloroplasts associates with intron containing RNAs and rarely co-localizes with nucleoids. Planta 2010, 232, 471–481. [Google Scholar]
- Prikryl, J.; Watkins, K.P.; Friso, G.; van Wijk, K.J.; Barkan, A. A member of the Whirly family is a multifunctional RNA- and DNA-binding protein that is essential for chloroplast biogenesis. Nucleic Acids Res 2008, 36, 5152–5165. [Google Scholar]
- Maréchal, A.; Parent, J.S.; Véronneau-Lafortune, F.; Joyeux, A.; Lang, B.F.; Brisson, N. Whirly proteins maintain plastid genome stability in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 14693–14698. [Google Scholar]
- Cappadocia, L.; Maréchal, A.; Parent, J.S.; Lepage, E.; Sygusch, J.; Brisson, N. Crystal structures of DNA-Whirly complexes and their role in Arabidopsis organelle genome repair. Plant Cell 2010, 22, 1849–1867. [Google Scholar]
- Majeran, W.; Friso, G.; Asakura, Y.; Qu, X.; Huang, M.; Ponnala, L.; Watkins, K.P.; Barkan, A.; van Wijk, K.J. Nucleoid-enriched proteomes in developing plastids and chloroplasts from maize leaves: A new conceptual framework for nucleoid functions. Plant Physiol 2012, 158, 156–189. [Google Scholar]
- Singh, B.; Gupta, R.S. Mitochondrial import of human and yeast fumarase in live mammalian cells: Retrograde translocation of the yeast enzyme is mainly caused by its poor targeting sequence. Biochem. Biophys. Res. Commum 2006, 346, 911–918. [Google Scholar]
- Valgardsdottir, R.; Brede, G.; Eide, L.G.; Frengen, E.; Prydz, H. Cloning and characterization of MDDX28, a putative dead-box helicase with mitochondrial and nuclear localization. J. Biol. Chem 2001, 276, 32056–32063. [Google Scholar]
- Settles, A.M.; Martienssen, R. Old and new pathways of protein export in chloroplasts and bacteria. Trends Cell Biol 1998, 8, 494–501. [Google Scholar]
- Köhler, R.H.; Cao, J.; Zipel, W.R.; Webb, W.W.; Hanson, M.R. Exchange of protein molecules through connections between higher plant plastids. Science 1997, 276, 2039–2042. [Google Scholar]
- Gray, J.C.; Sullivan, J.A.; Hibberd, J.M.; Hanson, M.R. Stromules: Mobile protrusions and interconnections between plastids. Plant Biol 2001, 3, 223–233. [Google Scholar]
- Gunning, B.E. Plastid stromules: Video microscopy of their outgrowth, retraction, tensioning, anchoring, branching, bridging, and tip-shedding. Protoplasma 2005, 225, 33–42. [Google Scholar]
- Schattat, M.; Barton, K.; Baudisch, B.; Klösgen, R.B.; Mathur, J. Plastid stromule branching coincides with contiguous endoplasmic reticulum dynamics. Plant Physiol 2011, 155, 1667–1677. [Google Scholar]
- Kwok, E.Y.; Hanson, M.R. GFP-labelled Rubisco and aspartate aminotransferase are present in plastid stromules and traffic between plastids. J. Exp. Bot 2004, 55, 595–604. [Google Scholar]
- Kwok, E.Y.; Hanson, M.R. Plastids and stromules interact with the nucleus and cell membrane in vascular plants. Plant Cell Rep 2004, 23, 188–195. [Google Scholar]
- Schattat, M.H.; Griffiths, S.; Mathur, N.; Barton, K.; Wozny, M.R.; Dunn, N.; Greenwood, J.S.; Mathur, J. Differential coloring reveals that plastids do not form networks for exchanging macromolecules. Plant Cell 2012, 24, 1465–1477. [Google Scholar]
- Natesan, S.K.; Sullivan, J.A.; Gray, J.C. Stromules: A characteristic cell-specific feature of plastid morphology. J. Exp. Bot 2005, 56, 787–797. [Google Scholar]
- Waters, M.T.; Fray, R.G.; Pyke, K.A. Stromule formation is dependent upon plastid size, plastid differentiation status and the density of plastids within the cell. Plant J 2004, 39, 655–667. [Google Scholar]
- Gray, J.C.; Hansen, M.R.; Shaw, D.J.; Graham, K.; Dale, R.; Smallman, P.; Natesan, S.K.; Newell, C.A. Plastid stromules are induced by stress treatments acting through abscisic acid. Plant J 2012, 69, 387–398. [Google Scholar]
- Wada, S.; Ishida, H.; Izumi, M.; Yoshimoto, K.; Ohsumi, Y.; Mae, T.; Makino, A. Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves. Plant Physiol 2009, 149, 885–893. [Google Scholar]
- Martínez, D.E.; Costa, M.L.; Guiamet, J.J. Senescence-associated degradation of chloroplast proteins inside and outside the organelle. Plant Biol 2008, 10, S15–S22. [Google Scholar]
- Andersson, M.X.; Goksör, M.; Sandelius, A.S. Optical manipulation reveals strong attracting forces at membrane contact sites between endoplasmic reticulum and chloroplasts. J. Biol. Chem 2007, 282, 1170–1174. [Google Scholar]
- Crotty, W.J.; Ledbetter, M.C. Membrane continuities involving chloroplasts and other organelles in plant cells. Science 1973, 182, 839–841. [Google Scholar]
- Whatley, J.M.; Mclean, B.; Juniper, B.E. Continuity of chloroplast and endoplasmic-reticulum membranes in Phaseolus vulgaris. New Phytol 1991, 117, 209–217. [Google Scholar]
- Soltys, B.J.; Gupta, R.S. Mitochondrial-matrix proteins at unexpected locations: Are they exported? Trends Biochem. Sci 1999, 24, 174–177. [Google Scholar]
- Andrade-Navarro, M.A.; Sanchez-Pulido, L.; McBride, H.M. Mitochondrial vesicles: An ancient process providing new links to peroxisomes. Curr. Opin. Cell Biol 2009, 21, 560–567. [Google Scholar]
- Mashburn-Warren, L.M.; Whiteley, M. Special delivery: Vesicle trafficking in prokaryotes. Mol. Microbiol 2006, 61, 839–846. [Google Scholar]
- Zheng, W.; Bergman, B.; Chen, B.; Zheng, S.; Guan, X.; Rasmussen, U. Cellular responses in the cyanobacterial symbiont during its vertical transfer between plant generations in the Azolla microphylla symbiosis. New Phytol 2009, 181, 53–61. [Google Scholar]
- Neuspiel, M.; Schauss, A.C.; Braschi, E.; Zunino, R.; Rippstein, P.; Rachubinski, R.A.; Andrade-Navarro, M.A.; McBride, H.M. Cargo-selected transport from the mitochondria to peroxisomes is mediated by vesicular carriers. Curr. Biol 2008, 18, 102–108. [Google Scholar]
- Augustin, S.; Nolden, M.; Müller, S.; Hardt, O.; Arnold, I.; Langer, T. Characterization of peptides released from mitochondria: Evidence for constant proteolysis and peptide efflux. J. Biol. Chem 2005, 280, 2691–2699. [Google Scholar]
- Young, L.; Leonhard, K.; Tatsuta, T.; Trowsdale, J.; Langer, T. Role of the ABC transporter Mdl1 in peptide export from mitochondria. Science 2001, 291, 2135–2138. [Google Scholar]
- Reits, E.A.; Vos, J.C.; Gromme, M.; Neefjes, J. The major substrates for TAP in vivo are derived from newly synthesized proteins. Nature 2000, 404, 774–778. [Google Scholar]
- Schneewind, O.; Missiakas, D.M. Protein secretion and surface display in Gram-positive bacteria. Philos. Trans. R. Soc. B 2012, 367, 1123–1139. [Google Scholar]
- Yusa, F.; Steiner, J.M.; Löffelhardt, W. Evolutionary conservation of dual Sec translocases in the cyanelles of Cyanophora paradoxa. BMC Evol. Biol 2008, 8, 304. [Google Scholar]
- Raghavan, S.; Manzanillo, P.; Chan, K.; Dovey, C.; Cox, J.S. Secreted transcription factor controls Mycobacterium tuberculosis virulence. Nature 2008, 454, 717–721. [Google Scholar]
- De Crécy-Lagard, V.; Hanson, A.D. Finding novel metabolic genes through plant-prokaryote phylogenomics. Trends Microbiol 2007, 15, 563–570. [Google Scholar]
- Roston, R.; Gao, J.; Xu, C.; Benning, C. Arabidopsis chloroplast lipid transport protein TGD2 disrupts membranes and is part of a large complex. Plant J 2011, 66, 759–769. [Google Scholar]
- Arpagaus, S.; Rawyler, A.; Braendle, R. Occurrence and characteristics of the mitochondrial permeability transition in plants. J. Biol. Chem 2002, 277, 1780–1787. [Google Scholar]
- Mulisch, M.; Krupinska, K. Ultrastructural Analyses of Senescence Asssociated Dismantling of Chloroplasts Revisited. In Plastid Development In Leaves During Growth and Senescence; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; AIPR, Springer: Dordrecht, The Netherlands; p. 2012.



{kind=link}
{kind=link}
| First described | Protein name(s) | Protein function | References | Release from plastids |
|---|---|---|---|---|
| 1995 | cp29B (p) SEBF (n) | RNA-binding protein transcriptional repressor | [21,27] [23] | not investigated |
| 1995 | cp31A (p) STEP1 (n) | RNA-binding protein telomere-binding | [21,27] [24,28] | not investigated |
| 1996 | MFP1 (n) MFP1 (p) | matrix attachment region binding nucleoid associated protein | [22] [26] | not investigated |
| 1996 | GSBF1 (n) PEND (p) | transcription factor nucleoid associated protein | [29] [30] | possible [31] |
| 1997 | DHFR (p + n) | dihydrofolate reductase/thymidylate synthase | [19] | no [19] |
| 2004 | LEM1 (p + n) | unknown (homologous to plastid ribosomal protein PRPS9) | [32] | not investigated |
| 2005 | WHIRLY1 (p) WHIRLY1 (n) | DNA + RNA binding; DNA maintenance transcriptional activator; telomere-binding | [33,34] [35,36] | yes [17] |
| 2005 | CDT1 (p) CDT1 (n) | plastid division (interaction with Arc6) DNA replication | [37] [37] | not investigated |
| 2006 | ATXR5 (p + n) | control of cell cycle and DNA replication in the nucleus; plastid function unknown | [38] | not investigated |
| 2006 | NtWIN4 (p) NtWIN4 (n) | induction of hypersensitive cell death transcriptional repressor | [39] [39] | no [40] |
| 2006 | pTAC12 (p) HEMERA (n) | nucleoid associated protein phytochrome signalling | [41] [42] | not investigated |
| 2007 | At2g44940 (p + n) | transcription factor with AP2 DNA binding motif | [43] | not investigated |
| 2008 | IPT3 (p + n) | cytokinin biosynthesis in plastids; nuclear function unknown | [44] | no [44] |
| 2008 | NRIP1 (p + n) | rhodanese sulfur transferase; immune receptor recognition; plastid function unknown | [45] | possible [45] |
| 2011 | SIB1, SIB2 (p + n) | proteins binding to Sigma factor1 of plastid encoded RNA-polymerase | [46] | not investigated |
| 2012 | ANAC102 (p + n) | NAC transcription factor | [47] | not investigated |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Krause, K.; Oetke, S.; Krupinska, K. Dual Targeting and Retrograde Translocation: Regulators of Plant Nuclear Gene Expression Can Be Sequestered by Plastids. Int. J. Mol. Sci. 2012, 13, 11085-11101. https://doi.org/10.3390/ijms130911085
Krause K, Oetke S, Krupinska K. Dual Targeting and Retrograde Translocation: Regulators of Plant Nuclear Gene Expression Can Be Sequestered by Plastids. International Journal of Molecular Sciences. 2012; 13(9):11085-11101. https://doi.org/10.3390/ijms130911085
Chicago/Turabian StyleKrause, Kirsten, Svenja Oetke, and Karin Krupinska. 2012. "Dual Targeting and Retrograde Translocation: Regulators of Plant Nuclear Gene Expression Can Be Sequestered by Plastids" International Journal of Molecular Sciences 13, no. 9: 11085-11101. https://doi.org/10.3390/ijms130911085
APA StyleKrause, K., Oetke, S., & Krupinska, K. (2012). Dual Targeting and Retrograde Translocation: Regulators of Plant Nuclear Gene Expression Can Be Sequestered by Plastids. International Journal of Molecular Sciences, 13(9), 11085-11101. https://doi.org/10.3390/ijms130911085