Proteome and Peptidome of Human Acquired Enamel Pellicle on Deciduous Teeth

Abstract
:1. Introduction
2. Results and Discussion
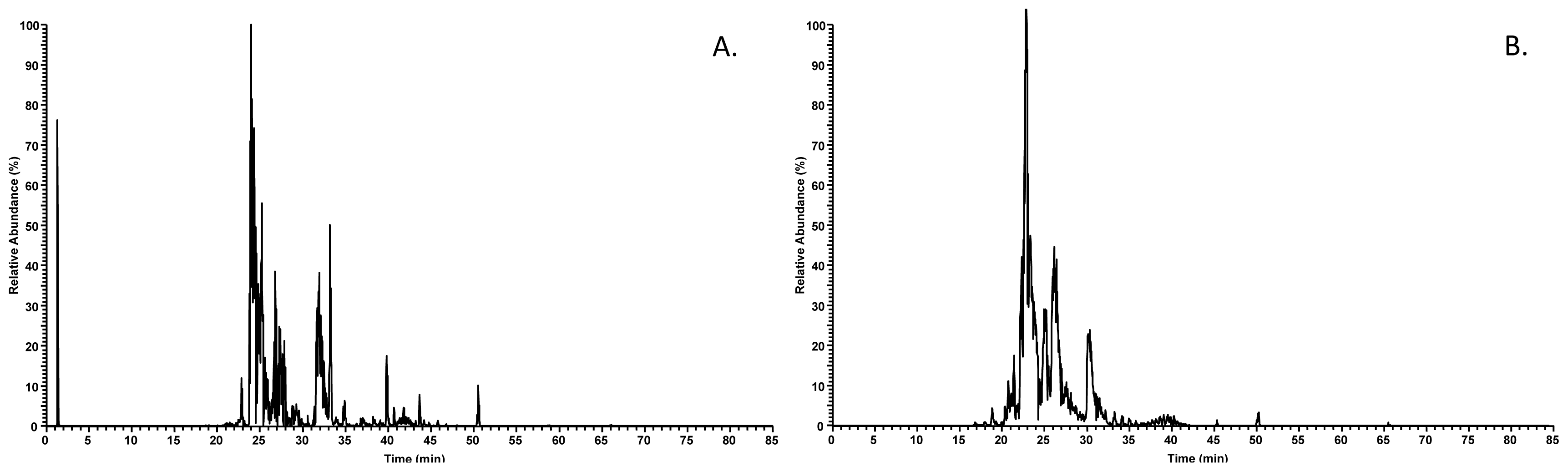
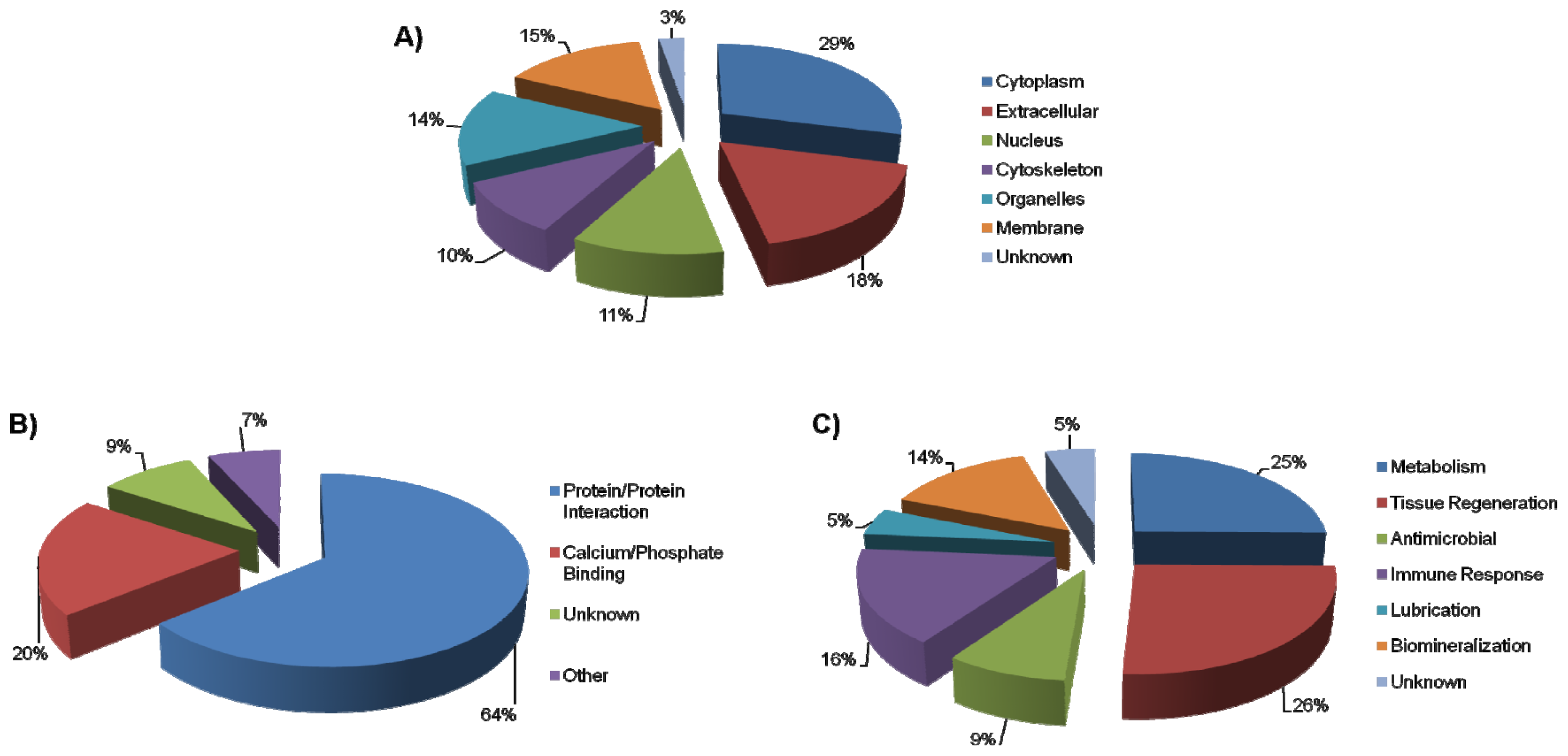
2.1. AEP Proteome
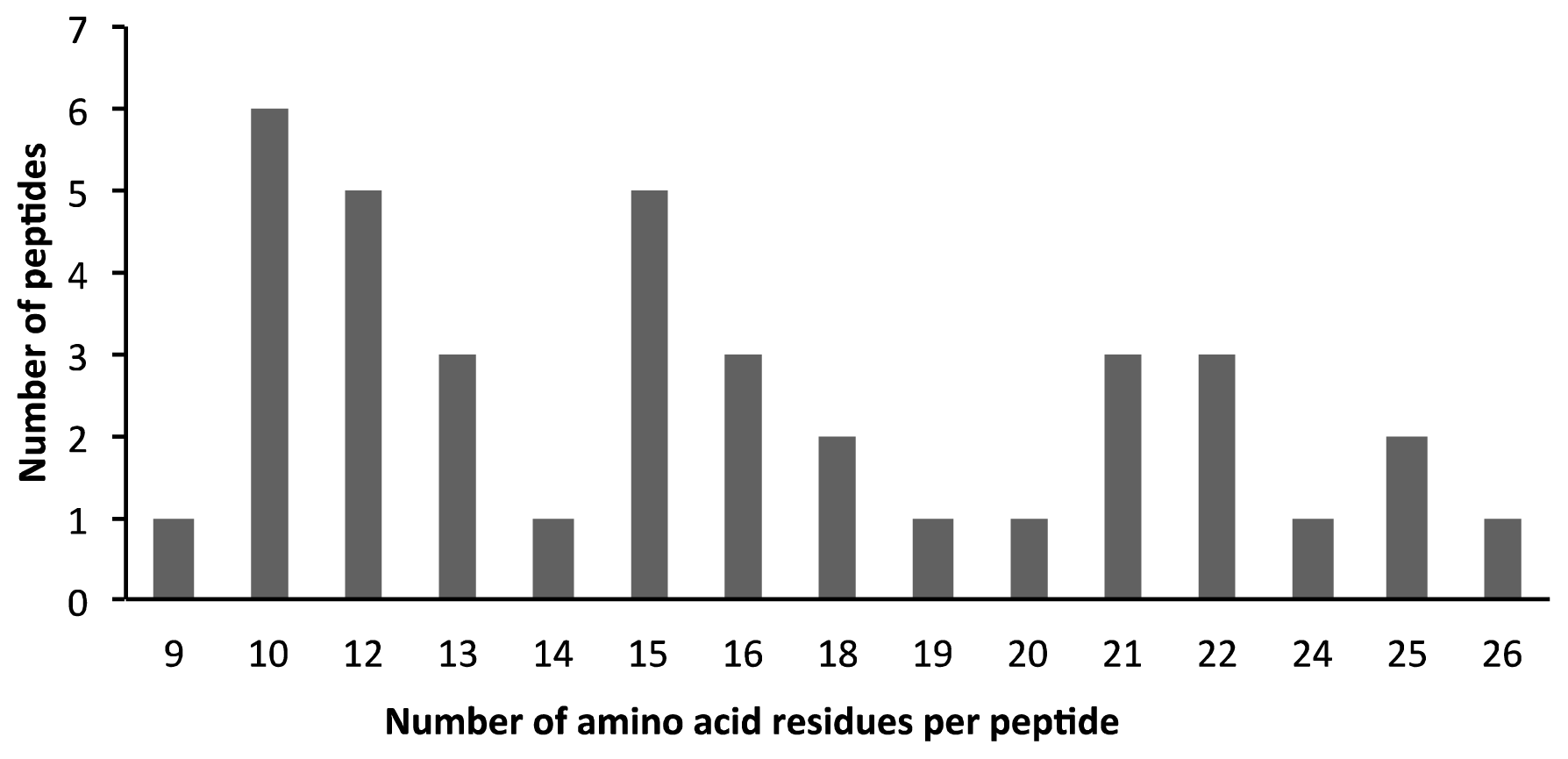
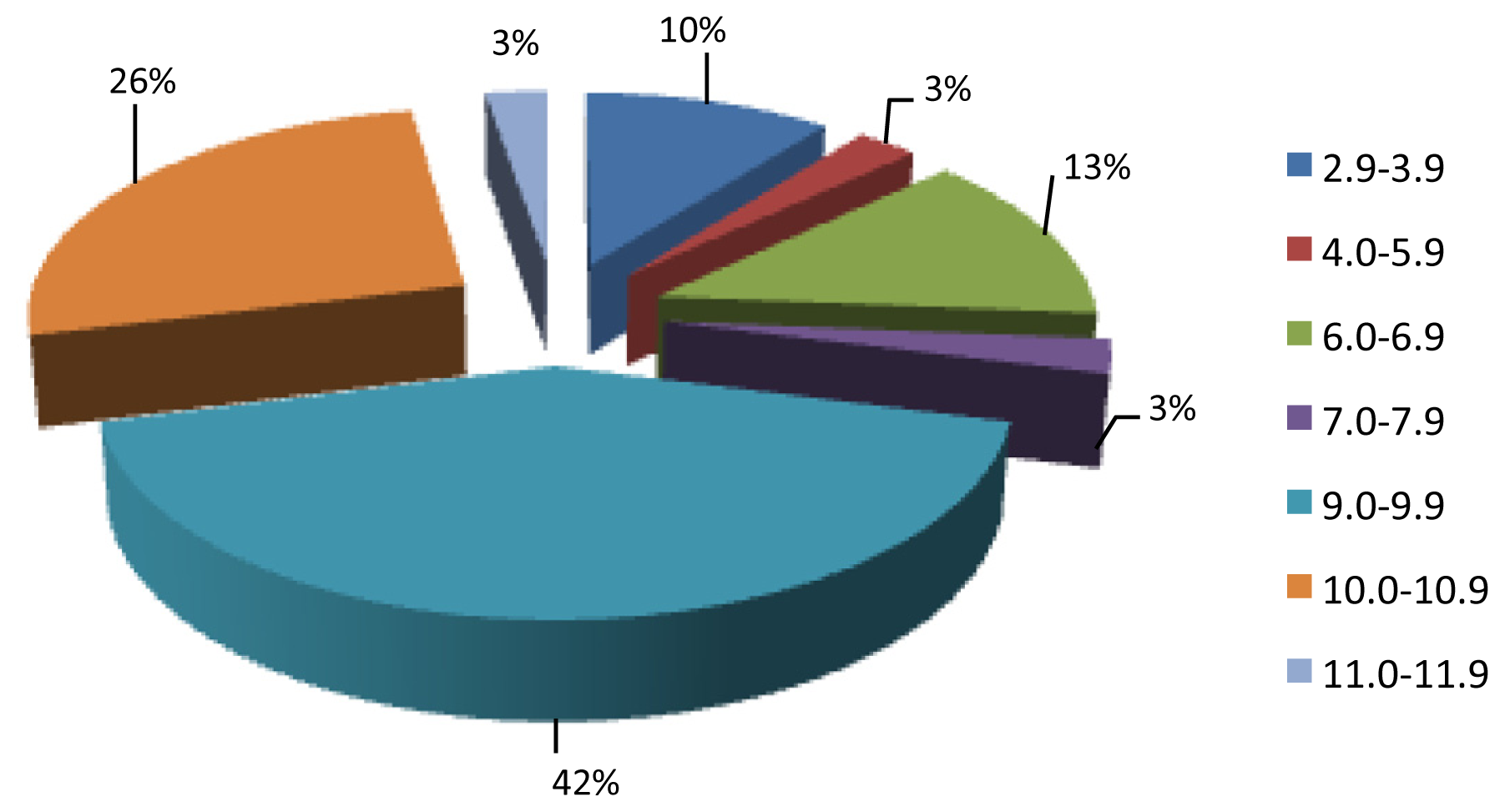
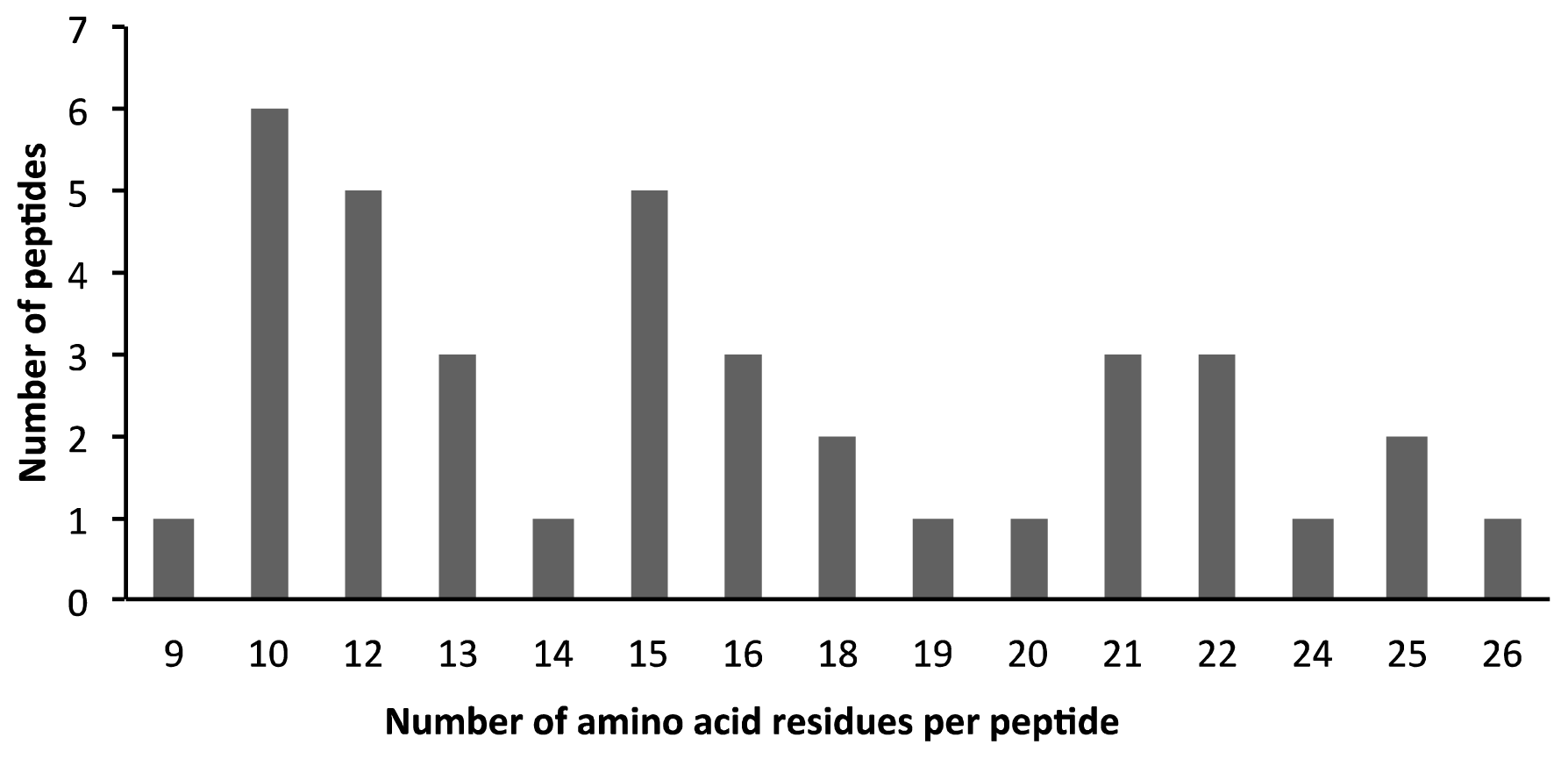
2.2. AEP Peptidome
3. Experimental Section
3.1. Human Subjects
3.2. AEP Collection
3.3. AEP Elution by Sonication
3.4. Deciduous AEP Proteome
3.5. In-Solution Digestion
3.6. Deciduous AEP Peptidome
3.7. LC-ESI-MS/MS Analyses
3.8. Peptide and Protein Identification
3.9. Protein Annotations
4. Conclusions
Supplementary Information
ijms-14-00920-s001.pdfAcknowledgments
References
- Hannig, M.; Joiner, A. The structure, function and properties of the acquired pellicle. Monogr. Oral Sci 2006, 19, 29–64. [Google Scholar]
- Siqueira, W.L.; Helmerhorst, E.J.; Zhang, W.; Salih, E.; Oppenheim, F.G. Acquired enamel pellicle and its potential role in oral diagnostics. Ann. N. Y. Acad. Sci 2007, 1098, 504–509. [Google Scholar]
- Rudiger, S.G.; Carlen, A.; Meurman, J.H.; Kari, K.; Olsson, J. Dental biofilms at healthy and inflamed gingival margins. J. Clin. Periodontol 2002, 29, 524–530. [Google Scholar]
- Siqueira, W.L.; Zhang, W.; Helmerhorst, E.J.; Gygi, S.P.; Oppenheim, F.G. Identification of protein components in in vivo human acquired enamel pellicle using LC-ESI-MS/MS. J. Proteome Res 2007, 6, 2152–2160. [Google Scholar]
- Vitorino, R.; Calheiros-Lobo, M.J.; Williams, J.; Ferrer-Correia, A.J.; Tomer, K.B.; Duarte, J.A.; Domingues, P.M.; Amado, F.M. Peptidomic analysis of human acquired enamel pellicle. Biomed. Chromatogr 2007, 21, 1107–1117. [Google Scholar]
- Siqueira, W.L.; Oppenheim, F.G. Small molecular weight proteins/peptides present in the in vivo formed human acquired enamel pellicle. Arch. Oral Biol 2009, 54, 437–444. [Google Scholar]
- Helmerhorst, E.J.; Alagl, A.S.; Siqueira, W.L.; Oppenheim, F.G. Oral fluid proteolytic effects on histatin 5 structure and function. Arch. Oral Biol 2006, 51, 1061–1070. [Google Scholar]
- Siqueira, W.L.; Bakkal, M.; Xiao, Y.; Sutton, J.N.; Mendes, F.M. Quantitative proteomic analysis of the effect of fluoride on the acquired enamel pellicle. PLoS One 2012, 7, e42204. [Google Scholar]
- Yao, Y.; Berg, E.A.; Costello, C.E.; Troxler, R.F.; Oppenheim, F.G. Identification of protein components in human acquired enamel pellicle and whole saliva using novel proteomics approaches. J. Biol. Chem 2003, 278, 5300–5308. [Google Scholar]
- Ripa, L.W.; Gwinnett, A.J.; Buonocore, M.G. The “prismless” outer layer of deciduous and permanent enamel. Arch. Oral Biol 1966, 11, 41–48. [Google Scholar]
- Gwinnett, A.J. The ultrastructure of the “prismless” enamel of permanent human teeth. Arch. Oral Biol 1967, 12, 381–388. [Google Scholar]
- Lussi, A.; Kohler, N.; Zero, D.; Schaffner, M.; Megert, B. A comparison of the erosive potential of different beverages in primary and permanent teeth using an in vitro model. Eur. J. Oral Sci 2000, 108, 110–114. [Google Scholar]
- Sonju Clasen, A.B.; Hannig, M.; Skjorland, K.; Sonju, T. Analytical and ultrastructural studies of pellicle on primary teeth. Acta odontol. Scand 1997, 55, 339–343. [Google Scholar]
- Hay, D.I.; Bennick, A.; Schlesinger, D.H.; Minaguchi, K.; Madapallimattam, G.; Schluckebier, S.K. The primary structures of six human salivary acidic proline-rich proteins (PRP-1, PRP-2, PRP-3, PRP-4, PIF-s and PIF-f). Biochem. J 1988, 255, 15–21. [Google Scholar]
- Moreno, E.C.; Varughese, K.; Hay, D.I. Effect of human salivary proteins on the precipitation kinetics of calcium phosphate. Calcif. Tissue Int 1979, 28, 7–16. [Google Scholar]
- Oppenheim, F.G.; Yang, Y.C.; Diamond, R.D.; Hyslop, D.; Offner, G.D.; Troxler, R.F. The primary structure and functional characterization of the neutral histidine-rich polypeptide from human parotid secretion. J. Biol. Chem 1986, 261, 1177–1182. [Google Scholar]
- Siqueira, W.L.; Salih, E.; Wan, D.L.; Helmerhorst, E.J.; Oppenheim, F.G. Proteome of human minor salivary gland secretion. J. Dent. Res 2008, 87, 445–450. [Google Scholar]
- Denny, P.; Hagen, F.K.; Hardt, M.; Liao, L.; Yan, W.; Arellanno, M.; Bassilian, S.; Bedi, G.S.; Boontheung, P.; Cociorva, D.; et al. The proteomes of human parotid and submandibular/sublingual gland salivas collected as the ductal secretions. J. Proteome Res 2008, 7, 1994–2006. [Google Scholar]
- Dawes, C. Estimates, from salivary analyses, of the turnover time of the oral mucosal epithelium in humans and the number of bacteria in an edentulous mouth. Arch. Oral Biol 2003, 48, 329–336. [Google Scholar]
- Siqueira, W.L.; Custodio, W.; McDonald, E.E. New insights into the composition and functions of the acquired enamel pellicle. J. Dent. Res 2012, 91, 1110–1118. [Google Scholar]
- Siqueira, W.L.; Margolis, H.C.; Helmerhorst, E.J.; Mendes, F.M.; Oppenheim, F.G. Evidence of intact histatins in the in vivo acquired enamel pellicle. J. Dent. Res 2010, 89, 626–630. [Google Scholar]
- Hay, D.I. Some observations on human saliva proteins and their role in the formation of the acquired enamel pellicle. J. Dent. Res 1969, 48, 806–810. [Google Scholar]
- Hannig, C.; Spies, B.; Spitzmuller, B.; Hannig, M. Efficacy of enzymatic mouth rinses for immobilisation of protective enzymes in the in situ pellicle. Arch. Oral Biol 2010, 55, 1–6. [Google Scholar]
- Taba, M., Jr; Kinney, J.; Kim, A.S.; Giannobile, W.V. Diagnostic biomarkers for oral and periodontal diseases. Dent. Clin. North Am. 2005, 49, 551–571. [Google Scholar]
- Angker, L.; Nockolds, C.; Swain, M.V.; Kilpatrick, N. Quantitative analysis of the mineral content of sound and carious primary dentine using BSE imaging. Arch. Oral Biol 2004, 49, 99–107. [Google Scholar]
- Cuy, J.L.; Mann, A.B.; Livi, K.J.; Teaford, M.F.; Weihs, T.P. Nanoindentation mapping of the mechanical properties of human molar tooth enamel. Arch. Oral Biol 2002, 47, 281–291. [Google Scholar]
- Vitorino, R.; Lobo, M.J.; Ferrer-Correira, A.J.; Dubin, J.R.; Tomer, K.B.; Domingues, P.M.; Amado, F.M. Identification of human whole saliva protein components using proteomics. Proteomics 2004, 4, 1109–1115. [Google Scholar]
- Amado, F.M.; Vitorino, R.M.; Domingues, P.M.; Lobo, M.J.; Duarte, J.A. Analysis of the human saliva proteome. Expert Rev. Proteomics 2005, 2, 521–539. [Google Scholar]
- Vitorino, R.; Guedes, S.; Manadas, B.; Ferreira, R.; Amado, F. Toward a standardized saliva proteome analysis methodology. J. Proteomics 2012, 75, 5140–5165. [Google Scholar]
- Bandhakavi, S.; Stone, M.D.; Onsongo, G.; Van Riper, S.K.; Griffin, T.J. A dynamic range compression and three-dimensional peptide fractionation analysis platform expands proteome coverage and the diagnostic potential of whole saliva. J. Proteome Res 2009, 8, 5590–5600. [Google Scholar]
- Zehetbauer, S.; Wojahn, T.; Hiller, K.A.; Schmalz, G.; Ruhl, S. Resemblance of salivary protein profiles between children with early childhood caries and caries-free controls. Eur. J. Oral Sci 2009, 117, 369–373. [Google Scholar]
- McDonald, E.E.; Goldberg, H.A.; Tabbara, N.; Mendes, F.M.; Siqueira, W.L. Histatin 1 resists proteolytic degradation when adsorbed to hydroxyapatite. J. Dent. Res 2011, 90, 268–272. [Google Scholar]
- Hay, D.I. The isolation from human parotid saliva of a tyrosine-rich acidic peptide which exhibits high affinity for hydroxyapatite surfaces. Arch. Oral Biol 1973, 18, 1531–1541. [Google Scholar]
- Hay, D.I. The interaction of human parotid salivary proteins with hydroxyapatite. Arch. Oral Biol 1973, 18, 1517–1529. [Google Scholar]
- Helmerhorst, E.J.; Sun, X.; Salih, E.; Oppenheim, F.G. Identification of Lys-Pro-Gln as a novel cleavage site specificity of saliva-associated proteases. J. Biol. Chem 2008, 283, 19957–19966. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession # | Protein Name |
|---|---|
| B7ZMD7 | Amylase, alpha 1A (Salivary) b, h, i, l, n, q |
| P02814 | Submaxillary gland androgen-regulated protein 3B b, k, r |
| F6KPG5 | Albumin (Fragment) a, b, e, f, h, i, l, m, q |
| P02533 | Keratin, Cytoskeletal 14 d, h, m |
| P08779 | Keratin, Cytoskeletal 16 a, d, e, h, m |
| B4DRW1 | Keratin, Cytoskeletal 4 a, d, h, m |
| A1A4E9 | Keratin, cytoskeletal 13 a, d, e, h, m |
| P02812 | Basic salivary proline-rich protein 2 a, d, h, l, o, p |
| H6VRF8 | Keratin 1d, h, l, o |
| P02808 | Statherin b, f, h, i, p, q |
| B5BU38 | Annexin a, b, c, d, e, f, h, i, l, m, o, q |
| P07476 | Involucrin d, f, h, l, m |
| B4DWU6 | cDNA FLJ51361, highly similar to Keratin, type II cytoskeletal 6A d, h, m |
| B4DVQ0 | cDNA FLJ58286, highly similar to Actin, cytoplasmic 2 a, f, h, m |
| P35908 | Keratin, type II cytoskeletal 2 epidermal a, b, h, m |
| C9JEV0 | Zinc-alpha-2-glycoprotein b, f, h, o |
| P02810 | Salivary acidic proline-rich phosphoprotein ½ b, f, h, i, q |
| P06702 | Calgranulin B a, b, c, d, e, f, h, i, n, o, q |
| P04280 | Basic salivary proline-rich protein 1 b, j, l, p |
| P13647 | Keratin, type II cytoskeletal 5 a, d, e, f, h, m |
| P01040 | Cystatin-A a, c, e, h, n |
| P01834 | Ig kappa chain C region b, f, h, o |
| P0CG05 | Ig lambda-2 chain C regions a, d, h, o |
| B7Z4X2 | Lactoferroxin-C b, i, l, q |
| P61626 | Lysozyme b, h, n, o |
| B2R4M6 | highly similar to Homo sapiens S100 calcium binding protein A9 a, c, d, i, l, m, o, q |
| P15515 | Histatin 1 c, h, n, q |
| P12273 | Prolactin-inducible protein b, h, r |
| B1AN48 | Small proline-rich protein 3 (Fragment) a, h, l, m |
| Q01546 | Keratin, type II cytoskeletal 2 oral d, h, m |
| P98088 | Mucin-5AC (Fragments) a, b, e, h, m, n, p, q |
| E7EQV5 | Actin, alpha skeletal muscle a, d, h, m |
| H0YKS4 | Annexin (Fragment) a, f, h, i, l, m, o, q |
| P01876 | Ig alpha-1 chain C region b, f, h, n, o |
| F8VV32 | Lysozyme C b, h, n, o |
| O60744 | Thioredoxin delta 3 (Fragment) a, b, c, e, h, l, m, o |
| P10163 | Basic salivary proline-rich protein 4 b, j, l, p |
| B4DIL4 | cDNA FLJ50166, highly similar to Dedicator of cytokinesis protein 6 a, h, o |
| B7Z7R8 | cDNA FLJ55622, highly similar to Multimerin-1 a, b, e, h, o |
| H0Y6K7 | Probable E3 ubiquitin-protein ligase HERC4 (Fragment) a, f, j, l, m |
| P31947 | 14-3-3 protein sigma OS=Homo sapiens a, b, c, h, l, m |
| P07108 | Acyl-CoA-binding protein a, e, h, l |
| D6RCA8 | Annexin (Fragment) b, f, h, i, l, m, o, q |
| H0YMD9 | Annexin A2 (Fragment) g, h, i, l, m, o, q |
| Q86VF0 | Beta-globin (Fragment) a, b, h, i, l, m, o, q |
| A0M8Q9 | C1 segment protein (Fragment) g, h, l, o |
| P27482 | Calmodulin-like protein 3 a, i, q |
| B4DWR5 | highly similar to Involucrin a, f, j, r |
| B4DGW2 | highly similar to Rho guanine nucleotide exchange factor 12 d, h, m |
| A8K5I6 | highly similar to Homo sapiens cornulin (CRNN) a, b, c, d, e, f, i, m, q |
| P04080 | Cystatin B b, h, n |
| P01036 | Cystatin S a, b, f, h, n |
| Q9UGM3 | Deleted in malignant brain tumors 1 protein b, h, i, n, q |
| G3V1R1 | HCG26567, isoform CRA_c a, c, d, e, f, j, p |
| F8WE04 | Heat shock protein beta-1 a, b, e, f, h, l |
| E5RG22 | Hemofiltrate peptide HF7665 (Fragment) b, h, m |
| A8K9J7 | Histone H2B g, k, m |
| H0Y3I2 | Lactoperoxidase a, b, c, e, i, n, q |
| P05164 | Myeloperoxidase a, b, c, e, i, n, q |
| C9J4S4 | Ras-related protein Rab-7a a, e, h, l, m |
| E9PBV3 | Suprabasin a, e, j, r |
| H0YDD8 | 60S acidic ribosomal protein P2 (Fragment) a, e, k, l, m |
| Q2MD48 | B-cell linker protein (Fragment) a, f, h, o |
| F8WBR5 | Calmodulin g, i, q |
| Q9Y6Y1 | Calmodulin-binding transcription activator 1 a, c, h, k, l |
| B4DT21 | cDNA FLJ50202, highly similar to Interleukin-12 receptor beta-2 chain f, h, l, m |
| H0YCD2 | DNA polymerase subunit gamma-1 (Fragment) a, e, f, h, k, l, m |
| G3V1M9 | HCG26567, isoform CRA_b b, j, l |
| O60393 | Homeobox protein NOBOX c, f, k, l, m |
| C9J0S5 | Lactoferroxin-C (Fragment) a, b, h, i, l, o, q |
| E9PNX2 | Neuronal acetylcholine receptor subunit alpha-10 f, h, m |
| Q59H51 | Pleckstrin homology domain-containing protein family A member 4 variant (Fragment) a, f, h, r |
| B7ZW15 | Putative uncharacterized protein a, h, l |
| F5H4J4 | Runt-related transcription factor 3 a, c, e, h, k, l, m |
| Q9BVH8 | VWA5B2 protein (Fragment) g, h, r |
| Accession | Protein name | Peptide sequence | # AA | XCorr | MS/MS charge | MW (Da) | pI | Net charge at pH 7 |
|---|---|---|---|---|---|---|---|---|
| P02808 | Statherin | FGYGYGPYQPVPEQP | 15 | 3.73 | 2 | 1699.59 | 3.3 | −1 |
| YGPYQPVPEQPLYPQP | 16 | 3.25 | 2 | 1874.25 | 3.3 | −1 | ||
| YQPVPEQPLYPQP | 13 | 3.24 | 2 | 1556.12 | 3.3 | −1 | ||
| P02810 | Salivary acidic proline-rich phosphoprotein 1/2 | GHQQGPPPPPPGKPQ | 15 | 3.17 | 3 | 1517.01 | 10.1 | 1.1 |
| GPPPQGGRPQ | 10 | 2.60 | 2 | 989.95 | 11 | 1 | ||
| GPPPQGGRPQGPPQGQSPQ | 19 | 4.05 | 2 | 1867.39 | 11 | 1 | ||
| GPPQQGGHPRPP | 12 | 2.65 | 2 | 1224.51 | 11 | 1.1 | ||
| GRPQGPPQGQSPQ | 13 | 3.32 | 2 | 1334.13 | 11 | 1 | ||
| QGPPQQGGHQQGPPPPPPGKPQ | 22 | 4.54 | 3 | 2211.75 | 10.1 | 1.1 | ||
| P02812 | Basic salivary proline-rich protein 2 | GNQPQGPPPP | 10 | 2.53 | 2 | 988.30 | 6.01 | 0 |
| GPPPPGKPQGPPPQ | 14 | 4.01 | 2 | 1351.15 | 10.1 | 1 | ||
| GPPSPPGKPQ | 10 | 2.59 | 2 | 961.79 | 10.1 | 1 | ||
| KPQGPPPPGKPQGPPPQGDK | 20 | 5.25 | 3 | 2004.77 | 10.3 | 2 | ||
| KPQGPPPPGKPQGPPPQGDNK | 21 | 3.40 | 3 | 2119.08 | 10.3 | 2 | ||
| PGKPQGPPPQGGSKSRSARSP | 21 | 2.71 | 2 | 2073.93 | 12.4 | 4 | ||
| PGKPQGPPPQGGSKSRSARSPPGKP | 25 | 2.74 | 2 | 2452.99 | 12.4 | 5 | ||
| PGKPQGPPPQGGSKSRSSRSPPGK | 24 | 3.14 | 2 | 2452.99 | 12.4 | 5 | ||
| P02814 | Submaxillary gland androgenregulated protein 3 | FGPGFVPPPPPPPYGPGR | 18 | 2.61 | 2 | 1834.38 | 9.8 | 1 |
| FVPPPPPPPYGPG | 13 | 3.11 | 2 | 1318.79 | 5.9 | 0 | ||
| GPGIFPPPPPQP | 12 | 2.95 | 2 | 1202.34 | 6 | 0 | ||
| GPLAPPQPFGPGFVPPPPPPPYGPGR | 26 | 3.27 | 3 | 2591.58 | 9.8 | 1 | ||
| P04080 | Cystatin-B | SQVVAGTNYFIK | 12 | 4.21 | 2 | 1327.52 | 9.7 | 1 |
| P04280 | Basic salivary proline-rich protein 1 | GGNKPQGPPPPPGKPQ | 16 | 3.37 | 3 | 1553.24 | 10.6 | 2 |
| GNKPQGPPPP | 10 | 2.63 | 2 | 987.61 | 10.1 | 1 | ||
| GNKPQGPPPPGKPQGPPPQGDK | 22 | 3.73 | 3 | 2175.93 | 10.3 | 2 | ||
| GNKPQGPPPPPGKPQ | 15 | 3.14 | 3 | 1496.85 | 10.6 | 2 | ||
| GNQPQGPPPP | 10 | 2.53 | 2 | 988.30 | 6 | 0 | ||
| GPPPPGKPQGPPAQG | 15 | 2.59 | 2 | 1381.12 | 10 | 1 | ||
| KPQGPPPPGKPQGPPAQGGSK | 21 | 4.01 | 3 | 2007.97 | 10.8 | 3 | ||
| PQGGNKPQGPPPPPGKPQ | 18 | 2.78 | 2 | 1779.23 | 10.6 | 2 | ||
| P06702 | S100-A9 | NIETIINTFHQYSVK | 15 | 4.61 | 2 | 1807.65 | 7.7 | 0.1 |
| NIETIINTFHQY | 12 | 3.27 | 2 | 1492.53 | 5.1 | −0.9 | ||
| P10163 | Basic salivary proline-rich protein 4 allele S | GNKPQGPPPP | 10 | 2.63 | 2 | 987.61 | 10 | 1 |
| LISGKPEGR | 9 | 2.84 | 2 | 956.46 | 10 | 1 | ||
| P15515 | Histatin 1 | EFPFYGDYGSNYLYDN | 16 | 5.24 | 2 | 1964.32 | 2.8 | −3 |
| REFPFYGDYGSN | 12 | 2.88 | 2 | 1452.20 | 4 | −1 | ||
| Q8TAX7 | Mucin-7 | PVNSPAPQDTTAAPPTPSATTP | 22 | 2.70 | 2 | 2277.10 | 3.1 | −1 |
| TSSSVATLAPVNSPAPQDTTAAPPT | 25 | 3.73 | 3 | 2621.38 | 3.1 | −1 | ||
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zimmerman, J.N.; Custodio, W.; Hatibovic-Kofman, S.; Lee, Y.H.; Xiao, Y.; Siqueira, W.L. Proteome and Peptidome of Human Acquired Enamel Pellicle on Deciduous Teeth. Int. J. Mol. Sci. 2013, 14, 920-934. https://doi.org/10.3390/ijms14010920
Zimmerman JN, Custodio W, Hatibovic-Kofman S, Lee YH, Xiao Y, Siqueira WL. Proteome and Peptidome of Human Acquired Enamel Pellicle on Deciduous Teeth. International Journal of Molecular Sciences. 2013; 14(1):920-934. https://doi.org/10.3390/ijms14010920
Chicago/Turabian StyleZimmerman, Jason N., William Custodio, Sahza Hatibovic-Kofman, Young Ho Lee, Yizhi Xiao, and Walter L. Siqueira. 2013. "Proteome and Peptidome of Human Acquired Enamel Pellicle on Deciduous Teeth" International Journal of Molecular Sciences 14, no. 1: 920-934. https://doi.org/10.3390/ijms14010920
APA StyleZimmerman, J. N., Custodio, W., Hatibovic-Kofman, S., Lee, Y. H., Xiao, Y., & Siqueira, W. L. (2013). Proteome and Peptidome of Human Acquired Enamel Pellicle on Deciduous Teeth. International Journal of Molecular Sciences, 14(1), 920-934. https://doi.org/10.3390/ijms14010920