Up-Regulation of mRNA Ventricular PRNP Prion Protein Gene Expression in Air Pollution Highly Exposed Young Urbanites: Endoplasmic Reticulum Stress, Glucose Regulated Protein 78, and Nanosized Particles

Abstract
:1. Introduction
2. Results
2.1. Study City and Air Quality
Air Quality Data
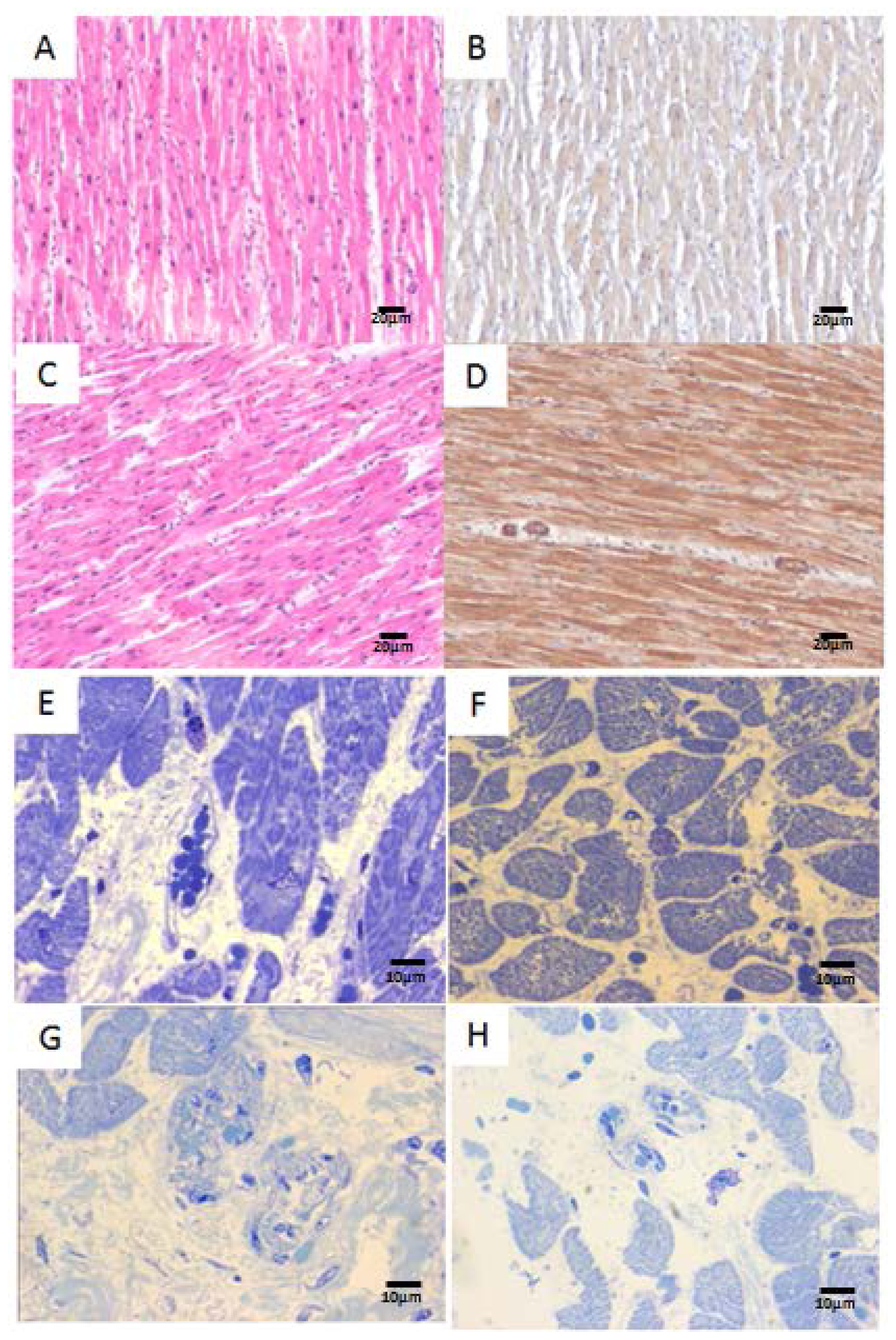
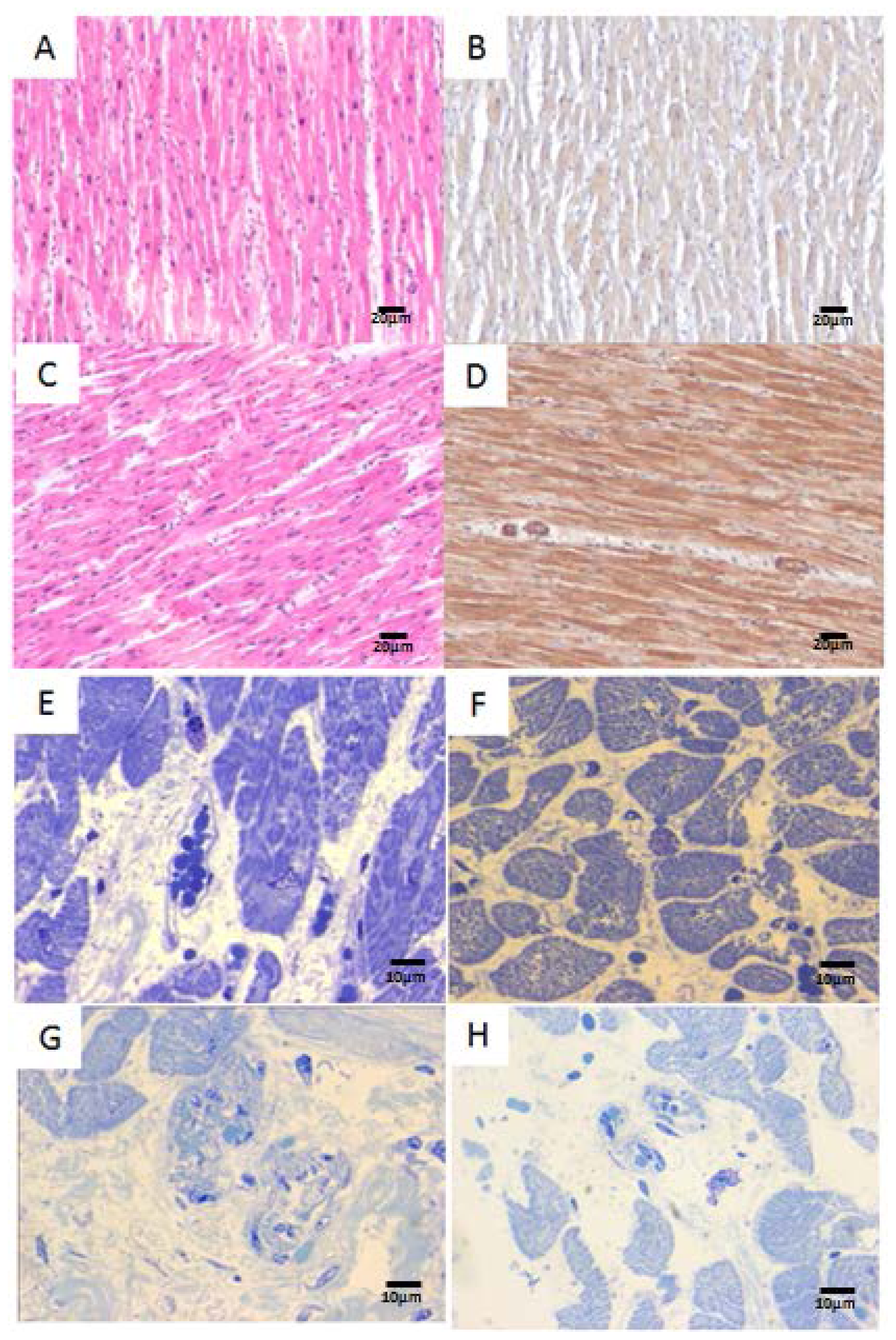
2.2. Human Heart Histopathology
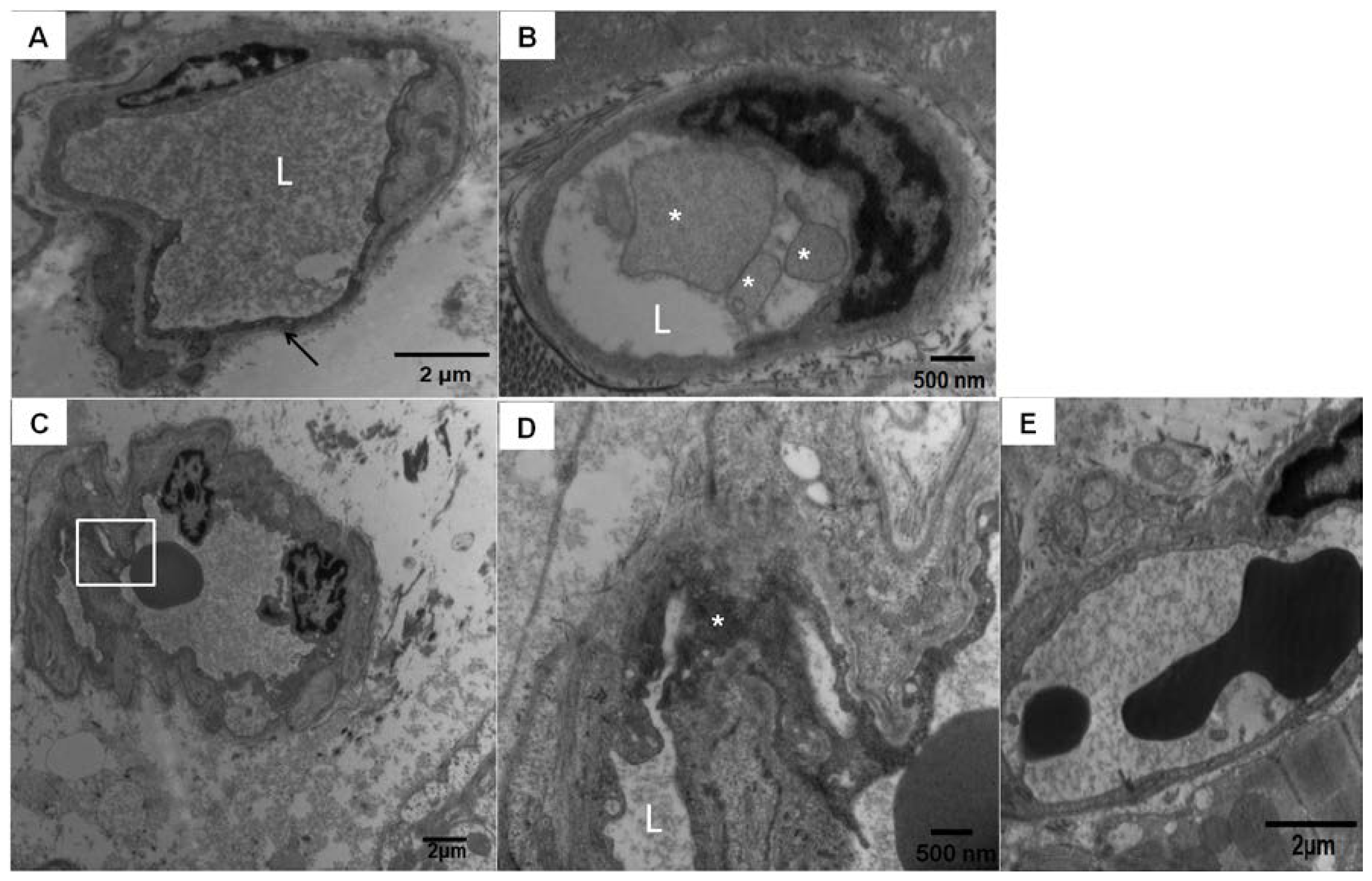
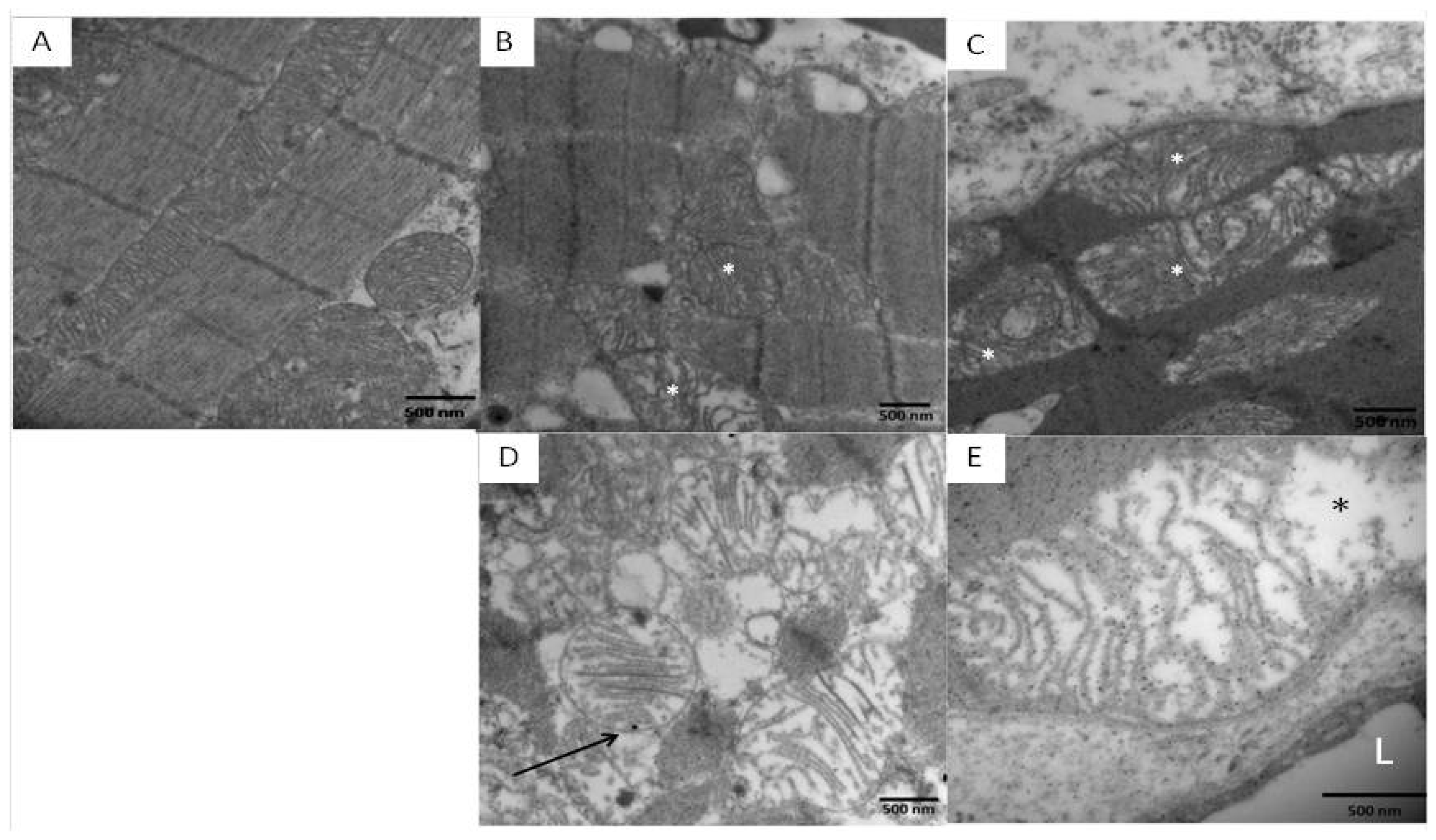
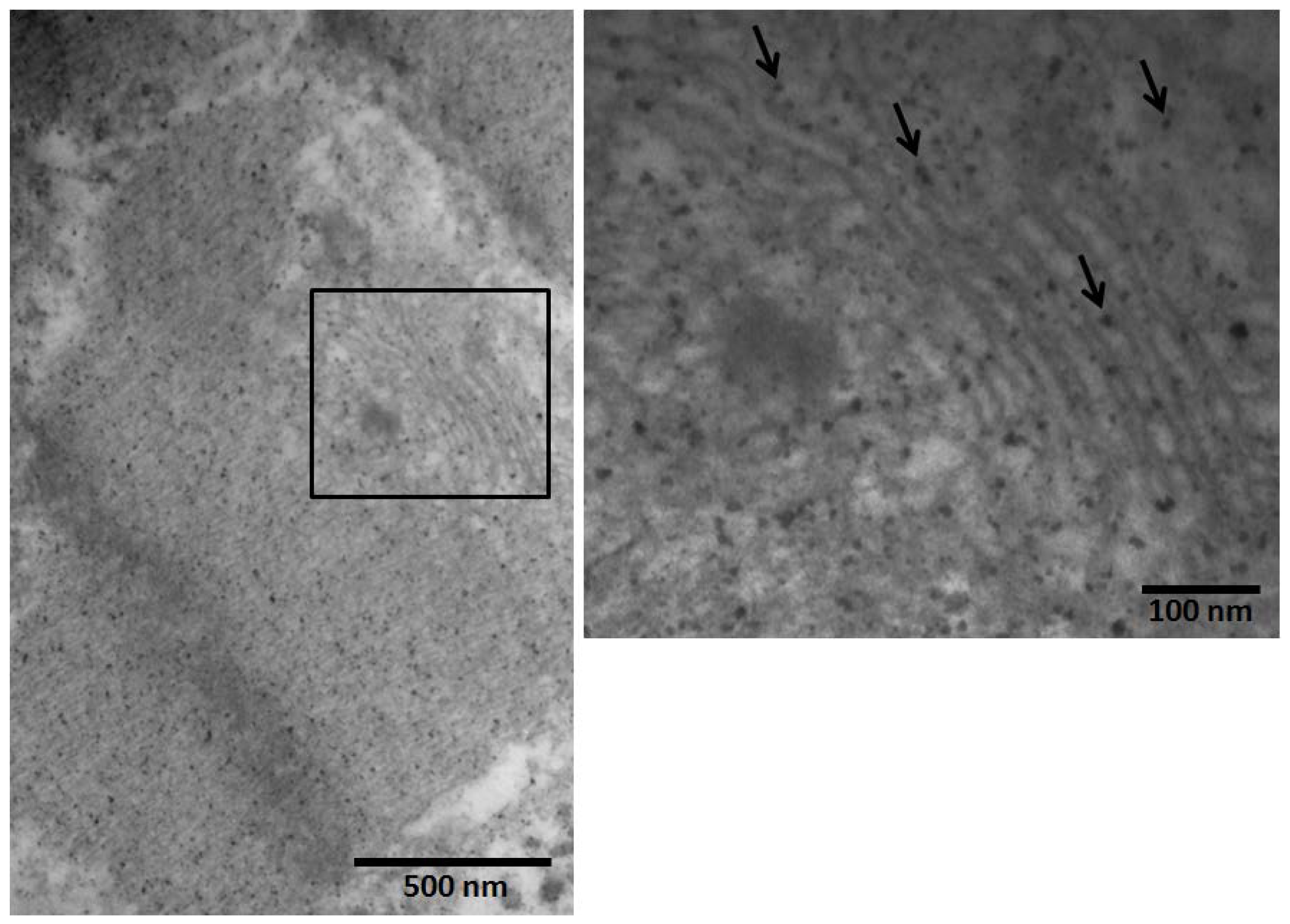
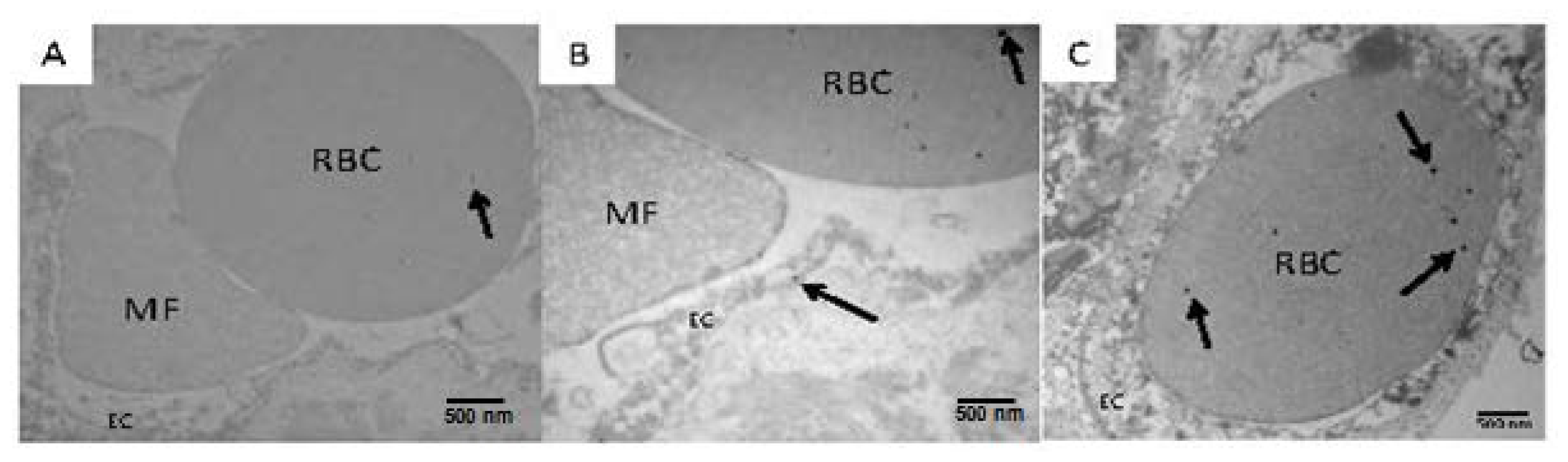
2.3. Electron Microscopic Findings in Mexico City Dogs and Teens
2.4. Real-Time PCR Analysis of Target Genes
3. Discussion
4. Experimental Section
4.1. Heart Samples
4.2. Light and Electron Microscopy
4.3. Examination of Heart Samples by Transmission Electron Microscopy (TEM)
4.4. Estimation of mRNA Abundance by RT-PCR
4.5. Statistics
4.6. Air Pollution Data
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Brook, R.D.; Rajagopalan, S.; Pope, C.A., 3rd; Brook, J.R.; Bhatnagar, A.; Diez-Roux, A.V.; Holguin, F.; Hong, Y.; Luepker, R.V.; Mittleman, M.A.; et al. Particulate matter air pollution and cardiovascular disease: An update to the scientific statement from the American Heart Association. Circulation 2010, 121, 2331–2378. [Google Scholar]
- Franchini, M.; Mannucci, P.M. Thrombogenicity and cardiovascular effects of ambient air pollution. Blood 2011, 118, 2405–2412. [Google Scholar]
- Martinelli, N.; Olivieri, O.; Girelli, D. Air particulate matter and cardiovascular disease: A narrative review. Eur. J. Intern. Med 2013, 24, 295–302. [Google Scholar]
- Shields, K.N.; Cavallari, J.M.; Hunt, M.J.; Lazo, M.; Molina, M.; Molina, L.; Holguin, F. Traffic-related air pollution exposures and changes in heart rate variability in Mexico City: A panel study. Environ. Health 2013, 12, 7. [Google Scholar]
- Chen, H.; Goldberg, M.S.; Burnett, R.T.; Jerrett, M.; Wheeler, A.J.; Villeneuve, P.J. Long-term exposure to traffic-related air pollution and cardiovascular mortality. Epidemiology 2013, 24, 35–43. [Google Scholar]
- Bravo-Alvarez, H.R.; Torres-Jardón, R.J. Air Pollution Levels and Trends in the Mexico City Metropolitan Area. In Urban Air Pollution and Forests: Resources at Risk in the Mexico City Air Basin Ecological Studies; Fenn, M., Bauer, L., Hernández, T., Eds.; Springer-Verlag: New York, NY, USA, 2002; Volume 156, pp. 121–159. [Google Scholar]
- Molina, L.T.; Madronich, S.; Gaffney, J.S.; Apel, E.; de Foy, B.; Fast, J.; Ferrare, R.; Herndon, S.; Jimenez, J.L.; Lamb, B.; et al. An overview of the MILAGRO 2006 Campaign: Mexico City emissions and their transport and transformation. Atmos. Chem. Phys 2010, 10, 8697–8760. [Google Scholar] [Green Version]
- Bravo-Alvarez, H.R.; Sosa-Echeverria, R.; Sanchez-Alvarez, P.; Krupa, S. Air quality standards for particulate matter (PM) at high altitude cities. Environ. Pollut 2013, 173, 255–262. [Google Scholar]
- Rosas-Pérez, I.; Serrano, J.; Alfaro-Moreno, E.; Baumgardner, D.; Garcia-Cuellar, C.; Martin del Campo, J.M.; Raga, G.B.; Castillejos, M.; Colin, R.D.; Osornio-Vargas, A.R. Relations between PM10 composition and cell toxicity: A multivariate and graphical approach. Chemosphere 2007, 67, 1218–1228. [Google Scholar]
- Querol, X.; Pey, J.; Minguillón, M.C.; Pérez, N.; Alastuey, A.; Viana, M.; Moreno, T.; Bernabe, R.M.; Blanco, S.; Cardenas, B.; et al. PM speciation and sources in Mexico during the MILAGRO-2006 Campaign. Atmos. Chem. Phys 2008, 8, 111–121. [Google Scholar]
- Villarreal-Calderon, R.; Reed, W.; Keefe, S.; Herritt, L.; Brooks, D.; Torres-Jardón, R.; Calderón-Garcidueñas, L. Urban air pollution produces up-regulation of myocardial inflammatory genes and dark chocolate provides cardioprotection. Exp. Toxicol. Pathol 2012, 64, 297–306. [Google Scholar]
- Osornio-Vargas, A.R.; Bonner, J.C.; Alfaro-Moreno, E.; Martinez, L.; Garcia-Cuellar, C.; Ponce-de-Leon-Rosales, S.; Miranda, J.; Rosas, I. Proinflammatory and cytotoxic effects of Mexico City air pollution particulate matter in vitro are dependent on particle size and composition. Environ. Health Perspect 2003, 111, 1289–1293. [Google Scholar]
- Villarreal-Calderon, R.; Dale, G.; Delgado-Chávez, R.; Torres-Jardón, R.; Zhu, H.; Herritt, L.; Gónzalez-Maciel, A.; Reynoso-Robles, R.; Yuan, Y.; Wang, J.; et al. Intracity differences in cardiac expression of inflammatory genes and inflammasomes in young urbanites: A pilot study. J. Toxicol. Pathol 2012, 25, 163–173. [Google Scholar]
- McKinley, M.P.; Taraboulus, A.; Kenaga, L.; Serban, D.; Stieber, A.; DeArmond, S.J.; Prusiner, S.B.; Gonatas, N. Ultrastructural localization of scrapie prion proteins in cytoplasmic vesicles of infected cultured cells. Lab. Investig 1991, 65, 622–630. [Google Scholar]
- Peralta, O.A.; Eyestone, W.H. Quantitative and qualitative analysis of cellular prion protein (PrPC) expression in bovine somatic tissues. Prion 2009, 3, 161–170. [Google Scholar]
- Milhavet, O.; Lehmann, S. Oxidative stress and the prion protein in transmissible spongiform encephalopathies. Brain Res. Brain Res. Rev 2002, 38, 328–339. [Google Scholar]
- Brown, D.R.; Schmidt, B.; Kretzschmar, H.A. Effects of oxidative stress on prion protein expression in PC12 cells. Int. J. Dev. Neurosci 1997, 15, 961–972. [Google Scholar]
- Bertuchi, F.R.; Bourgeon, D.M.; Landemberger, M.C.; Martins, V.R.; Cerchiaro, G. PrPC displays an essential protective role from oxidative stress in an astrocyte cell line derived from PrPC knockout mice. Biochem. Biophys. Res. Commun 2012, 418, 27–32. [Google Scholar]
- Sinclair, L.; Lewis, V.; Collins, S.J.; Haigh, C.L. Chronic prion infection depletes superoxide dismutase-2 through promoting exposure to cytosolic caspases. Dis. Model. Mech 2013, in press. [Google Scholar]
- Viles, J.H.; Cohen, F.E.; Prusiner, S.B.; Goodin, D.B.; Wright, P.E.; Dyson, H.J. Copper binding to the prion protein: Structural implications of four identical cooperative binding sites. Proc. Natl. Acad. Sci. USA 1999, 96, 2042–2047. [Google Scholar]
- Alfaidy, N.; Chauvet, S.; Donadio-Andrei, S.; Salomon, A.; Saoudi, Y.; Richaud, P.; Aude-garcia, C.; Hoffman, P.; Andrieux, A.; Moulis, J.M.; et al. Prion protein expression and functional importance in developmental angiogenesis: Role in oxidative stress and copper homeostasis. Antioxid. Redox Signal 2013, 18, 400–411. [Google Scholar]
- Westergard, L.; Christensen, H.M.; Harris, D.A. The cellular prion protein (PrPC): Its physiological function and role in disease. Biochem. Biophys. Acta 2007, 1772, 629–644. [Google Scholar]
- Hu, W.; Rosenberg, R.N.; Stüve, O. Prion proteins: A biological role beyond prion diseases. Acta Neurol. Scand 2007, 116, 75–82. [Google Scholar]
- Nico, P.B.; Lobão-Soares, B.; Landemberger, M.C.; Marques, W., Jr.; Tasca, C.I.; de mello, C.F.; Walz, R.; Carlotti, C.G., Jr; Brentani, R.R.; Sakamoto, A.C.; et al. Impaired exercise capacity, but unaltered mitochondrial respiration in skeletal or cardiac muscle of mice lacking cellular prion protein. Neurosci. Lett 2005, 388, 21–26. [Google Scholar]
- Soprana, C.; Souza, C.L.; Debbas, V.; Laurindo, M.F.R. Cellular prion protein (PrPC) and superoxide dismutase (SOD) in vascular cells under oxidative stress. Exp. Toxicol. Pathol 2011, 63, 229–236. [Google Scholar]
- Schneider, B.; Mutel, V.; Pietri, M.; Ermonval, M.; Mouillet-Richard, S.; Kellermann, O. NADPH oxidase and extracellular regulated kinases are targets of prion protein signaling in neuronal and nonneuronal cells. Proc. Natl. Acad. Sci. USA 2003, 100, 13326–13331. [Google Scholar]
- Smith, J.D.; Moylan, J.S.; Hardin, B.J.; Chambers, M.A.; Estus, S.; Telling, G.C.; Reid, M.B. Prion protein expression and functional importance in skeletal muscle. Antioxid. Redox Signal 2011, 15, 2465–2475. [Google Scholar]
- Whatley, S.A.; Powell, J.F.; Politopoulou, G.; Campbell, I.C.; Brammer, M.J.; Percy, N.S. Regulation of intracellular free calcium levels by the cellular prion protein. Neuroreport 1995, 6, 2333–2337. [Google Scholar]
- Lazzari, C.; Peggion, C.; Stella, R.; Massimino, M.L.; Lim, D.; Bertoli, A.; Sorgato, M.C. Cellular prion protein is implicated in the regulation of local Ca2+ movements in cerebellar granule neurons. J. Neurochem 2011, 116, 881–890. [Google Scholar]
- Beraldo, F.H.; Arantes, C.P.; Santos, T.G.; Queiroz, N.G.; Young, K.; Rvlett, R.J.; Markus, R.P.; Prado, M.A.; Martins, V.R. Role of alpha 7 nicotinic acetylycholine receptor in calcium signaling induced by prion protein interaction with stress-inducible protein 1. J. Biol. Chem 2010, 285, 36542–36550. [Google Scholar]
- Déry, M.A.; Jodoin, J.; Ursini-Siegel, J.; Aleynikova, O.; Ferrario, C.; Hassan, S.; Basik, M.; Leblanc, A.C. Endoplasmic reticulum stress induces PRNP prion protein gene expression in breast cancer. Breast Cancer Res 2013, 15, R1–R22. [Google Scholar]
- Glembotski, C.C. Endoplasmic reticulum stress in the heart. Circ. Res 2007, 101, 975–984. [Google Scholar]
- Groenendyk, J.; Sreenivasaiah, P.K.; Kim, D.H.; Agellon, L.B.; Michalak, M. Biology of endoplasmic reticulum stress in the heart. Circ. Res 2010, 107, 1185–1197. [Google Scholar]
- Doroudgar, S.; Glembotski, C.C. New concepts of endoplasmic reticulum function in the heart: Programmed to conserve. J. Mol. Cell. Cardiol 2013, 55, 85–91. [Google Scholar]
- Prins, D.; Michalak, M. Endoplasmic reticulum proteins in cardiac development and dysfunction. Can. J. Physiol. Pharmacol 2009, 87, 419–425. [Google Scholar]
- Brown, M.K.; Naidoo, N. The endoplasmic reticulum stress response in aging and age-related diseases. Front. Physiol 2012, 3, 263. [Google Scholar]
- Gorbatyuk, M.S.; Gorbatyuk, O.S. The molecular chaperone GRP78/BiP as a therapeutic target for neurodegenerative disorders: A mini review. J. Genet. Syndr. Gene Ther 2013, 4, 128. [Google Scholar]
- Christen, V.; Fent, K. Silica nanoparticles and silver-doped silica nanoparticles induce endoplasmatic reticulum stress response and alter cytochrome P4501A activity. Chemosphere 2012, 87, 423–434. [Google Scholar]
- Glembotski, C.C. Roles for ATF6 and the sarco/endoplasmic reticulum protein quality control system in the heart. J. Mol. Cell. Cardiol 2013, in press. [Google Scholar]
- Departamento del Distrito Federal. Available online: http://www.sma.df.gob.mx/simat2/ (accessed on 14 September 2012).
- Calderón-Segura, M.E.; Gómez-Arroyo, S.; Villalobos-Pietrini, R.; Butterworth, F.M.; Amador-Munoz, O. The effects of seasonal weather on the genotoxicity, cytokinetic properties, cytotoxicity and organochemical content of extracts of airborne particulates in Mexico City. Mutat. Res 2004, 558, 7–17. [Google Scholar]
- Dzepina, K.; Arey, J.; Marr, L.; Worsnop, D.R.; Salcedo, D.; Zhang, Q.; Onasch, T.B.; Molina, L.T.; Molina, M.J.; Jimenez, J.L. Detection of particle-phase polycyclic aromatic hydrocarbons in Mexico City using an aerosol mass spectrometer. Int. J. Mass Spectrom 2007, 263, 152–170. [Google Scholar]
- Estrada-Garcia, T.; Cerna, J.F.; Thompson, M.R.; López-Saucedo, C. Fecal contamination and enterotoxigenic Escherichia coli in street-vended chili sauces in Mexico and its public relevance. Epidemiol. Infect 2002, 129, 223–226. [Google Scholar]
- García-Pérez, C.; Schneider, T.G.; Hajnoczky, G.; Csordas, G. Alignment of sarcoplasmic reticulum-mitochondrial junctions with mitochondrial contact points. Am. J. Physiol. Heart Circ. Physiol 2011, 301, H1907–H1915. [Google Scholar]
- Mitsios, N.; Saka, M.; Krupinski, J.; Pennucci, R.; Sanfeliu, C.; Miguel-Turu, M.; Gaffney, J.; Kumar, P.; Kumar, S.; Sullivan, M.; et al. Cellular prion protein is increased in the plasma and peri-infacted brain tissue after acute stroke. J. Neurosci. Res 2007, 85, 602–611. [Google Scholar]
- Anantharam, V.; Kanthasamy, A.; Choi, C.J.; Martin, D.P.; Latchoumycandane, C.; Richt, J.A.; Kanthasamy, A.G. Opposing roles of prion protein in oxidative stress-and ER stress-induced apoptotic signaling. Free Radic. Biol. Med 2008, 45, 1530–1541. [Google Scholar]
- Singh, N.; Singh, A.; Das, D.; Mohan, M.L. Redox control of prion and disease pathogenesis. Antioxid. Redox Signal 2010, 12, 1271–1294. [Google Scholar]
- Choi, C.J.; Anantharam, V.; Saetveit, N.J.; Houk, R.S.; Kanthasamy, A.; Kanthasamy, A.G. Normal cellular prion protein protects against manganese-induced oxidative stress and apoptotic cell death. Toxicol. Sci 2007, 98, 495–509. [Google Scholar]
- Park, Y.G.; Jeong, J.K.; Lee, J.H.; Lee, Y.J.; Seol, J.W.; Kim, S.J.; Hur, T.Y.; Jung, Y.H.; Kang, S.J.; Park, S.Y. Lactoferrin protects against prion-protein-induced cell death in neuronal cells by preventing mitochondrial dysfunction. Int. J. Mol. Med 2013, 31, 325–330. [Google Scholar]
- Singh, N.; Das, D.; Singh, A.; Mohan, M.L. Prion protein and metal interaction: Physiological and pathological implications. Curr. Issues Mol. Biol 2010, 12, 99–107. [Google Scholar]
- Choi, C.J.; Anantharam, V.; Martin, D.P.; Nicholson, E.M.; Richt, J.A.; Kanthasamy, A.; Kanthasamy, A.G. Manganese upregulates cellular prion protein and contributes to altered stabilization and proteolysis: Relevance to role of metals in pathogenesis of prion disease. Toxicol. Sci 2010, 115, 535–546. [Google Scholar]
- Holland, M.R.; Gibson, A.A.; Kirshner, C.A.; Hicks, D.; Ludomirsky, A.; Singh, G.K. Intrinsic myoarchitectural differences between the left and right ventricles of fetal human hearts: An ultrasonic backscatter feasibility study. J. Am. Soc. Echocardiogr 2009, 22, 170–176. [Google Scholar]
- Tverskaya, M.S.; Sukhoparova, V.V.; Karpova, V.V.; Raksha, A.P.; Kadyrova, M.K.; Abdulkerimova, N.Z.; Bobrova, N.A. Pathomorphology of myocardial circulation: Comparative study in increased left or right ventricle overload. Bull. Exp. Biol. Med 2008, 145, 377–381. [Google Scholar]
- Schulz, R.; Heusch, G. Tumor necrosis factor-alpha and its receptors 1 and 2. Circulation 2009, 119, 1355–1357. [Google Scholar]
- Schulz, R. TNF α in myocardial ischemia/reperfusion: Damage vs. protection. J. Mol. Cell. Cardiol 2008, 45, 712–714. [Google Scholar]
- Chen, Y.; Ke, Q.; Yang, Y.; Rana, J.S.; Tang, J.; Morgan, J.P.; Xiao, Y.F. Cardiomyocytes overexpressing TNF-alpha attract migration of embryonic stem cells via activation of p38 and c-Jun amino-terminal kinase. Fed. Am. Soc. Exp. Biol. J 2003, 17, 2231–2239. [Google Scholar]
- Bao, C.; Guo, J.; Lin, G.; Hu, Z. TNFR gene-modified mesenchymal stem cells attenuate inflammation and cardiac dysfunction following MI. Scand. Cardiovasc. J 2008, 42, 56–62. [Google Scholar]
- Sovolyova, N.; Healy, S.; Samali, A.; Logue, S.E. Stressed to death-mechanisms of ER stress-induced cell death. Biol. Chem 2013, in press. [Google Scholar]
- McAlpine, C.S.; Werstuck, G.H. The development and progression of atherosclerosis: Evidence supporting a role for endoplasmic reticulum (ER) stress signaling. Cardiovasc. Hematol. Disord. Drug Targets 2013, 13, 158–164. [Google Scholar]
- Abdelhalim, M.A. Gold nanoparticles administration induces disarray of heart muscle, hemorrhagic, chronic inflammatory cells infiltrated by small lymphocytes, cytoplasmic vacuolization and congested and dilated blood vessels. Lipids Health Dis 2011, 10, 233. [Google Scholar]
- Helfenstein, M.; Miragoli, M.; Rohr, S.; Müller, L.; Wick, P.; Mohr, M.; Gehr, P.; Rothen-Rutishauser, B. Effects of combustion-derived ultrafine particles and manufactured nanoparticles on heart cells in vitro. Toxicology 2008, 253, 70–78. [Google Scholar]
- Okayama, Y.; Kuwahara, M.; Suzuki, A.K.; Tsubone, H. Role of reactive oxygen species on diesel exhaust particle-induced cytotoxicity in rat cardiac myocytes. J. Toxicol. Environ. Health 2006, 69, 1699–1710. [Google Scholar]
- Guo, D.; Bi, H.; Liu, B.; Wu, Q.; Wang, D.; Cui, Y. Reactive oxygen species-induced cytotoxic effects of zinc oxide nanoparticles in rat retinal ganglion cells. Toxicol. In Vitro 2013, 27, 731–738. [Google Scholar]
- Ariano, P.; Zamburlin, P.; Gilardino, A.; Mortera, R.; Onida, B.; Tomatis, M.; Ghiazza, M.; Fubini, B.; Lovisolo, D. Interaction of spherical nanoparticles with neuronal cells: Size-dependent toxicity and perturbation of calcium homeostasis. Small 2011, 6, 766–774. [Google Scholar]
- Tsai, Y.Y.; Huang, Y.H.; Chao, Y.L.; Hu, K.Y.; Chin, L.T.; Chou, S.H.; Hour, A.L.; Yao, Y.D.; Tu, C-S.; Liang, Y.J.; et al. Identification of the nanogold particle induced endoplasmic reticulum stress by Omic techniques and systems biology analysis. ACS Nano 2011, 12, 9354–9369. [Google Scholar]
- Miragoli, M.; Novak, P.; Ruenraroengsak, P.; Shevchuck, A.I.; Korchev, Y.E.; Lab, M.J.; Tetley, T.D.; Gorelik, J. Functional interaction between charged nanoparticles and cardiac tissue: A new paradigm for cardiac arrhythmia? Nanomedicine 2013, 8, 725–737. [Google Scholar]
- Calderón-Garcidueñas, L.; Hazucha, M.J.; Herbst, M.C.; Reed, W.; Cascio, W.E. Exposure to Urban Air Pollution is Associated with Decreased Vagal Tone in Clinically Healthy Children. In Fibers and Nanoparticles in Lung and Cardiovascular Disease, Proceedings of 8th International Meeting on Mechanisms of Action of Inhaled Particles, Research Triangle Park: NC, USA, 2005; 4 June 2005. [Google Scholar]
- Papanicolau, K.N.; Phillippo, M.M.; Walsh, K. Mitofusins and the mitochondrial permeability transition: The potential downside of mitochondrial fusion. Am. J. Physiol. Heart Circ. Physiol 2012, 303, H243–H255. [Google Scholar]
- Papanicolau, K.N.; Khairallah, R.J.; Ngoh, G.A.; Chikando, A.; Luptak, I.; O’Shea, K.M.; Riley, D.D.; Lugus, J.J.; Colucci, W.S.; Lederer, W.J.; et al. Mitofusin-2 maintains mitochondrial structure and contributes to stress-induced permeability transition in cardiac myocytes. Mol. Cell. Biol 2011, 31, 1309–1328. [Google Scholar]
- Chen, Y.; Csordas, G.; Jowdy, C.; Schneider, T.G.; Csordas, N.; Wang, W.; Liu, Y.; Kohlhaas, M.; Meiser, M.; bergem, S.; et al. Mitofusin 2-containing mitochondrial-reticular microdomains direct rapid cardiomyocyte bioenergetics responses via interorganelle Ca2+ crosstalk. Circ. Res 2012, 111, 863–875. [Google Scholar]
- Levick, S.P.; Murray, D.B.; Janicki, J.S.; Brower, G.L. Sympathetic nervous system modulation of inflammation and remodeling in the hypertensive heart. Hypertension 2010, 55, 270–276. [Google Scholar]
- Eszlári, E.; Czóbel, M.; Molnar, G.; Kaszaki, J.; Nagy, S.; Boros, M. Modulation of cardiac contractility through endothelin-1 release and myocardial mast cell degranulation. Acta Physiol. Hung 2008, 95, 267–285. [Google Scholar]
- Martinon, F.; Mayor, A.; Tschopp, J. The inflammasomes: Guardians of the body. Annu. Rev. Immunol 2009, 27, 229–265. [Google Scholar]
- Martinon, F. Signaling by ROS drives inflammasome activation. Eur. J. Immunol 2010, 40, 616–619. [Google Scholar]
- Yin, Y.; Yan, Y.; Jiang, X.; Mai, J.; Chen, N.C.; Wang, H.; Yang, X.F. Inflammasomes are differentially expressed in cardiovascular and other tissues. Int. J. Immunopathol. Pharmacol 2009, 22, 311–322. [Google Scholar]
- Golomb, E.; Nyska, A.; Schwalb, H. Occult cardiotoxicity-toxic effects on cardiac ischemic tolerance. Toxicol. Pathol 2009, 37, 572–593. [Google Scholar]
- Brandenberger, C.; Clift, M.J.D.; Vanhecke, D.; Mühlfeld, C.; Stone, V.; Gehr, P.; Rothen-Rutishauser, B. Intracellular imaging of nanoparticles: Is it an elemental mistake to believe what you see? Part. Fibre Toxicol 2010, 7, 15–20. [Google Scholar]
- Livak, K.J.; Schmittgen, D.T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔC(T) method. Methods 2001, 25, 402–408. [Google Scholar]
- The Green Book Nonattainment Areas for Criteria Pollutants. Available online: http://www.epa.gov/airquality/greenbk/ (accessed on 28 November 2013).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Pedregal | Xalostoc | ||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| 1997 | 21.6 | 16.6 | 71.3 | 34.1 |
| 1998 | 29.3 | 16.8 | 64.9 | 25.4 |
| 1999 | 24.4 | 9.2 | 71.0 | 26.6 |
| 2000 | 24.7 | 11.3 | 54.8 | 25.3 |
| 2001 | 23.6 | 10.1 | 41.1 | 17.2 |
| 2002 | 23.1 | 9.7 | 38.0 | 13.7 |
| 2003 | 23.4 | 11.3 | 41.8 | 14.4 |
| 2004 | 18.4 | 9.4 | 35.5 | 14.7 |
| 2005 | 20.9 | 11.5 | 30.4 | 17.1 |
| 2006 | 17.8 | 8.4 | 29.8 | 15.6 |
| 2007 | 16.2 | 8.5 | 25.3 | 11.3 |
| 2008 | 18.0 | 8.3 | 26.3 | 10.0 |
| 2009 | 18.4 | 8.7 | 26.4 | 10.7 |
| 2010 | 14.4 | 7.4 | 24.9 | 13.2 |
| 2011 | 16.7 | 8.3 | 24.7 | 11.5 |
| 2012 | 17.0 | 7.5 | 25.9 | 11.7 |
| RT-PCR | PrPC left ventricle | PrPC right ventricle | BiP left ventricle | BiP right ventricle |
|---|---|---|---|---|
| Mean | 6.852 | 0.7209 | 3.410 | 2.465 |
| Std. deviation | 7.681 | 0.7207 | 2.254 | 1.794 |
| p value exposed vs. controls | 0.0062 | 0.1360 | 0.00004 | 0.0007 |
| p value left vs. right ventricles in exposed subjects | <0.0001 | 0.1189 | ||
| Pearson and Kendall’s Stats | Pearson’s r PrPC_L & BiP_L | Pearson’s r PrPC_R & BiP_R | Kendall’s tau PrPC_L & BiP_L | Kandall’s tau PrPC_R & BiP_R |
|---|---|---|---|---|
| r | 0.6645 | 0.2409 | 0.6230 | 0.1530 |
| p value | 5.435 × 10−4 | 0.3063 | 3.334 × 10−5 | 0.3465 |
| Two-sample test | ||||
| PrPC_L | PrPC_R | BiP_L | BiP_R | |
| p value | 0.1272 | 0.9024 | 0.4563 | 0.6817 |
| Wilcoxon rank sum test | ||||
| PrPC_L | PrPC_R | BiP_L | BiP_R | |
| p value | 0.3050 | 0.8366 | 0.2973 | 0.9864 |
| PrPC LV North | PrPC LV South | PrPC RV North | PrPC RV South | BiP LV North | BiP LV South | BiP RV North | BiP RV South | |
|---|---|---|---|---|---|---|---|---|
| Mean | 8.548 | 9.292 | 0.9723 | 0.5889 | 4.905 | 3.317 | 3.151 | 1.542 |
| Std. deviation | 9.151 | 11.04 | 1.149 | 0.6262 | 2.720 | 1.355 | 2.333 | 0.8514 |
| p value | 0.5273 | 0.9273 | 0.4970 | 0.3825 | ||||
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Villarreal-Calderon, R.; Franco-Lira, M.; González-Maciel, A.; Reynoso-Robles, R.; Harritt, L.; Pérez-Guillé, B.; Ferreira-Azevedo, L.; Drecktrah, D.; Zhu, H.; Sun, Q.; et al. Up-Regulation of mRNA Ventricular PRNP Prion Protein Gene Expression in Air Pollution Highly Exposed Young Urbanites: Endoplasmic Reticulum Stress, Glucose Regulated Protein 78, and Nanosized Particles. Int. J. Mol. Sci. 2013, 14, 23471-23491. https://doi.org/10.3390/ijms141223471
Villarreal-Calderon R, Franco-Lira M, González-Maciel A, Reynoso-Robles R, Harritt L, Pérez-Guillé B, Ferreira-Azevedo L, Drecktrah D, Zhu H, Sun Q, et al. Up-Regulation of mRNA Ventricular PRNP Prion Protein Gene Expression in Air Pollution Highly Exposed Young Urbanites: Endoplasmic Reticulum Stress, Glucose Regulated Protein 78, and Nanosized Particles. International Journal of Molecular Sciences. 2013; 14(12):23471-23491. https://doi.org/10.3390/ijms141223471
Chicago/Turabian StyleVillarreal-Calderon, Rodolfo, Maricela Franco-Lira, Angélica González-Maciel, Rafael Reynoso-Robles, Lou Harritt, Beatriz Pérez-Guillé, Lara Ferreira-Azevedo, Dan Drecktrah, Hongtu Zhu, Qiang Sun, and et al. 2013. "Up-Regulation of mRNA Ventricular PRNP Prion Protein Gene Expression in Air Pollution Highly Exposed Young Urbanites: Endoplasmic Reticulum Stress, Glucose Regulated Protein 78, and Nanosized Particles" International Journal of Molecular Sciences 14, no. 12: 23471-23491. https://doi.org/10.3390/ijms141223471
APA StyleVillarreal-Calderon, R., Franco-Lira, M., González-Maciel, A., Reynoso-Robles, R., Harritt, L., Pérez-Guillé, B., Ferreira-Azevedo, L., Drecktrah, D., Zhu, H., Sun, Q., Torres-Jardón, R., Aragón-Flores, M., Calderón-Garcidueñas, A., Diaz, P., & Calderón-Garcidueñas, L. (2013). Up-Regulation of mRNA Ventricular PRNP Prion Protein Gene Expression in Air Pollution Highly Exposed Young Urbanites: Endoplasmic Reticulum Stress, Glucose Regulated Protein 78, and Nanosized Particles. International Journal of Molecular Sciences, 14(12), 23471-23491. https://doi.org/10.3390/ijms141223471