Optimization of the Preparation of Fish Protein Anti-Obesity Hydrolysates Using Response Surface Methodology

Abstract
:1. Introduction
2. Results and Discussion
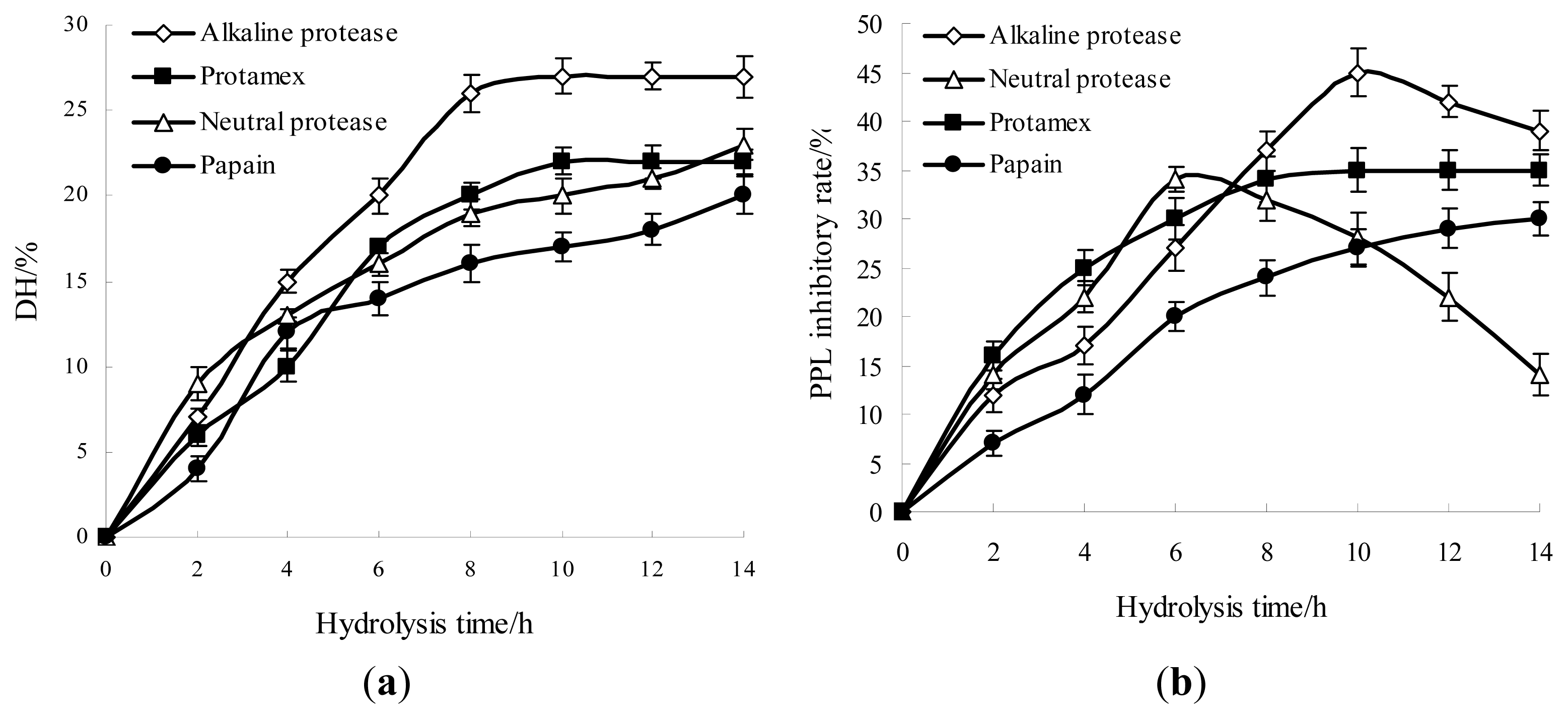
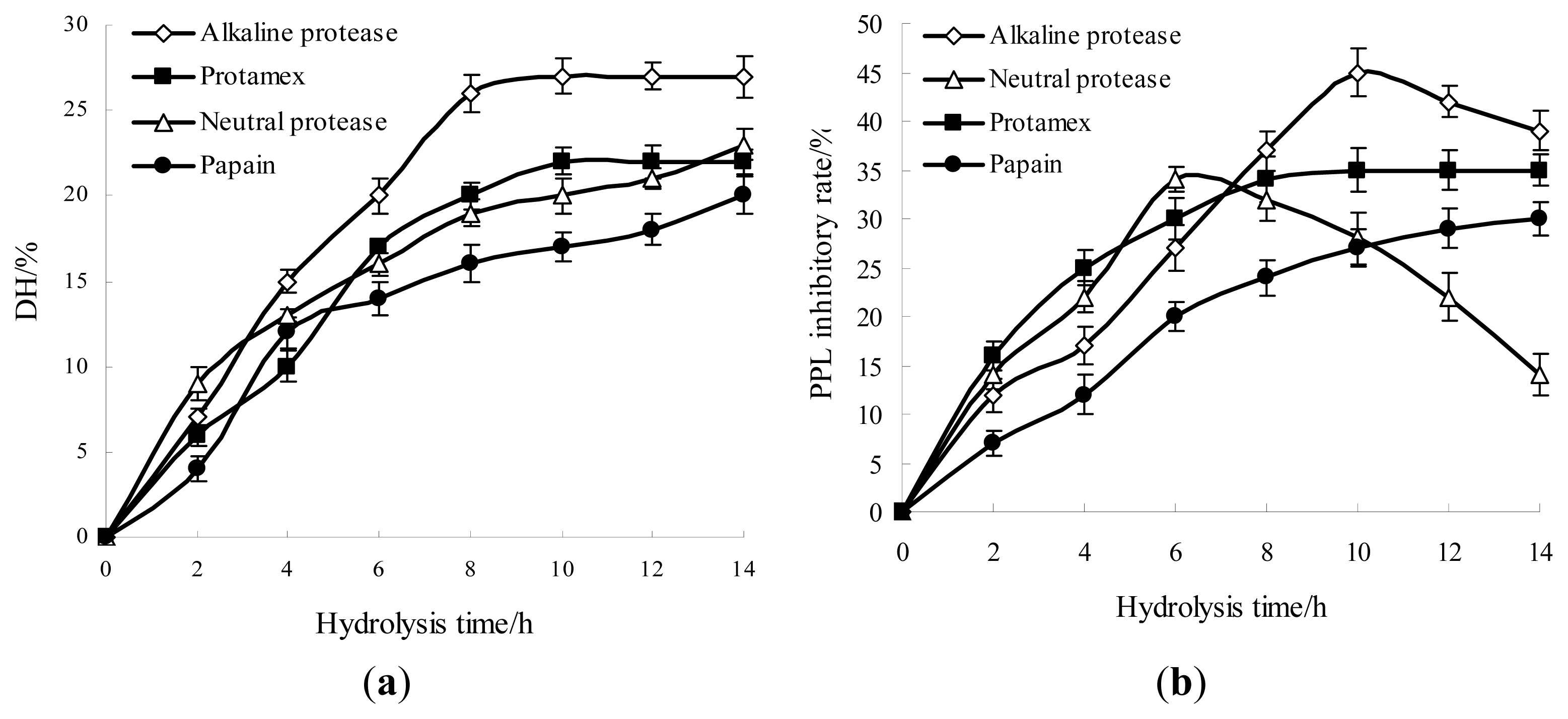
2.1. Effects of Different Proteases Hydrolysis for Producing Anti-Obesity Hydrolysates
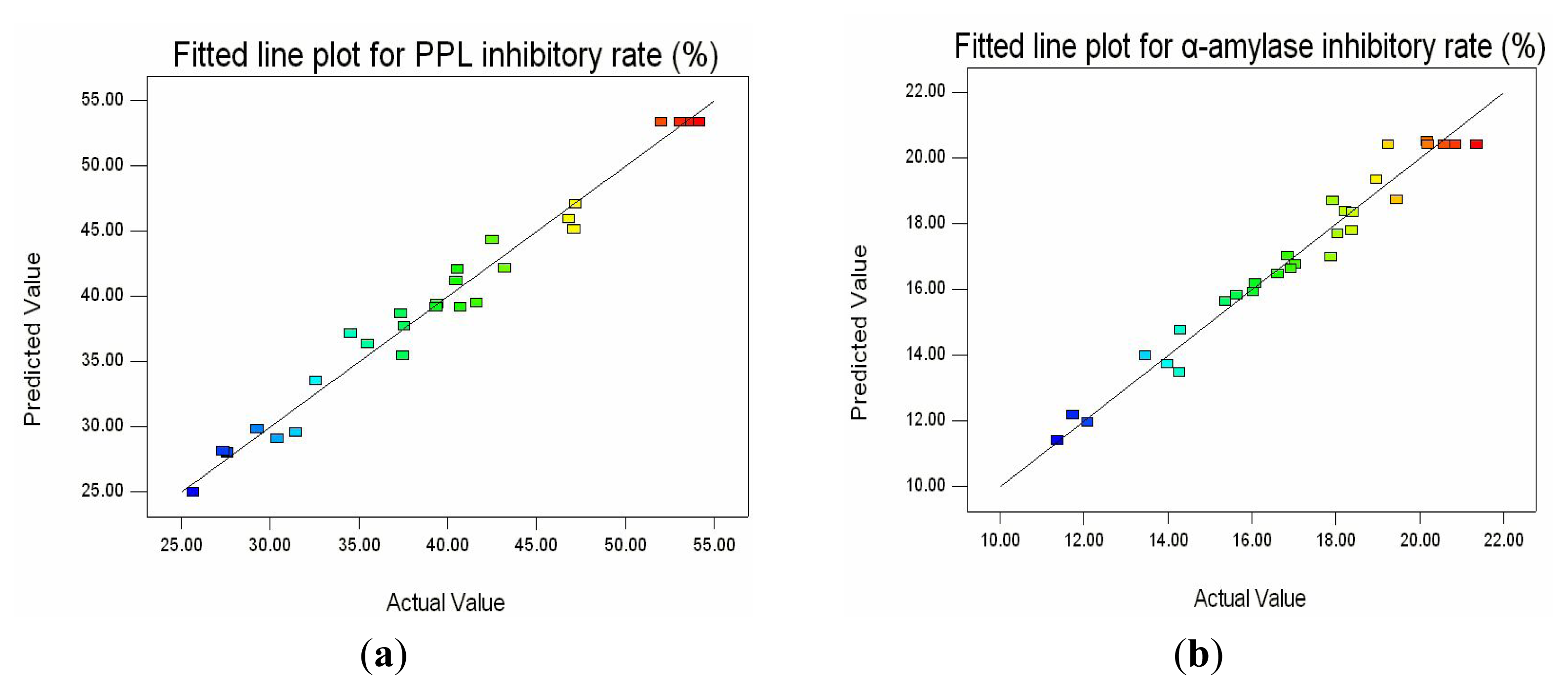
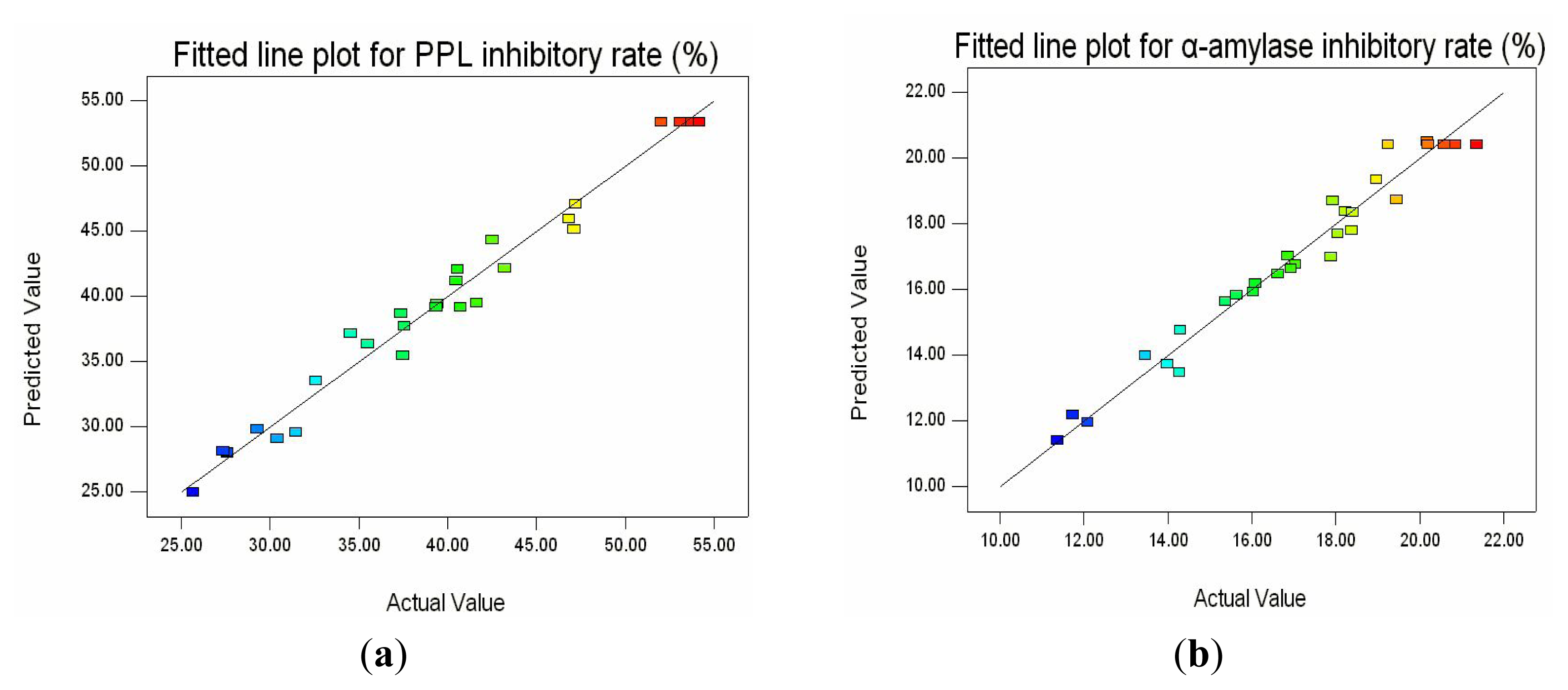
2.2. Fitting the Model
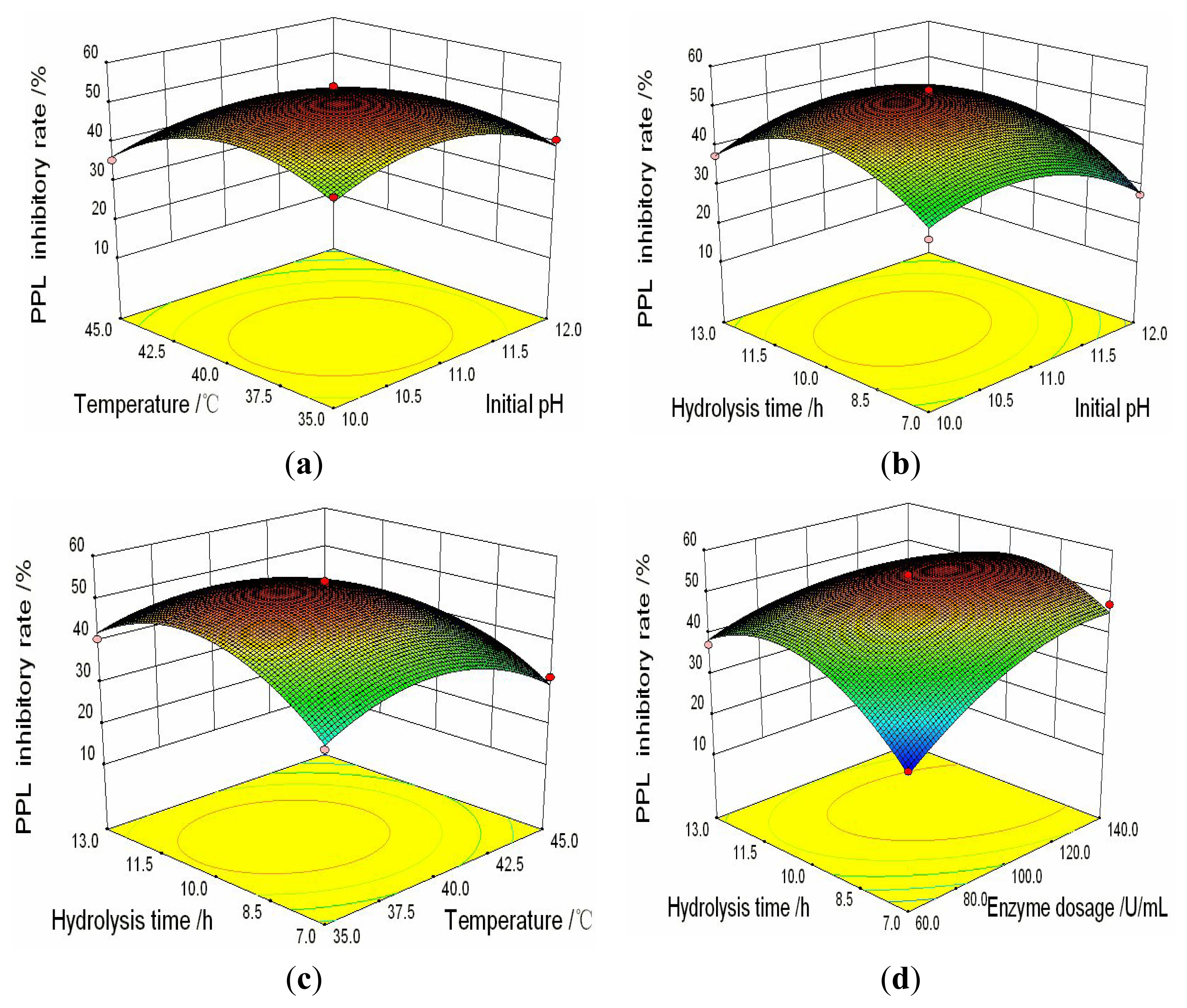
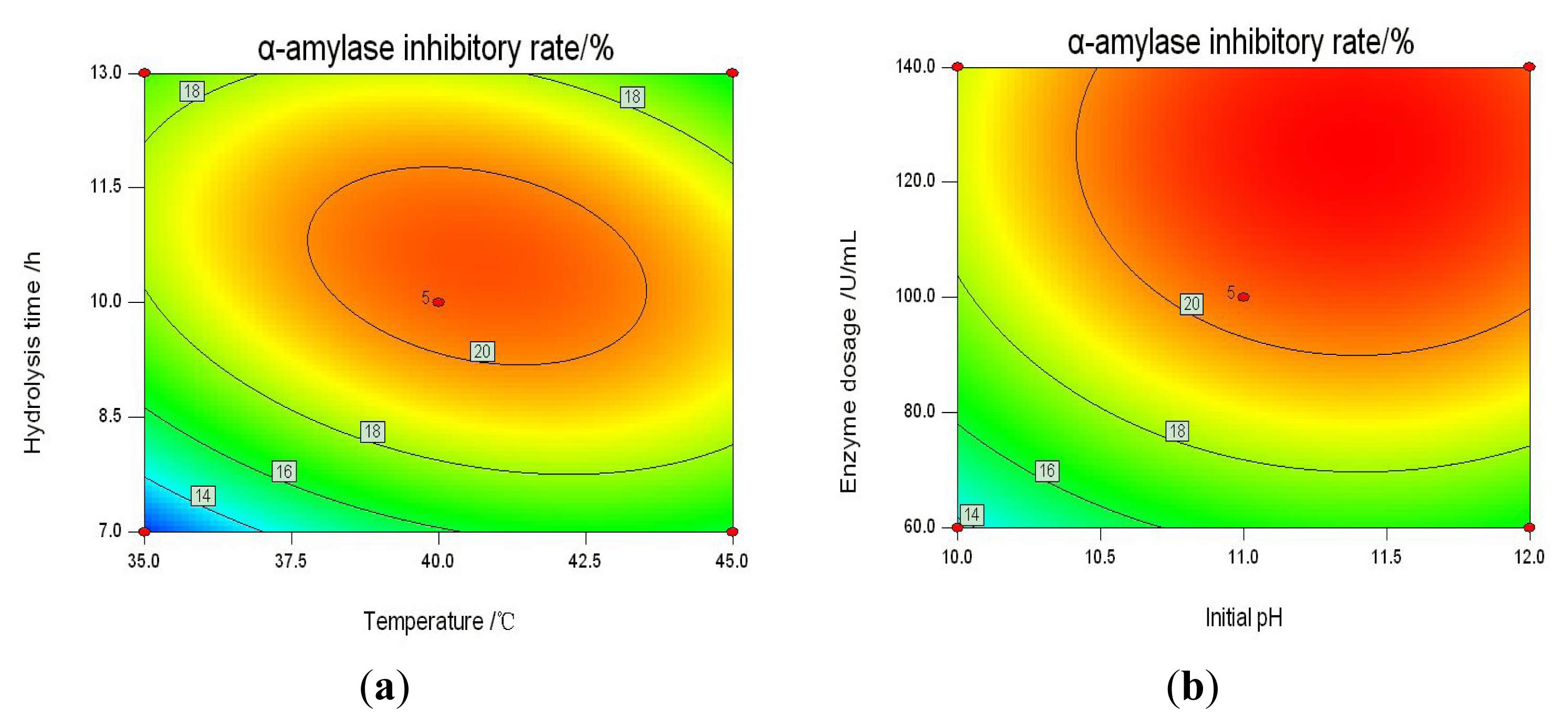
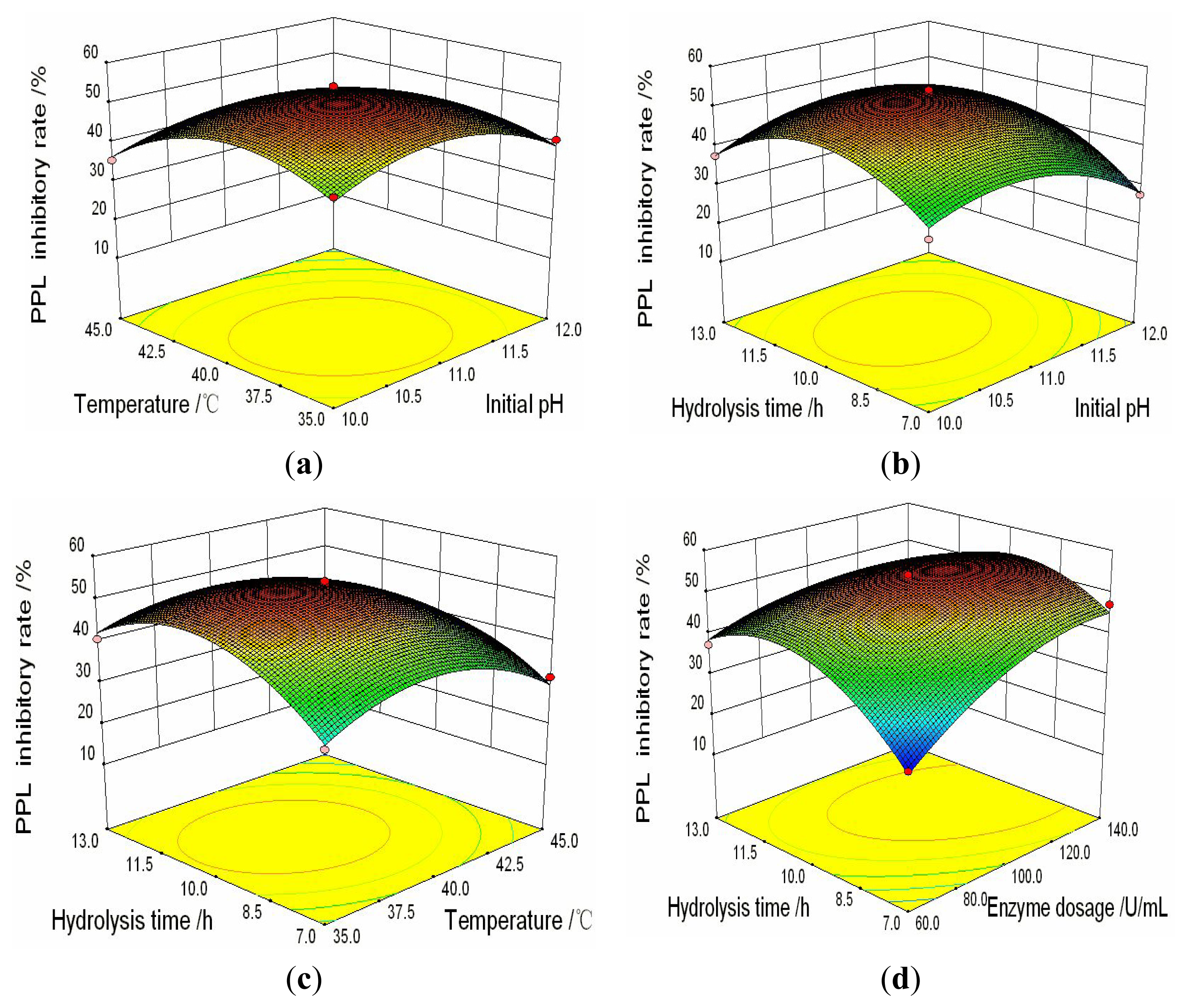
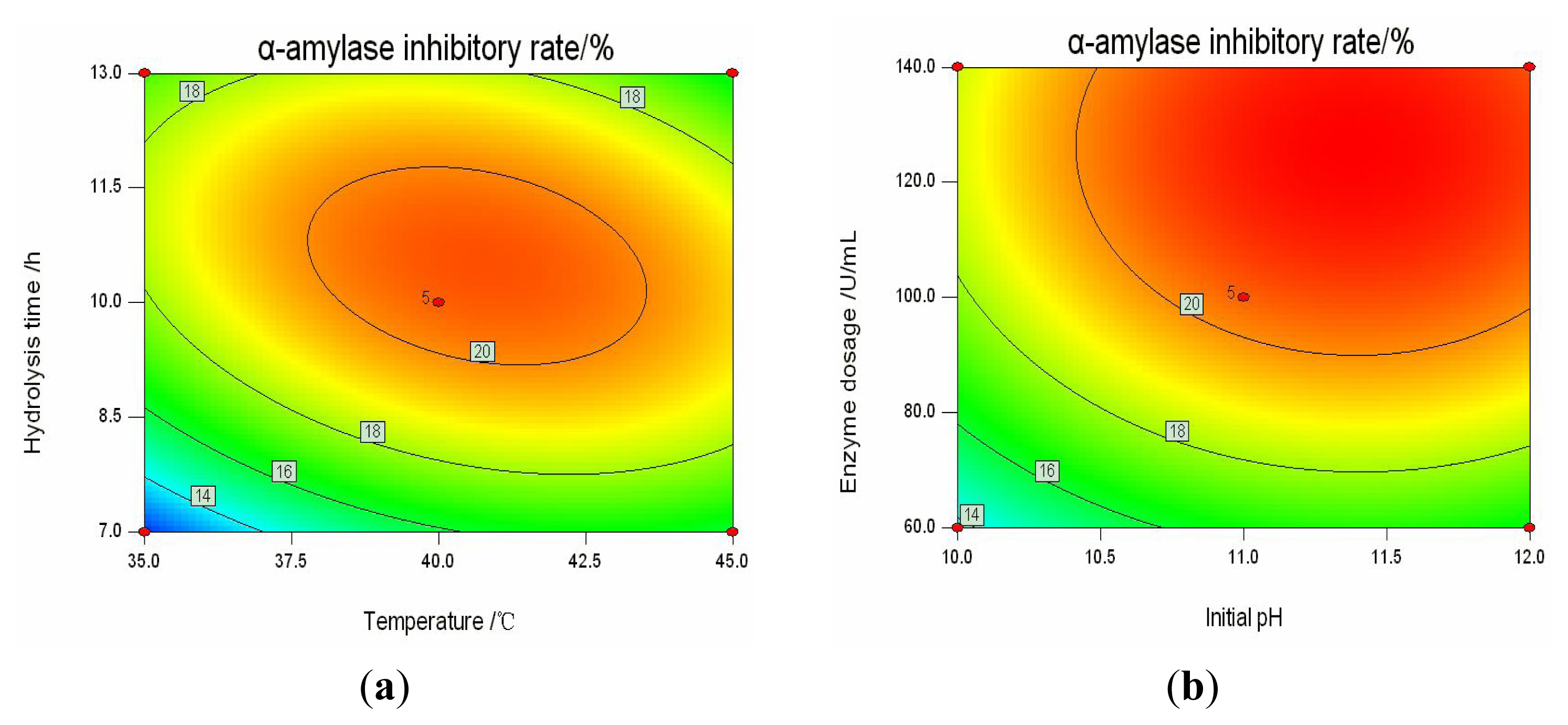
2.3. Analysis of Response Surfaces
2.4. Optimization of the Hydrolysis Conditions and Validation of Model
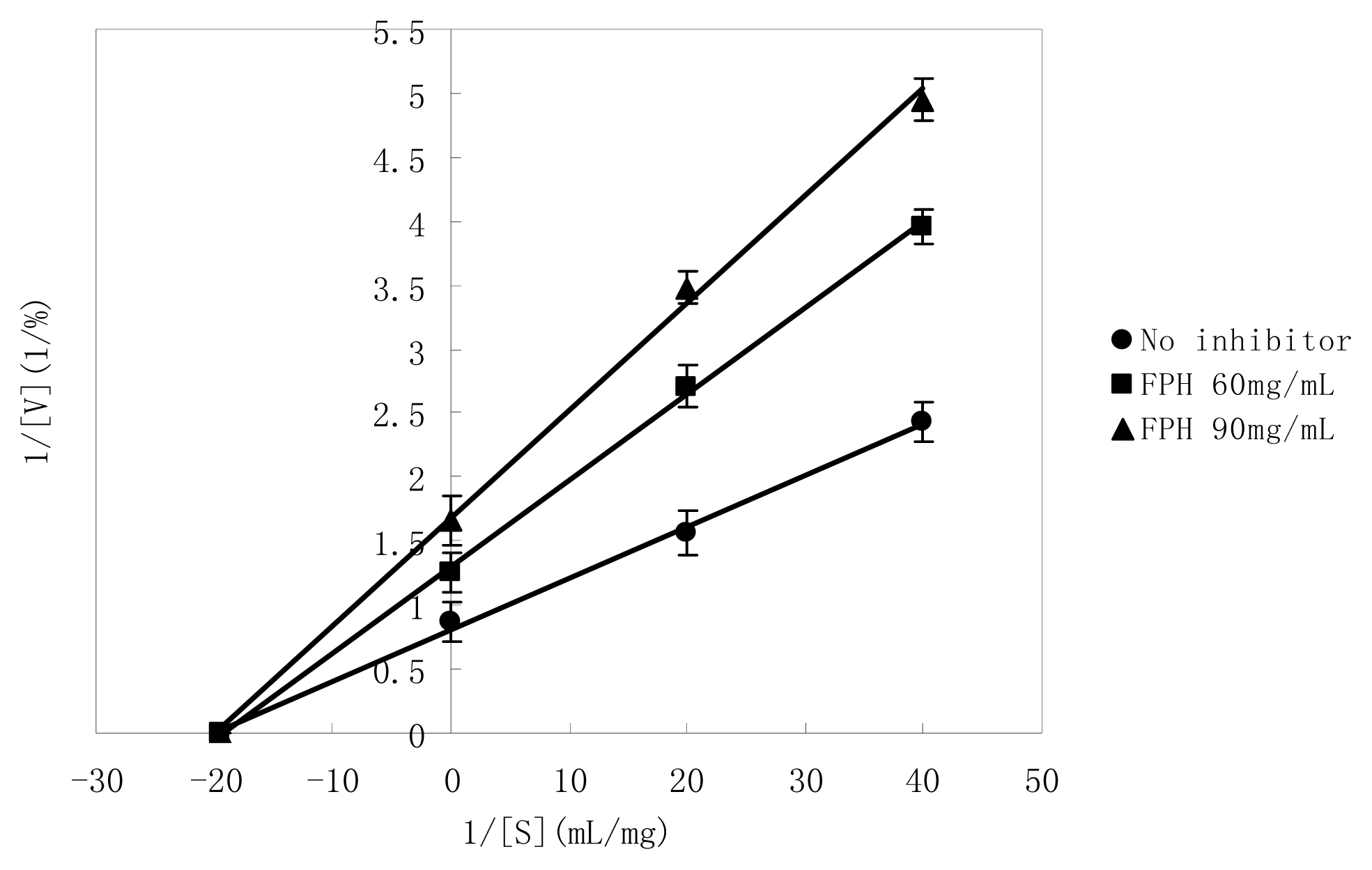
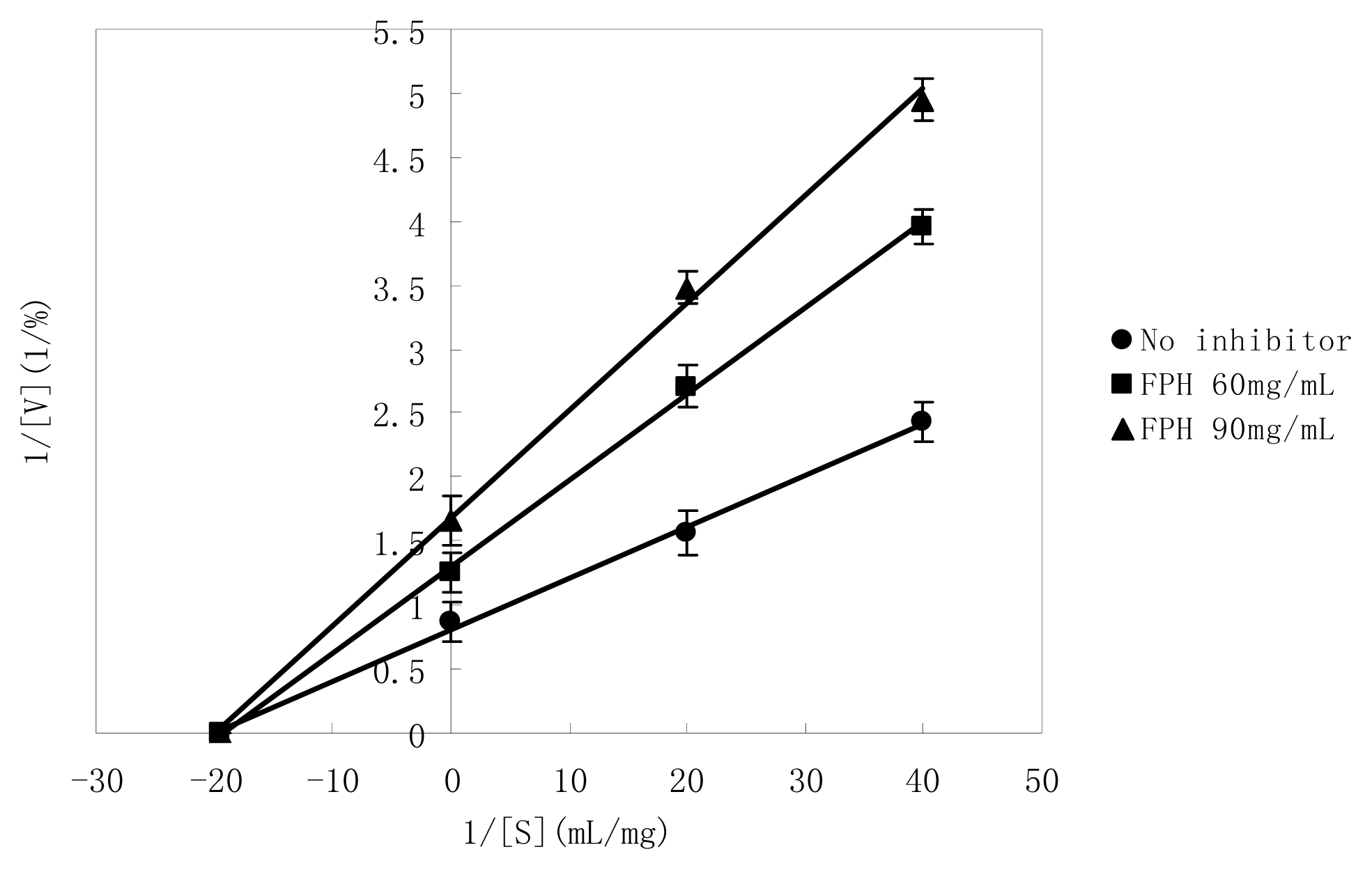
2.5. Manner of Inhibition by FPH
3. Experimental Section
3.1. Materials and Methods
3.1.1. Materials
3.1.2. Pretreatment and Enzymatic Hydrolysis
3.1.3. Assay for PPL Inhibitory Activity
3.1.4. Assay for α-Amylase Inhibitory Activity
3.1.5. Assay for Degree of Hydrolysis
3.1.6. Experimental Design and Statistical Analysis
4. Conclusions
References
- Hill, J.O.; Melanson, E.L.; Wyatt, H.T. Dietary fat intake and regulation of energy balance: Implications for obesity. J. Nutr 2000, 130, 284S–288S. [Google Scholar]
- Leslie, W.S.; Lean, M.E.J.; Baillie, H.M.; Hankey, C.R. Weight management: A comparison of existing dietary approaches in a work-site setting. Int. J. Obesity 2002, 26, 1469–1475. [Google Scholar]
- Leonhardt, M.; Hrupka, B.; Langhans, W. New approaches in the pharmacological treatment of obesity. Eur. J. Nutr 1999, 38, 1–13. [Google Scholar]
- Gargouri, Y.; Ransac, S.; Verger, R. Covalent inhibition of digestive lipases: An in vitro study. Biochim. Biophysica. Acta 1997, 1344, 6–37. [Google Scholar]
- McNeely, W.; Goa, K.L. Sibutramine: A review of its contribution to the management of obesity. Drugs 1998, 56, 1093–1124. [Google Scholar]
- Lee, S.H.; Park, M.H.; Heo, S.J.; Kang, S.M.; Ko, S.C.; Han, J.S.; Jeon, Y.J. Dieckol isolated from Ecklonia cava inhibits alpha-glucosidase and alpha-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol 2010, 48, 2633–2637. [Google Scholar]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-Anun, S.; Adisakwattana, S. Inhibitory activities of cyanidin and its glycosides and synergistic effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Int. J. Mol. Sci 2010, 11, 3387–3396. [Google Scholar]
- Baron, A.D. Postprandial hyperglycaemia and alpha-glucosidase inhibitors. Diabetes Res. Clin. Pract 1998, 40, S51–S55. [Google Scholar]
- Tsujita, T.; Matsuura, Y.; Okuda, H. Studies on the inhibition of pancreatic and carboxylester lipases by protamine. J. Lipid Res 1996, 37, 1481–1487. [Google Scholar]
- Borel, P.; Lairon, D.; Termine, E.; Grataroli, R.; Lafont, H. Isolation and properties of lipolysis inhibitory proteins from wheat germ and wheat bran. Plant Foods Hum. Nutr 1989, 39, 339–348. [Google Scholar]
- Tani, H.; Ohishi, H.; Watanabe, K. Purification and characterization of proteinous inhibitor of lipase from wheat flour. J. Agric. Food Chem 1994, 42, 2382–2385. [Google Scholar]
- Satouchi, K.; Mori, T.; Matsushita, S. Characterization of inhibitor protein for lipase in soybean seeds. Agric. Biol. Chem 1974, 38, 97–101. [Google Scholar]
- Sirtori, C.R.; Lovati, M.R.; Manzoni, C.; Castiglioni, S.; Duranti, M.; Magni, C.; Morandi, S.; D’Agostina, A.; Arnoldi, A. Proteins of white lupin seed, a naturally isoflavone-poor legume, reduce cholesterolemia in rats and increase LDL receptor activity in HepG2 cells. J. Nutr 2004, 134, 18–23. [Google Scholar]
- Tsutsumi, K.; Kawauchi, Y.; Kondo, Y.; Inoue, Y.; Koshitani, O.; Kohri, H. Water extract of defatted rice bran suppresses visceral fat accumulation in rats. J. Agric. Food Chem 2000, 48, 1653–1656. [Google Scholar]
- Niiho, Y.; Yamazaki, T.; Hosono, T.; Nakajima, Y.; Ishizaki, M.; Kurashige, T. Pharmacological studies on small peptide fraction derived soybean. Yakugaku Zasshi 1993, 113, 334–342. [Google Scholar]
- Hyun, J.K.; In, Y.B.; Chang-Won, A.; Suyong, L.; Hyeon, G.L. Purification and identification of adipogenesis inhibitory peptide from black soybean protein hydrolysate. Peptides 2007, 28, 2098–2103. [Google Scholar]
- Rho, S.J.; Park, S.; Ahn, C.W.; Shin, J.K.; Lee, H.G. Dietetic and hypocholesterolaemic action of black soy peptide in dietary obese rats. J. Sci. Food Agric 2007, 87, 908–913. [Google Scholar]
- Kagawa, K.; Matsutaka, H.; Fukuhama, C.; Watanabe, Y.; Fujino, H. Globin digest, acidic protease hydrolysate, inhibits dietary hypertriglyceridemia and Val-Val-Tyr-Pro, one of its constituents, possesses most superior effect. Life Sci 1996, 58, 1745–1755. [Google Scholar]
- Wergedahl, H.; Liaset, B.; Gudbrandsen, O.A.; Lied, E.; Espe, M.; Muna, Z.; Mork, S.; Berge, R.K. Fish protein hydrolysate reduces plasma total cholesterol, increases the proportion of HDL cholesterol, and lowers acyl-CoA: Cholesterol acyltransferase activity in liver of Zucker rats. J. Nutr 2004, 134, 1320–1327. [Google Scholar]
- Je, J.Y.; Park, P.J.; Kwon, J.Y.; Kim, S.K. A novel Angiotensin I converting enzyme inhibitory peptide from Alaska Pollack (Theragra chalcogramma) frame protein hydrolysate. J. Agric. Food Chem 2004, 52, 7842–7845. [Google Scholar]
- Je, J.Y.; Park, P.J.; Kim, S.K. Antioxidant activity of a peptide isolated from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. Food Res. Int 2005, 38, 45–50. [Google Scholar]
- Sampath Kumar, N.S.; Nazeer, R.A.; Jaiganesh, R. Purification and identification of antioxidant peptides from the skin protein hydrolysate of two marine fishes, horse mackerel (Magalaspis cordyla) and croaker (Otolithes ruber). Amino Acids 2012, 42, 1641–1649. [Google Scholar]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem 2007, 102, 1317–1327. [Google Scholar]
- Chabeaud, A.; Dutournié, P.; Guérard, F.; Vandanjon, L.; Bourseau, P. Application of response surface methodology to optimise the antioxidant activity of a Saithe (Pollachius virens) hydrolysate. Mar. Biotechnol 2009, 11, 445–455. [Google Scholar]
- Duartea, J.; Vinderola, G.; Ritzc, B.; Perdigón, G.; Matar, C. Immunomodulating capacity of commercial fish protein hydrolysate for diet supplementation. Immunobiology 2006, 211, 341–350. [Google Scholar]
- Murraya, A.L.; Pascho, R.J.; Alcornb, S.W.; Fairgrievec, W.T.; Shearerc, K.D.; Roley, D. Effects of various feed supplements containing fish protein hydrolysate or fish processing by-products on the innate immune functions of juvenile coho salmon (Oncorhynchus kisutch). Aquaculture 2003, 220, 643–653. [Google Scholar]
- Nilsang, S.; Lertsiri, S.; Suphantharika, M.; Assavanig, A. Optimization of enzymatic hydrolysis of fish soluble concentrate by commercial proteases. J. Food Eng 2005, 70, 571–578. [Google Scholar]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmentier, M. Influence of hydrolysis degree on the functional properties of salmon byproducts hydrolysates. J. Food Sci 2004, 69, 615–622. [Google Scholar]
- Bhaskar, N.; Benila, T.; Radha, C.; Lalitha, R.G. Optimization of enzymatic hydrolysis of visceral waste proteins of Catla (Catla catla) for preparing protein hydrolysate using a commercial protease. Bioresour. Technol 2008, 99, 335–343. [Google Scholar]
- Ovissipour, M.; Kenari, A.A.; Motamedzadegan, A.; Nazari, R.M. Optimization of enzymatic hydrolysis of visceral waste proteins of yellowfin tuna (Thunnus albacares). Food Bioprocess Technol 2012, 5, 696–705. [Google Scholar]
- Guerard, F.; Sumaya-Martinez, M.T.; Laroque, D.; Chabeaud, A.; Dufossé, L. Optimization of free radical scavenging activity by response surface methodology in the hydrolysis of shrimp processing discards. Process Biochem 2007, 42, 1486–1491. [Google Scholar]
- Dufossé, L.; De La Broise, D.; Guérard, F. Fish protein hydrolysates as nitrogen sources for microbial growth and metabolite production. Recent Res. Dev. Microbiol 1997, 1, 365–381. [Google Scholar]
- Normah, I.; Jamilah, B. Optimization of hydrolysis conditions for the production of thread finbream ( Nemipterus japonicus) hydrolysate by Alcalase. J. Muscle Food. 2005, 16, 87–102. [Google Scholar]
- Ren, J.Y.; Zhao, M.M.; Shi, J.; Wang, J.S.; Jiang, Y.M.; Cui, C.; Kakuda, Y.; Xue, S.J. Optimization of anti-oxidant peptide production from grass carp sarcoplasmic protein using response surface methodology. Food Sci. Technol 2008, 41, 1624–1632. [Google Scholar]
- Zhuang, Y.L.; Zhao, X.; Li, B.F. Optimization of antioxidant activity by response surface methodology in hydrolysates of jellyfish (Rhopilema esculentum) umbrella collagen. J. Zhejiang Univ.-Sci. B 2009, 10, 572–579. [Google Scholar]
- Hou, L.X.; Wang, J.S.; Zhang, D. Optimization of debittering of soybean antioxidant hydrolysates with β-cyclodextrins using response surface methodology. J. Food Sci. Technol 2011. [Google Scholar] [CrossRef]
- Habulin, M.; Knez, Z. High-pressure enzymatic hydrolysis of oil. Eur. J. Lipid Sci. Technol 2002, 104, 381–386. [Google Scholar]
- Cao, W.H.; Zhang, C.H.; Hong, P.Z.; Ji, H.W. Response surface methodology for autolysis parameters optimization of shrimp head and amino acids released during autolysis. Food Chem 2008, 109, 176–183. [Google Scholar]
- Song, R.; Wei, R.B.; Zhang, B.; Wang, D.F. Optimization of the antibacterial activity of Half-Fin Anchovy (Setipinna taty) Hydrolysates. Food Bioprocess Technol 2012, 5, 1979–1989. [Google Scholar]
- Li, Y.Q.; Yang, P.; Gao, F.; Zhang, Z.W.; Wu, B. Probing the interaction between 3 flavonoids and pancreatic lipase by methods of fluorescence spectroscopy and enzymatic kinetics. Eur. Food Res. Technol 2011, 233, 63–69. [Google Scholar]
- Kim, J.H.; Kim, H.J.; Park, H.W.; Youn, S.H.; Choi, D.Y.; Shin, C.S. Development of inhibitors against lipase and alpha-glucosidase from derivatives of monascus pigment. FEMS Microbiol. Lett 2007, 276, 93–98. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem 1959, 3, 426–428. [Google Scholar]
- The United States Pharmacopeial Convention, United States Pharmacopea, 22nd ed; Rockville, MD, USA, 1989.
- Association of Official Analytical Chemists, Official Methods of Analysis, 15th ed; Washington: DC, USA, 1990.
- Yemul, O.; Imae, T. Covalent-bonded immobilization of lipase on poly(phenylene sulfide) dendrimers and their hydrolysis ability. Biomacromolecules 2005, 6, 2809–2814. [Google Scholar]
- Yadav, G.D.; Devi, K.M. Kinetics of hydrolysis of tetrahydrofurfuryl butyrate in a three phase system containing immobilized lipase from Candida Antarctica. Biochem. Eng. J 2003, 17, 57–63. [Google Scholar]
- Kaambre, T.; Tougu, V.; Kaambre, P.; Vija, H.; Sikk, P. Hydrolysis of emulsified mixtures of triacylglycerols by pancreatic lipase. Biochim. Biophys. Acta 1999, 1431, 97–106. [Google Scholar]
- Linfield, W.M.; Barauskas, R.A.; Sivieri, L.; Serota, S.; Stevenson, R.W. Enzymatic fat hydrolysis and synthesis. J. Am. Oil Chem. Soc 1984, 61, 191–195. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RUN | Independent variables | Response (Y1) PPL inhibitory rate (%) | Response (Y2) α-amylase inhibitory rate (%) | |||
|---|---|---|---|---|---|---|
| Initial pH (χ1) | Temperature (χ2), °C | Enzyme dosage (χ3), U/mL | Hydrolysis time (χ4), h | |||
| 1 | 0 | −1 | 1 | 0 | 47.19 ± 1.24 | 19.44 ± 0.82 |
| 2 a | 0 | 0 | 0 | 0 | 53.54 ± 1.15 | 20.58 ± 0.69 |
| 3 | 0 | 0 | 1 | 1 | 39.39 ± 0.53 | 18.22 ± 0.77 |
| 4 | 1 | −1 | 0 | 0 | 40.71 ± 1.01 | 17.92 ± 1.04 |
| 5 | 1 | 1 | 0 | 0 | 27.58 ± 0.93 | 18.04 ± 1.12 |
| 6 a | 0 | 0 | 0 | 0 | 53.07 ± 1.52 | 19.24 ± 0.62 |
| 7 | 1 | 0 | 0 | −1 | 27.56 ± 1.33 | 16.61 ± 0.45 |
| 8 | −1 | 0 | 0 | 1 | 37.54 ± 1.49 | 16.92 ± 0.51 |
| 9 | −1 | −1 | 0 | 0 | 43.19 ± 0.58 | 13.45 ± 0.44 |
| 10 | 1 | 0 | −1 | 0 | 29.27 ± 1.02 | 16.08 ± 0.73 |
| 11 | 0 | 1 | −1 | 0 | 27.32 ± 0.75 | 15.36 ± 0.77 |
| 12 | 0 | 0 | −1 | −1 | 25.63 ± 1.65 | 11.36 ± 0.36 |
| 13 | 0 | 1 | 1 | 0 | 40.45 ± 0.87 | 18.96 ± 0.32 |
| 14 a | 0 | 0 | 0 | 0 | 54.15 ± 1.07 | 20.19 ± 0.54 |
| 15 | 1 | 0 | 1 | 0 | 42.50 ± 1.44 | 20.17 ± 0.62 |
| 16 | −1 | 1 | 0 | 0 | 35.48 ± 0.95 | 18.37 ± 0.65 |
| 17 | 0 | −1 | −1 | 0 | 39.32 ± 1.27 | 14.27 ± 0.49 |
| 18 | 1 | 0 | 0 | 1 | 37.47 ± 0.82 | 17.88 ± 0.61 |
| 19 | 0 | 1 | 0 | −1 | 31.44 ± 0.98 | 16.02 ± 0.32 |
| 20 | −1 | 0 | 0 | −1 | 34.52 ± 1.02 | 11.73 ± 0.47 |
| 21 | −1 | 0 | −1 | 0 | 41.61 ± 1.31 | 13.98 ± 0.56 |
| 22 | 0 | 0 | −1 | 1 | 37.36 ± 0.64 | 14.29 ± 0.55 |
| 23 | 0 | 1 | 0 | 1 | 30.39 ± 1.16 | 15.63 ± 0.69 |
| 24 | 0 | −1 | 0 | −1 | 32.56 ± 1.09 | 12.08 ± 0.44 |
| 25 a | 0 | 0 | 0 | 0 | 54.16 ± 0.85 | 20.83 ± 0.39 |
| 26 | 0 | 0 | 1 | −1 | 47.09 ± 0.72 | 17.03 ± 0.52 |
| 27 a | 0 | 0 | 0 | 0 | 52.01 ± 0.64 | 21.35 ± 0.42 |
| 28 | 0 | −1 | 0 | 1 | 40.53 ± 0.58 | 16.85 ± 0.31 |
| 29 | −1 | 0 | 1 | 0 | 46.82 ± 0.62 | 18.39 ± 0.37 |
| Source | The PPL inhibitory rate (%) | The α-amylase inhibitory rate (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Sum of squares | Mean square | F-value | p-value | Sum of squares | Mean square | F-value | p-value | |
| Model | 2104.13 | 150.29 | 48.88 | <0.0001 ** | 209.26 | 14.95 | 30.14 | <0.0001 ** |
| χ1 | 96.73 | 96.73 | 31.46 | <0.0001 ** | 16.01 | 16.01 | 32.28 | <0.0001 ** |
| χ2 | 215.39 | 215.39 | 70.05 | <0.0001 ** | 5.84 | 5.84 | 11.77 | 0.0041 ** |
| χ3 | 330.02 | 330.02 | 107.33 | <0.0001 ** | 60.17 | 60.17 | 121.34 | <0.0001 ** |
| χ4 | 47.52 | 47.52 | 15.46 | 0.0015 ** | 18.65 | 18.65 | 37.61 | <0.0001 ** |
| χ1χ2 | 7.34 | 7.34 | 2.39 | 0.1445 ns | 5.76 | 5.76 | 11.62 | 0.0042 ** |
| χ1χ3 | 16.08 | 16.08 | 5.23 | 0.0383 * | 0.026 | 0.026 | 0.052 | 0.8235 ns |
| χ1χ4 | 11.87 | 11.87 | 3.86 | 0.0696 ns | 3.84 | 3.84 | 7.75 | 0.0147 * |
| χ2χ3 | 6.92 | 6.92 | 2.25 | 0.1559 ns | 0.62 | 0.62 | 1.24 | 0.2837 ns |
| χ2χ4 | 20.34 | 20.34 | 6.62 | 0.0222 * | 6.66 | 6.66 | 13.42 | 0.0026 ** |
| χ3χ4 | 94.38 | 94.38 | 30.70 | <0.0001 ** | 0.76 | 0.76 | 1.53 | 0.2370 ns |
| χ12 | 409.97 | 409.97 | 133.33 | <0.0001 ** | 14.34 | 14.34 | 28.92 | <0.0001 ** |
| χ22 | 520.90 | 520.90 | 169.41 | <0.0001 ** | 22.94 | 22.94 | 46.27 | <0.0001 ** |
| χ32 | 195.33 | 195.33 | 63.53 | <0.0001 ** | 19.54 | 19.54 | 39.41 | <0.0001 ** |
| χ42 | 756.07 | 756.07 | 245.90 | <0.0001 ** | 72.88 | 72.88 | 146.98 | <0.0001 ** |
| Residual | 43.05 | 3.07 | - | - | 6.94 | 0.50 | - | - |
| Lack of fit | 39.85 | 3.98 | 4.98 | 0.0678 ns | 4.44 | 0.44 | 0.71 | 0.7002 ns |
| Pure error | 3.20 | 0.80 | - | - | 2.50 | 0.63 | - | - |
| Total | 2147.17 | - | - | - | 216.20 | - | - | - |
| Variables | Coded level | ||
|---|---|---|---|
| −1 | 0 | +1 | |
| Initial pH (X1) | 10 | 11 | 12 |
| Temperature (X2), °C | 35 | 40 | 45 |
| Enzyme dosage (X3), U/mL | 60 | 100 | 140 |
| Hydrolysis time (X4), h | 7 | 10 | 13 |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, L.; Wang, Y.; Peng, C.; Wang, J. Optimization of the Preparation of Fish Protein Anti-Obesity Hydrolysates Using Response Surface Methodology. Int. J. Mol. Sci. 2013, 14, 3124-3139. https://doi.org/10.3390/ijms14023124
Liu L, Wang Y, Peng C, Wang J. Optimization of the Preparation of Fish Protein Anti-Obesity Hydrolysates Using Response Surface Methodology. International Journal of Molecular Sciences. 2013; 14(2):3124-3139. https://doi.org/10.3390/ijms14023124
Chicago/Turabian StyleLiu, Liyuan, Yanping Wang, Chen Peng, and Jinju Wang. 2013. "Optimization of the Preparation of Fish Protein Anti-Obesity Hydrolysates Using Response Surface Methodology" International Journal of Molecular Sciences 14, no. 2: 3124-3139. https://doi.org/10.3390/ijms14023124
APA StyleLiu, L., Wang, Y., Peng, C., & Wang, J. (2013). Optimization of the Preparation of Fish Protein Anti-Obesity Hydrolysates Using Response Surface Methodology. International Journal of Molecular Sciences, 14(2), 3124-3139. https://doi.org/10.3390/ijms14023124