Abstract
Lung cancer is the most common cause of cancer-related death. Nonetheless, a decrease in overall incidence and mortality has been observed in the last 30 years due to prevention strategies and improvements in the use of chemotherapeutic agents. In recent studies, Simvastatin (SIM) has demonstrated anti-tumor activity, as well as potent chemopreventive action. As an inhibitor of 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMG-CoA), SIM has been shown to stimulate apoptotic cell death. In this study, an MTT assay revealed the cytotoxic activity of SIM against human large cell lung cancer (Non-small cell lung cancer; NSCLC) cells (NCI-H460); however, induced apoptosis was not observed in NCI-H460 cells. Protein expression levels of cell cycle regulating proteins Cdk4, Cyclin D1, p16 and p27 were markedly altered by SIM. Collectively, our results indicate that SIM inhibits cell proliferation and arrests NCI-H460 cell cycle progression via inhibition of cyclin-dependent kinases and cyclins and the enhancement of CDK inhibitors p16 and p27. Our findings suggest that, in addition to the known effects on hypercholesterolemia therapy, SIM may also provide antitumor activity in established NSCLC.
1. Introduction
Lung cancer is the leading cause of cancer-related death worldwide. The survival rate of lung carcinoma has remained relatively unchanged below 15% for the past two decades [1]. NSCLC accounts for approximately 80% of all forms of lung cancer, and early-stage NSCLC is potentially curative through complete surgical resection. Nonetheless, the overall five-year survival rate for these patients is only 45% to 70%, due to metastatic recurrence [2].
Statins have been shown to alter several cellular mechanisms, resulting in apoptosis and reduced tumor cell growth, as well as angiogenesis and impaired metastatic processes [3]. Simvastatin (SIM), like statins, is used in hypercholesterolemia therapy to inhibit the 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase, which catalyzes the conversion of HMG-CoA to mevalonate [4]. In addition to its cholesterol-lowering activity, evidence is mounting to support claims that SIM may have potential anti-cancer benefits [5,6]. SIM appears to mediate angiogenesis, cell proliferation, distant metastasis, cell cycle arrest and apoptosis, as well as suppress NF-κB activation [7,8]. The anti-tumor effects of SIM have been demonstrated in a variety of tumors, including myeloma cells [9], breast cancer [10], leukemia [11], prostate cancer [12], malignant glioblastoma [13] and hepatoma [14]; however, the anti-tumor effects of SIM on NSCLC have not been studied in depth.
This study characterized the anti-proliferation, cell cycle arrest and anti-tumorigenesis activities of Simvastatin on NCI-H460 cells. Our results provide valuable data for the further development of treatments for NSCLC.
2. Results and Discussion
2.1. SIM Inhibits the Proliferation of NCI-H460 Cells
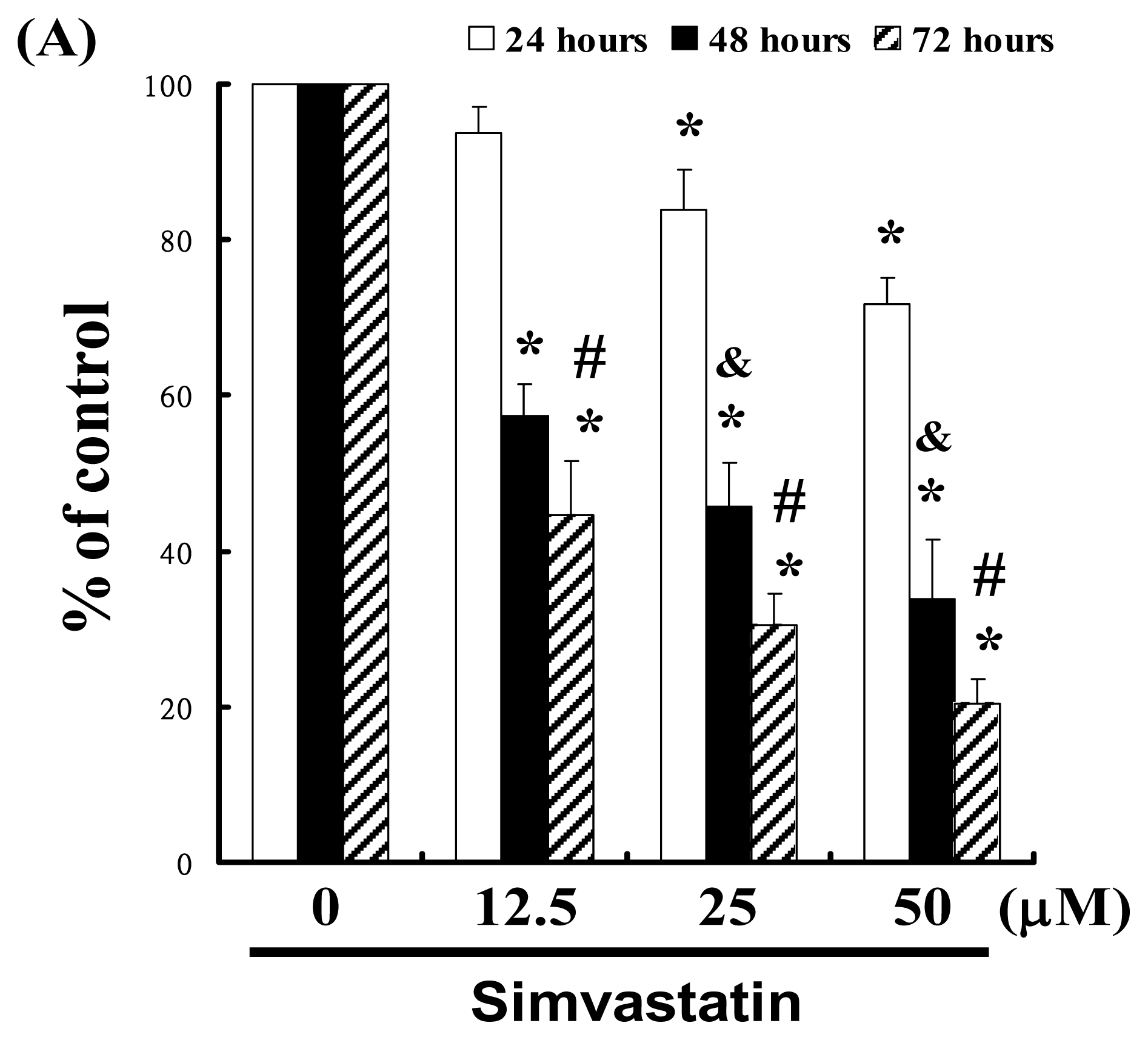
For this study, we hypothesized that SIM could mediate the survival of NCI-H460 cells by inhibiting growth. To explore the anti-tumor activity of SIM against NCI-H460 cells, we performed an in vitro cell viability study in which NCI-H460 cell samples were respectively treated with increasing doses of SIM (0, 12.5, 25, and 50 μM) for 24 to 72 h. The results in Figure 1A indicate that SIM treatment impaired the survival and proliferation of NCI-H460 cells in a dose-dependent manner. The 24 h IC50 of Simvastatin in the NCI-H460 cancer cells was determined to be 86.09 μM; y = −1.7209x + 172.14, R2 = 0.9899 (* p < 0.05 vs. SIM 0 μM group) in a time-course experiment (& p < 0.05 vs. 24 h treatment, # p < 0.05 vs. 48 h treatment). The cell viability was assayed by trypan blue exclusion and cell counting (Data not shown).
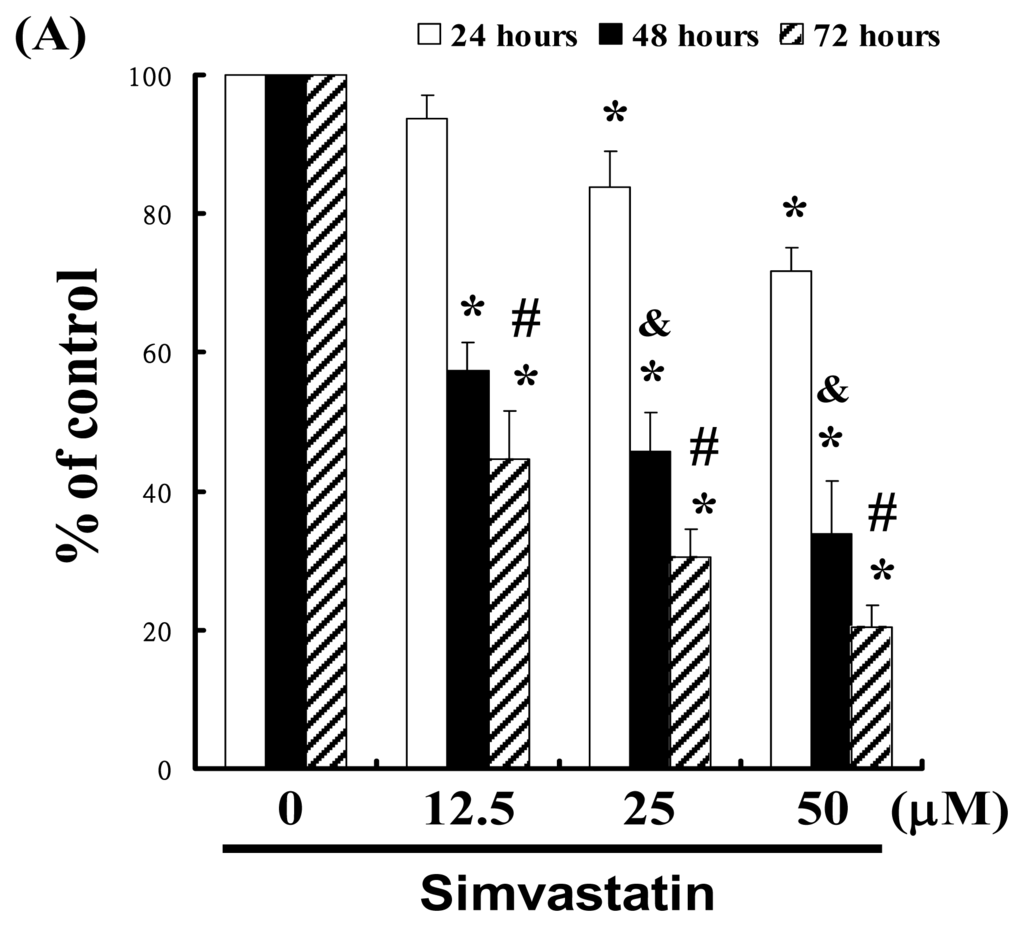
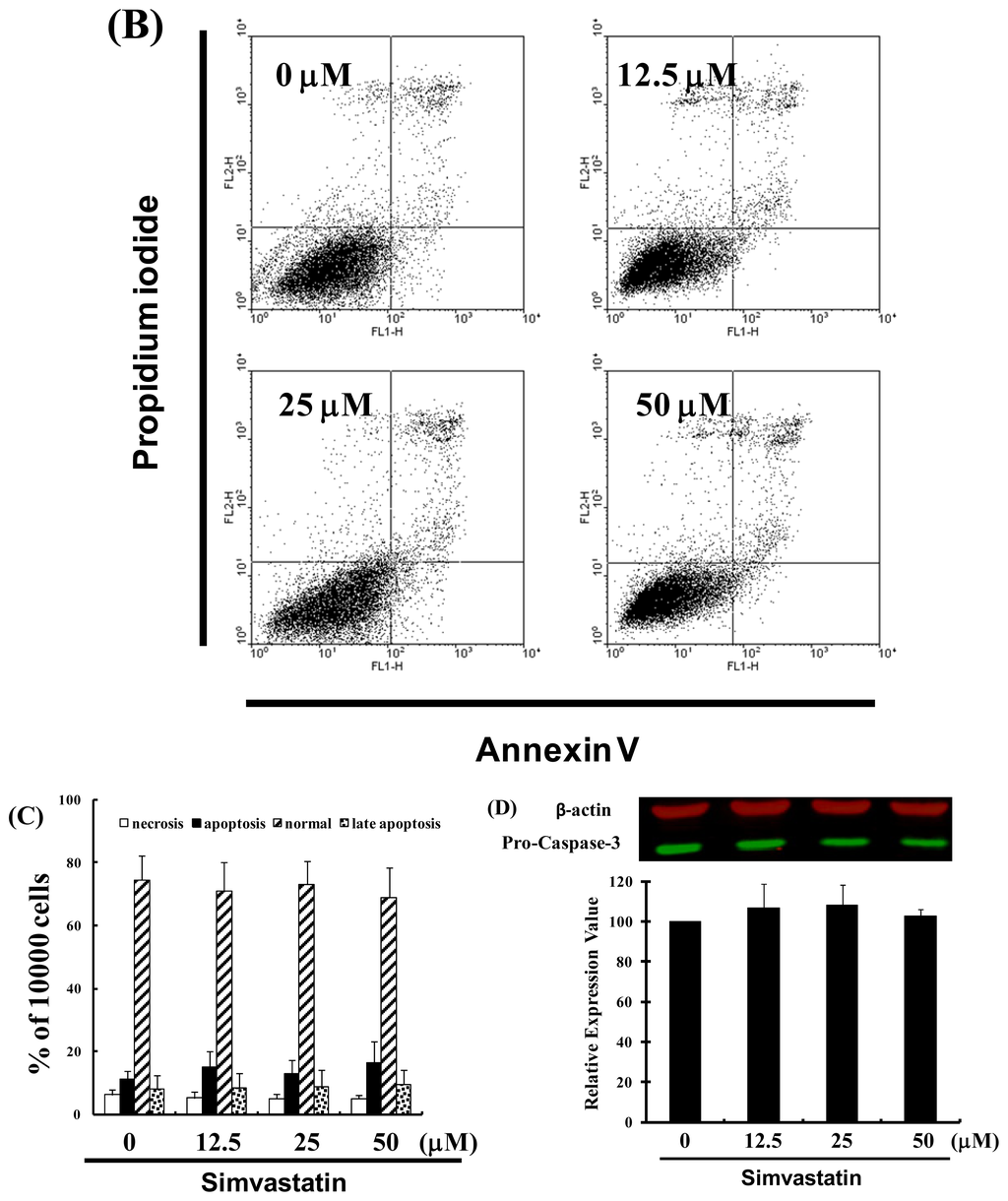
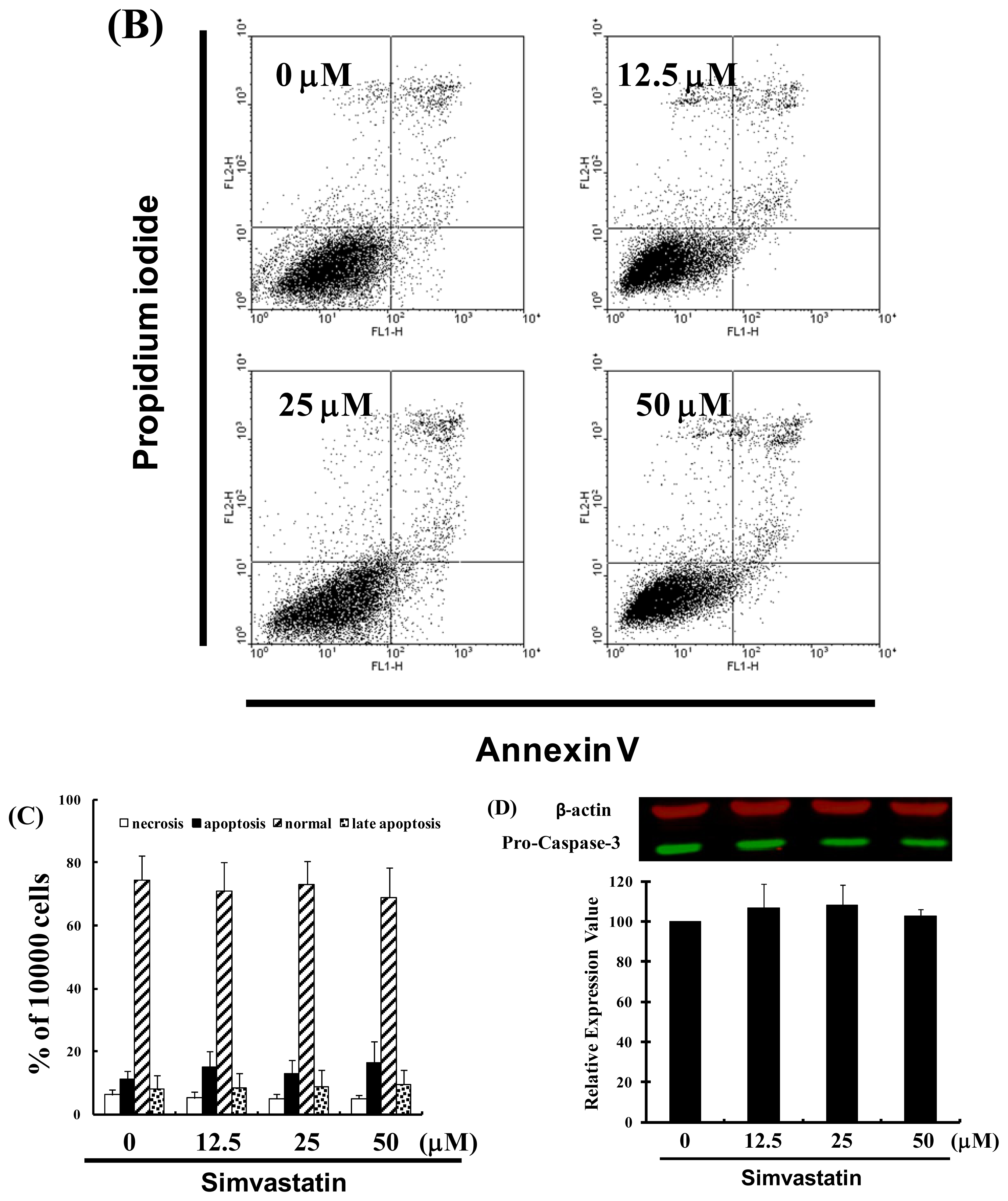
Figure 1.
Simvastatin (SIM) mediated the survival of NCI-H460 cells, thereby inhibiting proliferation: (A) In vitro study involved the treatment of NCI-H460 cells with increasing doses of SIM for 24 to 72 h. The survival of the SIM-treated cancer cells was then determined using an MTT assay; (B) Influence of SIM on apoptosis in NCI-H460 cells; (C) Total apoptosis in NCI-H460 cells following 4 hours of incubation with SIM; (D) Caspase-3 activation in NCI-H460 cells induced by SIM treatment. Cells were treated with SIM, and proteins were subsequently analyzed via Western blot analysis. Band intensities (pro-caspase 3) were quantified using a Li-COR near infrared imaging system. Statistical analysis included the t-test. Symbols (*, & and #) in each group of bars indicate that the difference resulting from treatment with SIM is statistically significant at p < 0.05.
2.2. SIM Impaired the Viability of NCI-H460 Cells without Inducing Apoptosis
We also conducted a study on apoptosis to elucidate the anti-cancer mechanism associated with SIM in NCI-H460 cells. After treating the cells with various doses of SIM, the percentage of apoptotic cells was assessed using Annexin V-FITC and propidium iodide staining, followed by flow cytometry (Figure 1B). A dot-plot of Annexin V-FITC fluorescence versus PI fluorescence indicated a non-significant increase in the percentage of apoptotic cells after treatment with SIM. At SIM concentrations of 12.5 to 50 μM, no significant increase was observed in the percentage of cells undergoing necrosis or apoptosis (Figure 1C) or caspase-3 activation (Figure 1D). The results summarized in Figure 1 indicate that SIM could mediate the survival of NCI-H460 cells and, thus, inhibit their proliferation not via non-apoptotic pathways.
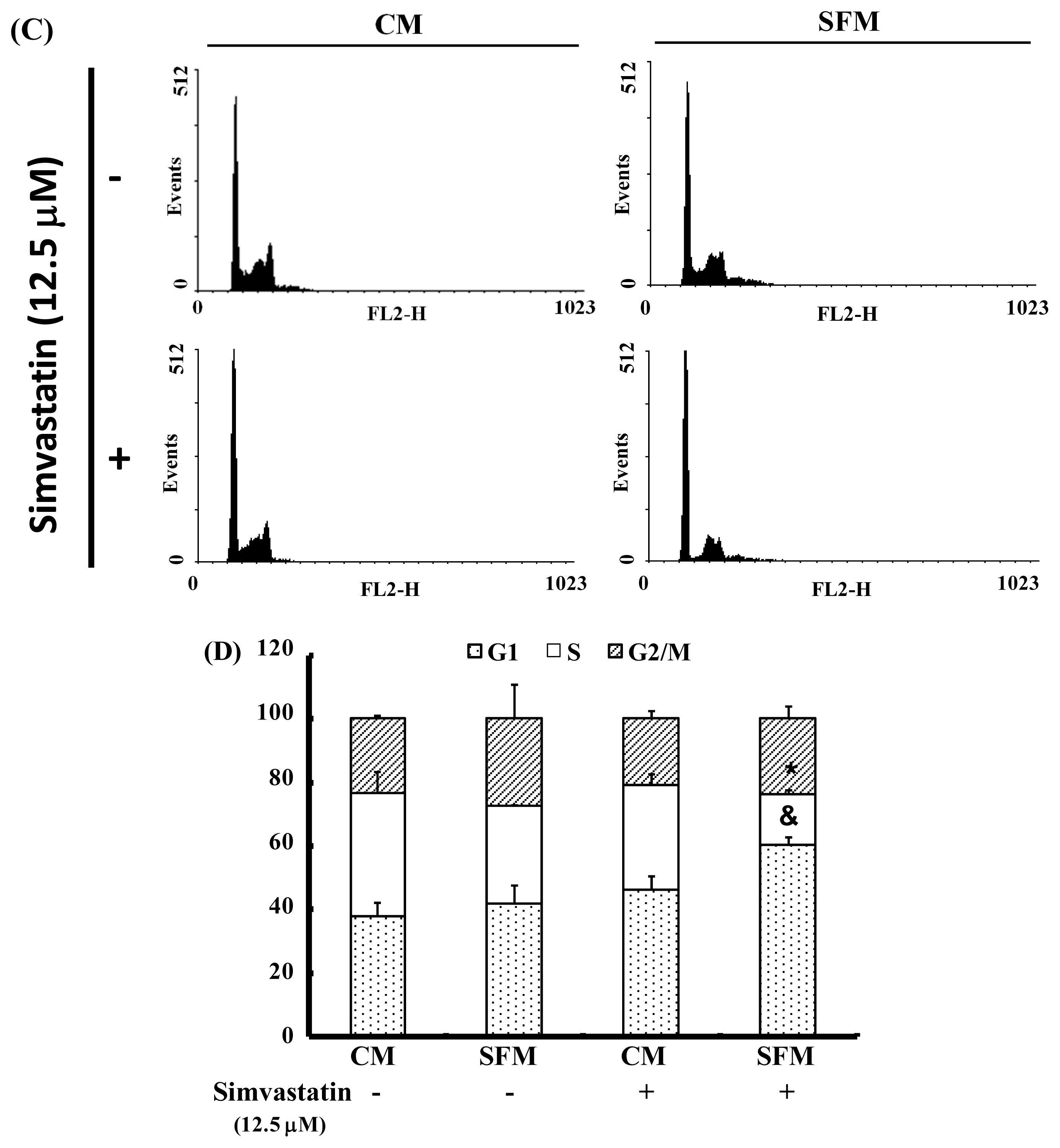
2.3. SIM Treatment Induced Accumulation of G1 Phase in NCI-H460 Cells
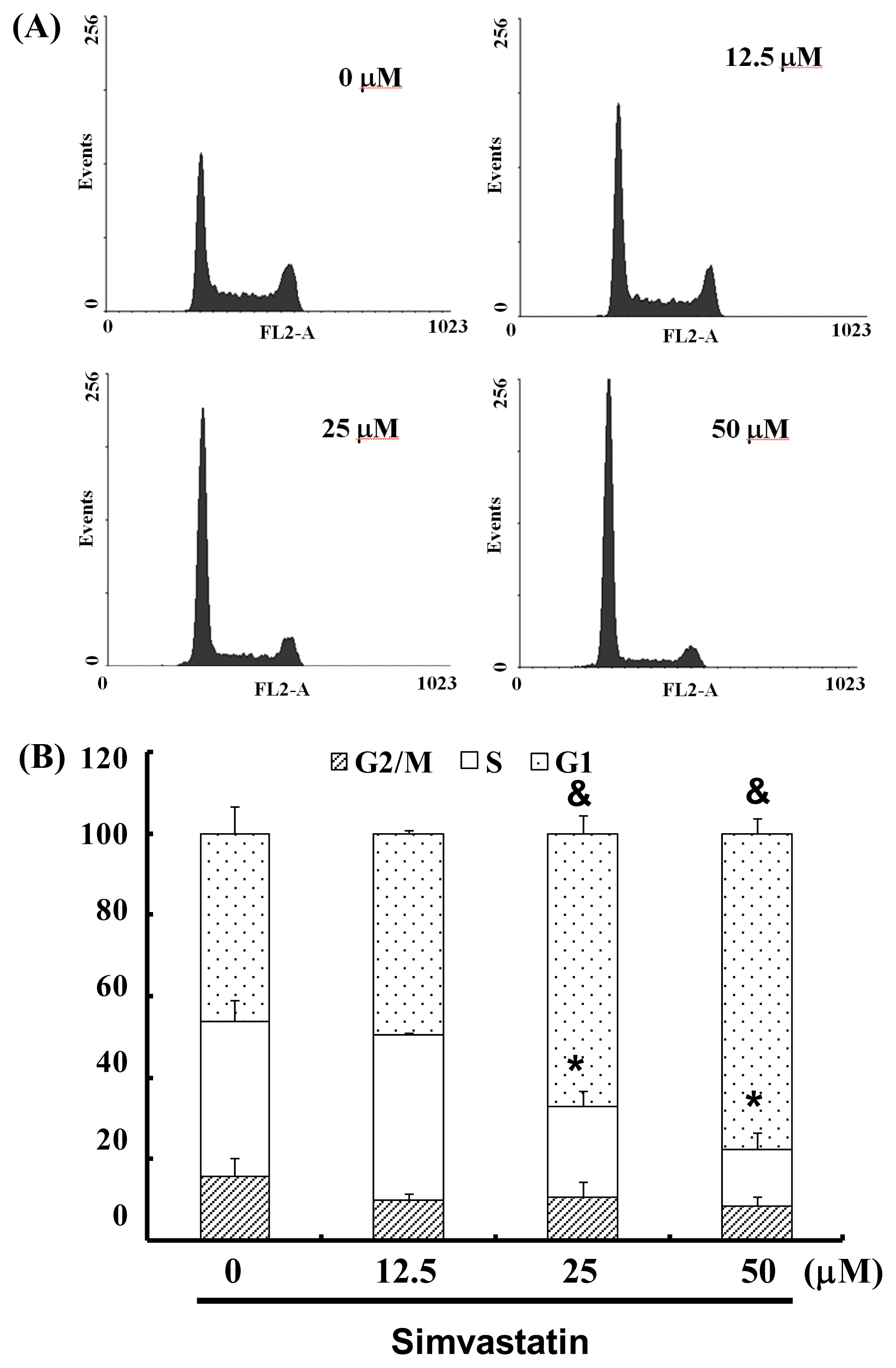
The cell-cycle distribution of SIM-treated NCI-H460 cells was analyzed using flow cytometry. Prior to processing and analysis, cells were exposed to SIM for 24 h. As illustrated in Figure 2A, the exposed cells showed an increase in the population of G1 phase cells, compared with untreated cells. These observations imply that the NCI-H460 cells may have undergone cell cycle arrest. Treatment with SIM increased the number of G1 phase cells (& p < 0.05 vs. SIM 0 μM) and simultaneously reduced the number of S phase cells (* p < 0.05 vs. SIM 0 μM) (Figure 2B). To investigate the role of SIM in G1 cell cycle arrest, we employed a serum deprivation (serum free) procedure to synchronize NCI-H460 cells. We chose this method due to its simplicity, reliability and reversibility. The addition of SIM increased the number of cells in G1 phase (& p < 0.05 vs. SIM 0 μM) and simultaneously reduced the number of cells in S phase (* p < 0.05 vs. SIM 0 μM) in CM (complete medium) and SFM (serum free medium), as shown in Figure 2C,D, respectively.
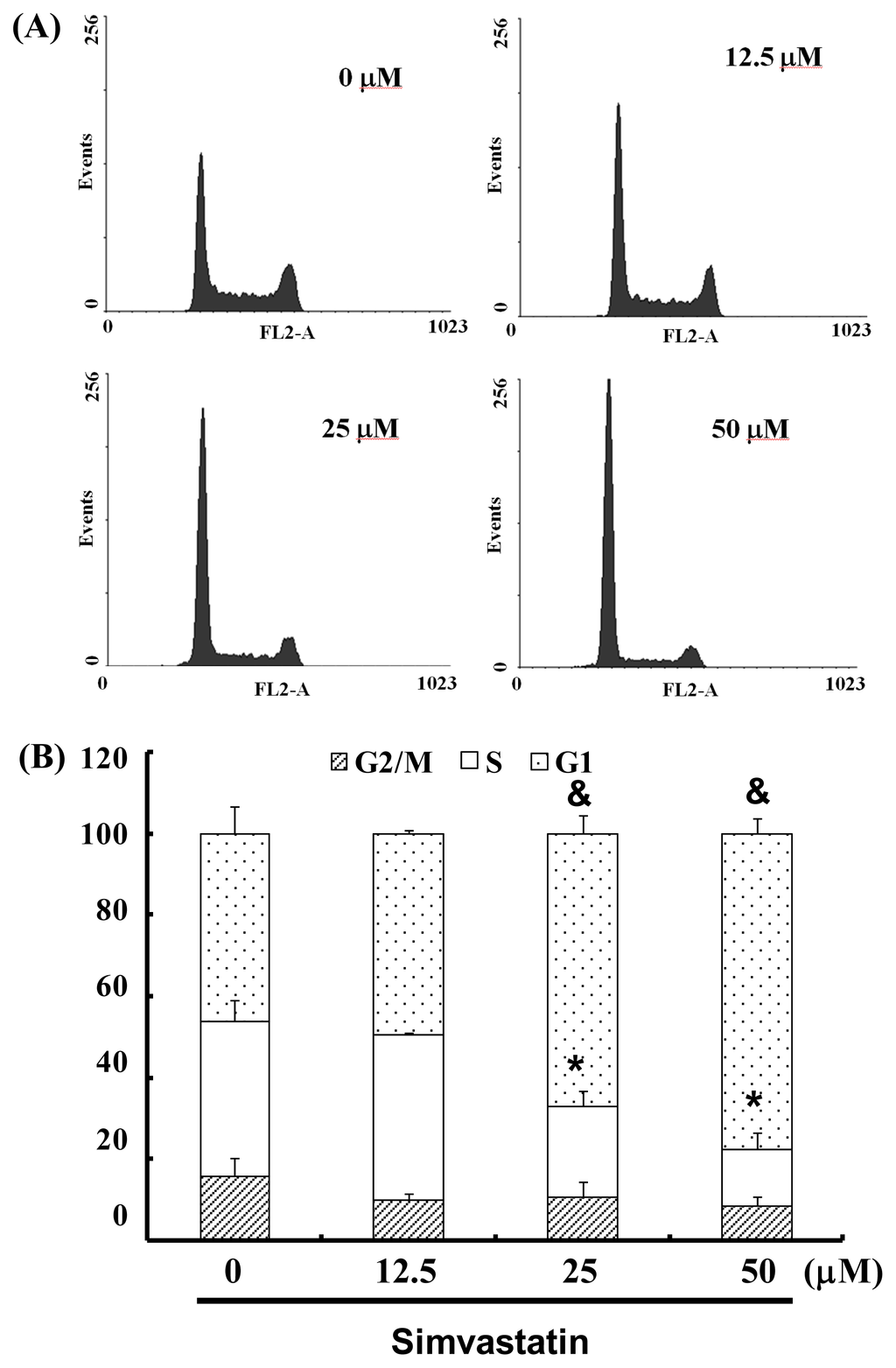
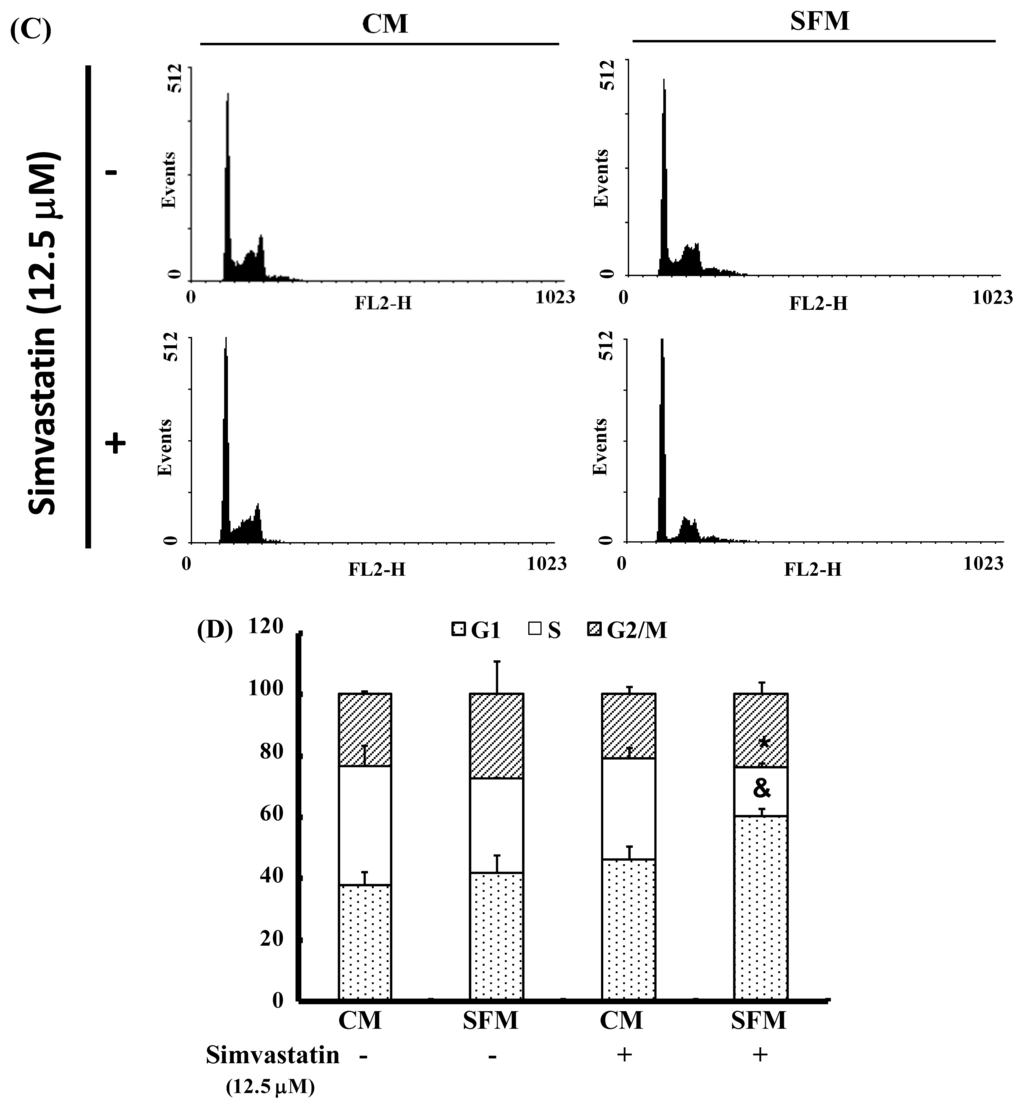
Figure 2.
Influence of SIM on cell cycle progression/distribution in NCI-H460 cells: (A) Cell cycle analysis of NCI-H460 cells after being cultured with SIM for 24 h; (B) SIM induced an increase in the number of cells in the G1 phase (%) of the cell cycle. (C) Cell cycle analysis of NCI-H460 cells after being cultured in complete medium (CM) or serum free medium (SFM) with SIM 12.5 μM. (D) SIM and SFM induced an increase in the number of cells in G1 phase (%). Symbols (* and &) in each group of bars indicates that the differences resulting from SIM treatment is statistically significant at p < 0.05.
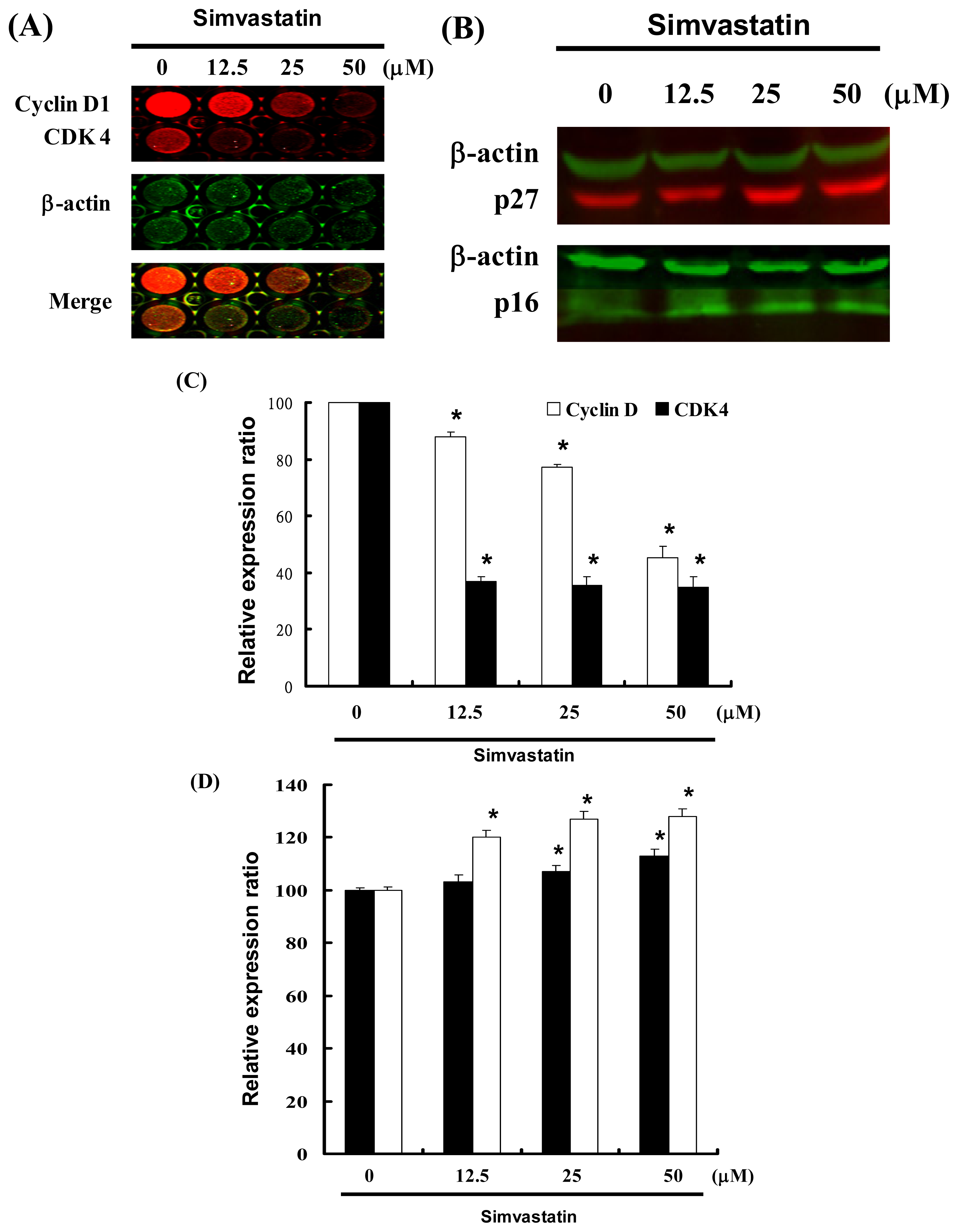
2.4. Cell Cycle Arrest Was Induced by SIM in NCI-H460 Cells via Cyclin D and Cdk4 Downregulation and Enhanced CDK Inhibition
Figure 3 illustrates a protein immunoblot analysis of SIM treated NCI-H460 cells. Cyclin D and Cdk4 protein expression was quantified by measuring the relative band intensities. Results revealed a decrease in Cyclin D and Cdk4 following incubation with SIM (Figure 3A). The CDK inhibitors, p16 and p27, were also quantified by measuring the relative band intensities (Figure 3B). Cyclin D and Cdk4 levels were significantly lower in cells incubated with SIM (Figure 3C). Results show that p16 and p27 levels were significantly upregulated in cells incubated with SIM (Figure 3D).
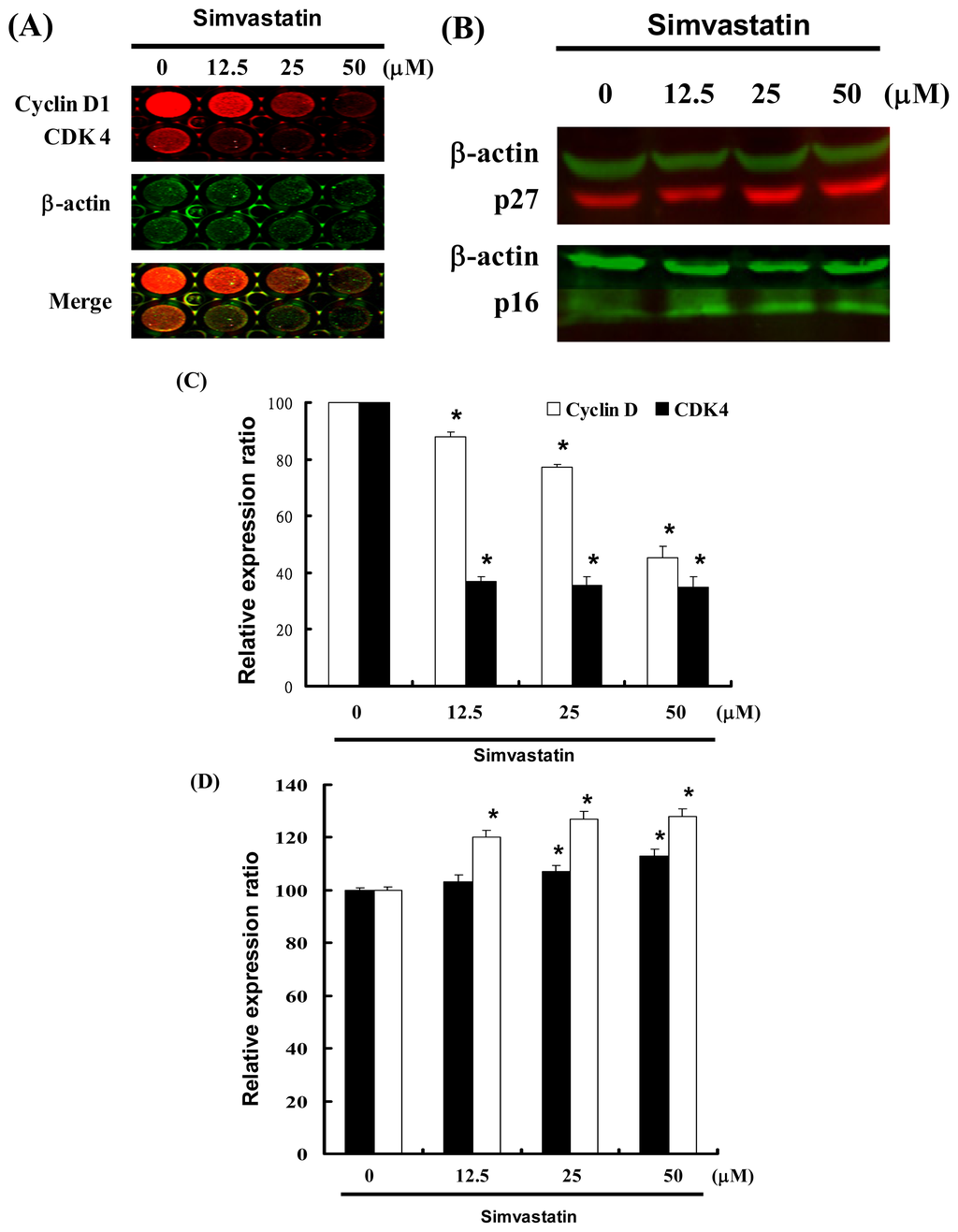
Figure 3.
Cell cycle arrest by SIM in NCI-H460 cells via inhibition of Cdk4 and Cyclin D, in which the effects of CDK inhibitors p16 and p27 were enhanced: Cells were treated with SIM followed by (A) In-cell Western blot analysis (B) Western blot analysis (C and D) Quantification of band intensities by Li-COR near infrared imaging system. Significant differences were determined at a level of * p < 0.05 versus the 0 μM control group.
These results indicate that SIM treatment increased the number of cells in the G1 phase via a downregulation of Cyclin D and Cdk4, as well as through the over expression of CDK inhibitors (p16 and p27).
HMG-CoA reductase inhibitors have demonstrated the ability to reduce albuminuria among patients with diabetes mellitus [15], transform growth factor-β1 (TGF-β1) expression [16], improve wound healing [17] and enhance endothelial function [18]. SIM is an HMG-CoA reductase inhibitor with a lipophilic compound that may provide more potent effects, such as chemo-prevention or chemo-therapy in extra-hepatic sites [19].
Cell cycle progression is regulated by CDKs and cyclin-dependent kinase inhibitors, the activity of which is largely controlled by their association with cyclins [20]. Analyses of the most common malignancies in humans have revealed mutations in these cell cycle regulators [21].
3. Materials and Methods
3.1. Materials
Simvastatin[1S,3R,7S,8S,8aR)-8-{2-[(2R,4R)-4-hydroxy-6-oxotetrahydro-2H-pyran-2-yl]ethyl}-3,7- dimethyl-1,2,3,7,8,8a-hexahydronaphthalen-1-yl 2,2-dimethylbutanoate] was obtained from Sigma (St. Louis, MO, USA). RPMI 1640 medium, fetal bovine serum, antibiotics, trypsin and phosphate-buffered saline (PBS) were purchased from Gibco, BRL (Grand Island, NY, USA). All other reagents and compounds were analytical grade.
3.2. Cell Culture
NCI-H460 cells were purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan). The cells were maintained on culture dishes, in RPMI 1640 medium supplemented with 10% (v/v) FBS and cultured in an incubator at 37 °C in an atmosphere containing 5% CO2.
3.3. Cell Proliferation Assay
Cells were seeded into 96-well culture plates at 10,000 cells/well and treated with SIM for 1–3 days. MTT dye (1 mg/mL) was added for an additional 4 h following treatment. The reaction was stopped by the addition of DMSO, whereupon OD540 was measured using a multi-well plate reader (Powerwave XS, Biotek, Seattle, WA, USA). In the absence of cells, the background absorbance of the medium was subtracted. The cell viability assays were determined by trypan blue analysis. Results were expressed as a percentage of the control, which was considered 100%. Each assay was performed in triplicate, and the results were expressed as the mean (±SEM).
3.4. Measurement of Apoptosis
NCI-H460 cells were first seeded in 6-well plates (Orange Scientific, E.U.). The cells were harvested after treatment with SIM for 4 h. The cells were then re-centrifuged (the supernatant was discarded) and resuspended/incubated for 15 min in 1× annexin-binding buffer, 5 μL of FITC annexin V (BD Pharmingen, San Jose, CA, USA) and 1 μL of 100 μg/mL PI working solution. Following incubation, the stained cells were analyzed using flow cytometry (FACSCalibur, San Jose, CA, USA). Data was analyzed using WinMDI 2.8 free software (BD, San Jose, CA, USA).
3.5. Cell Cycle Analysis
For cell cycle analysis, we employed the fluorescent nucleic acid dye, propidium iodide (PI), to identify the proportion of cells in each of the three interphase stages of the cell cycle. Cells were treated with SIM for 24 h and then harvested and fixed in 1 mL cold 70% ethanol for at least 8 h at −20 °C. DNA was stained in PI/RNaseA solution, and the DNA content was detected using flow cytometry. Data was analyzed using WinMDI 2.8 free software (BD, San Jose, CA, USA).
3.6. Western Blot Assay
A total of 30–50 μg of proteins were separated using 12% SDS-PAGE and transferred to PVDF membranes (Millipore, Billerica, MA, USA). The membranes were blocked with blocking buffer (Odyssey, Englewood, NJ, USA) overnight and subsequently incubated with anti-β-actin (Sigma-Aldrich, St. Louis, MO, USA), anti-p16 (Abcam, Cambridge, UK), anti-caspase-3 and anti-p27 (Santa Cruz BioTechnology, Santa Cruz, CA, USA) antibodies for 1.5–2 h. The blots were then washed and incubated with a secondary antibody (IRDye Li-COR, Lincoln, NE, USA) at a dilution of 1/20,000 for 30 min. The antigen was then visualized using a near infrared imaging system (Odyssey LI-COR, Lincoln, NE, USA) and data was analyzed using Odyssey 2.1 software.
In-cell Western blotting is an effective means to quantify intracellular signaling in whole cells. Briefly, this assay process involves seeding cells in microtiter plates, followed by fixation/permeabilization and subsequent labeling using anti-Cyclin D (Santa Cruz BioTechnology, Santa Cruz, CA, USA), Cdk4 (Abcam, Cambridge, UK), β-actin (Sigma-Aldrich, St. Louis, MO, USA) and infrared-conjugated secondary antibodies. The Odyssey near infrared imaging system obtains well-level data.
3.7. Statistical Analysis
All data was reported as the mean (±SEM) of at least three separate experiments. A t-test or one-way ANOVA with post-hoc test was employed for statistical analysis, with significant differences determined as p < 0.05.
4. Conclusions
In the current study, SIM induced G1 cell cycle arrest in NCI-H460 cells, indicating the existence of an additional mechanism by which SIM may inhibit the proliferation of NSCLC cells. To analyze the biological mechanisms underlying SIM-induced G1 cell cycle arrest, we evaluated the protein and gene expressions of CDKs and cyclins, as well as CDK-inhibitors p16 and p27. The SIM-induced reduction of the number of cells in S phase and G1 cell cycle arrest was associated with a marked reduction in Cyclin D-Cdk4 protein levels. Our results suggest that SIM-induced G1 cell cycle arrest in NCI-H460 cells is mediated by the upregulation of p16 and p27 proteins, which in turn indicates that the cell cycle-suppressing activity of SIM is associated with its anti-cancer properties.
References
- Ou, J.P.; Lin, H.Y.; Su, K.Y.; Yu, S.L.; Tseng, I.H.; Chen, C.J.; Hsu, H.C.; Chan, D.C.; Sophia Chen, Y.L. Potential therapeutic role of Z-isochaihulactone in lung cancer through induction of apoptosis via notch signaling. Evid. Based Complement. Alternat. Med 2012, 2012. [Google Scholar] [CrossRef]
- Chien, C.R.; Shih, Y.C. Economic evaluation of bevacizumab in the treatment of non-small cell lung cancer (NSCLC). Clinicoecon. Outcomes Res 2012, 4, 201–208. [Google Scholar]
- Kah, J.; Wüstenberg, A.; Keller, A.D.; Sirma, H.; Montalbano, R.; Ocker, M.; Volz, T.; Dandri, M.; Tiegs, G.; Sass, G. Selective induction of apoptosis by HMG-CoA reductase inhibitors in hepatoma cells and dependence on p53 expression. Oncol. Rep 2012, 28, 1077–1083. [Google Scholar]
- Alexandre, L.; Clark, A.B.; Cheong, E.; Lewis, M.P.; Hart, A.R. Systematic review: Potential preventive effects of statins against oesophageal adenocarcinoma. Aliment. Pharmacol. Ther 2012, 36, 301–311. [Google Scholar]
- Broughton, T.; Sington, J.; Beales, I.L. Statin use is associated with a reduced incidence of colorectal cancer: A colonoscopy-controlled case-control study. BMC Gastroenterol 2012, 12, 36. [Google Scholar]
- Papadopoulos, G.; Delakas, D.; Nakopoulou, L.; Kassimatis, T. Statins and prostate cancer: Molecular and clinical aspects. Eur. J. Cancer 2011, 47, 819–830. [Google Scholar]
- Zhou, P.; Cheng, S.W.; Yang, R.; Wang, B.; Liu, J. Combination chemoprevention: Future direction of colorectal cancer prevention. Eur. J. Cancer Prev 2012, 21, 231–240. [Google Scholar]
- Piechota-Polanczyk, A.; Goraca, A.; Demyanets, S.; Mittlboeck, M.; Domenig, C.; Neumayer, C.; Wojta, J.; Nanobachvili, J.; Huk, I.; Klinger, M. Simvastatin decreases free radicals formation in the human abdominal aortic aneurysm wall via NF-κB. Eur. J. Vasc. Endovasc. Surg 2012, 44, 133–137. [Google Scholar]
- Tu, Y.S.; Kang, X.L.; Zhou, J.G.; Lv, X.F.; Tang, Y.B.; Guan, Y.Y. Involvement of Chk1-Cdc25A-cyclin A/CDK2 pathway in simvastatin induced S-phase cell cycle arrest and apoptosis in multiple myeloma cells. Eur. J. Pharmacol 2011, 670, 356–364. [Google Scholar]
- Higgins, M.J.; Prowell, T.M.; Blackford, A.L.; Byrne, C.; Khouri, N.F.; Slater, S.A.; Jeter, S.C.; Armstrong, D.K.; Davidson, N.E.; Emens, L.A.; et al. A short-term biomarker modulation study of simvastatin in women at increased risk of a new breast cancer. Breast Cancer Res. Treat 2012, 131, 915–924. [Google Scholar]
- Podhorecka, M.; Halicka, D.; Klimek, P.; Kowal, M.; Chocholska, S.; Dmoszynska, A. Simvastatin and purine analogs have a synergic effect on apoptosis of chronic lymphocytic leukemia cells. Ann. Hematol 2010, 89, 1115–1124. [Google Scholar]
- Kochuparambil, S.T.; Al-Husein, B.; Goc, A.; Soliman, S.; Somanath, P.R. Anticancer efficacy of simvastatin on prostate cancer cells and tumor xenografts is associated with inhibition of Akt and reduced prostate-specific antigen expression. J. Pharmacol. Exp. Ther 2011, 336, 496–505. [Google Scholar]
- Bababeygy, S.R.; Polevaya, N.V.; Youssef, S.; Sun, A.; Xiong, A.; Prugpichailers, T.; Veeravagu, A.; Hou, L.C.; Steinman, L.; Tse, V. HMG-CoA reductase inhibition causes increased necrosis and apoptosis in an in vivo mouse glioblastoma multiforme model. Anticancer Res 2009, 29, 4901–4908. [Google Scholar]
- Polo, M.P.; Crespo, R.; de Bravo, M.G. Geraniol and simvastatin show a synergistic effect on a human hepatocarcinoma cell line. Cell Biochem. Funct 2011, 29, 452–458. [Google Scholar]
- Ruggenenti, P.; Cattaneo, D.; Rota, S.; Iliev, I.; Parvanova, A.; Diadei, O.; Ene-Iordache, B.; Ferrari, S.; Bossi, A.C.; Trevisan, R.; et al. Effects of combined ezetimibe and simvastatin therapy as compared with simvastatin alone in patients with type 2 diabetes: A prospective randomized double-blind clinical trial. Diabetes Care 2010, 33, 1954–1956. [Google Scholar]
- Schaafsma, D.; McNeill, K.D.; Mutawe, M.M.; Ghavami, S.; Unruh, H.; Jacques, E.; Laviolette, M.; Chakir, J.; Halayko, A.J. Simvastatin inhibits TGFβ1-induced fibronectin in human airway fibroblasts. Respir. Res 2011, 12, 113. [Google Scholar]
- Karadeniz Cakmak, G.; Irkorucu, O.; Ucan, B.H.; Emre, A.U.; Bahadir, B.; Demirtas, C.; Tascilar, O.; Karakaya, K.; Acikgoz, S.; et al. Simvastatin improves wound strength after intestinal anastomosis in the rat. J. Gastrointest. Surg 2009, 13, 1707–1716. [Google Scholar]
- Lei, J.; Gu, X.; Ye, Z.; Shi, J.; Zheng, X. Antiaging effects of simvastatin on vascular endothelial cells. Clin. Appl. Thromb. Hemost 2012. [Google Scholar] [CrossRef]
- Murtaza, G. Solubility enhancement of simvastatin: A review. Acta Pol. Pharm 2012, 69, 581–590. [Google Scholar]
- Chen, M.J.; Tang, W.Y.; Hsu, C.W.; Tsai, Y.T.; Wu, J.F.; Lin, C.W.; Cheng, Y.M.; Hsu, Y.C. Apoptosis induction in primary human colorectal cancer cell lines and retarded tumor growth in SCID mice by sulforaphane. Evid. Based Complement. Alternat. Med 2012, 2012, 415231. [Google Scholar]
- Kim, Y.R.; Byun, H.S.; Jeon, J.; Choi, B.L.; Park, K.A.; Won, M.; Zhang, T.; Shin, S.; Lee, H.; Oh, J. Apoptosis signal-regulating kinase1 is inducible by protein kinase Cδ and contributes to phorbol ester-mediated G1 phase arrest through persistent JNK activation. Cell Biochem. Biophys 2011, 61, 199–207. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).