Role of UPR Pathway in Defense Response of Aedes aegypti against Cry11Aa Toxin from Bacillus thuringiensis




Abstract
:1. Introduction
2. Results
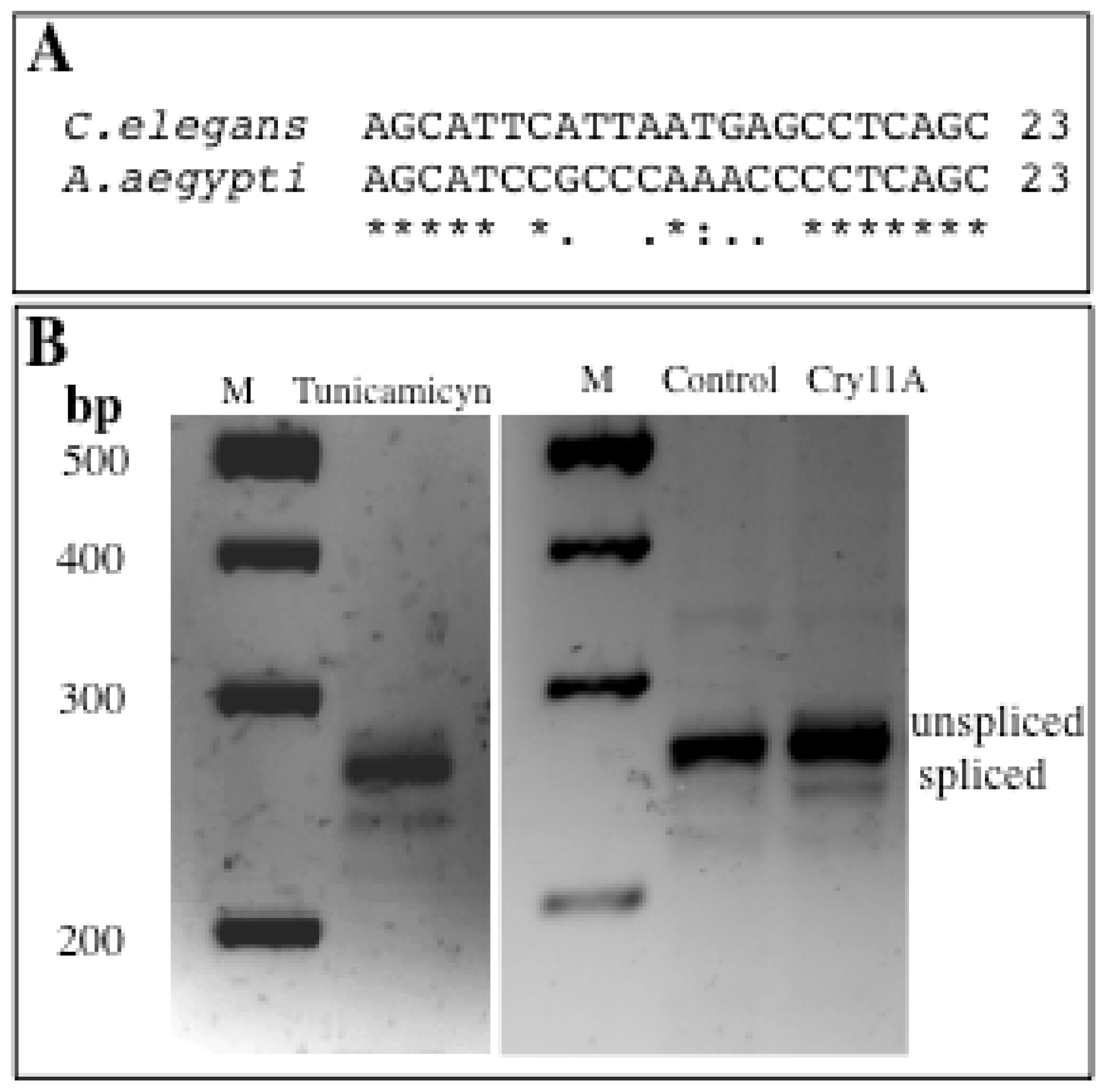
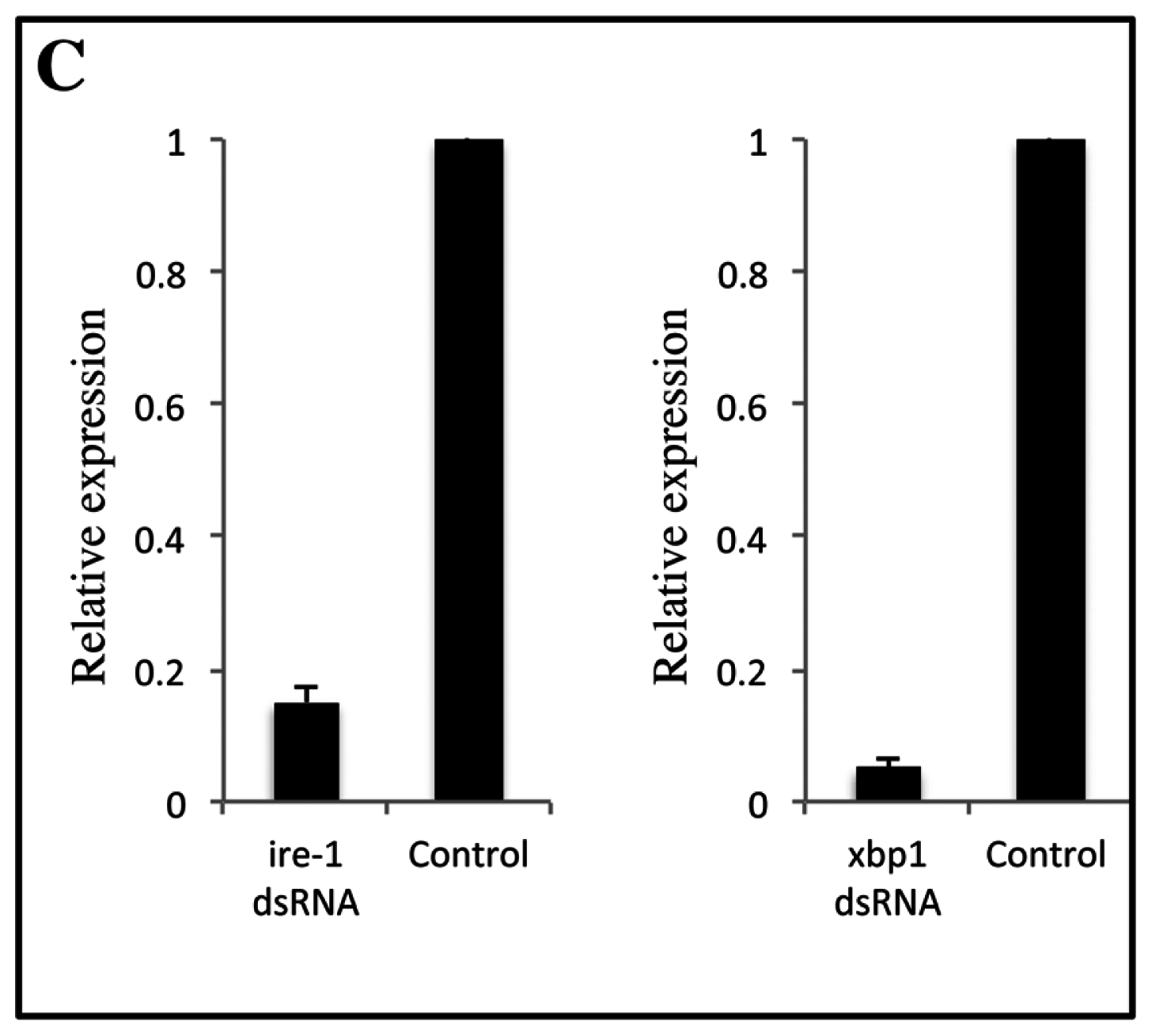
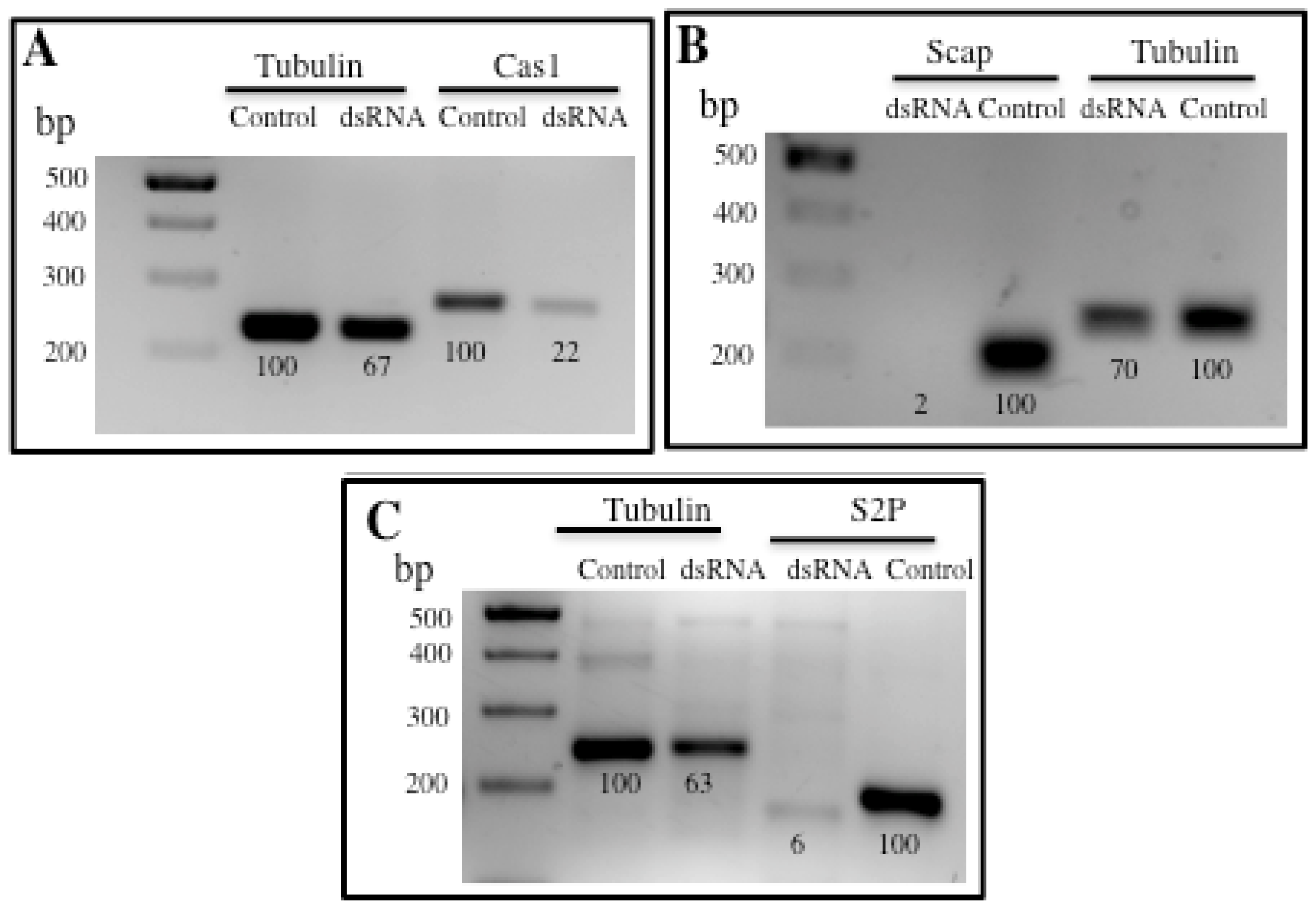
2.1. Role of IRE-1 UPR Pathway in Response to Cry11Aa in A. aegypti
2.2. Role of SREBP Pathway in Response to Cry11Aa in A. aegypti
3. Discussion
4. Experimental Section
4.1. Production of Cry11Aa Crystal Inclusions
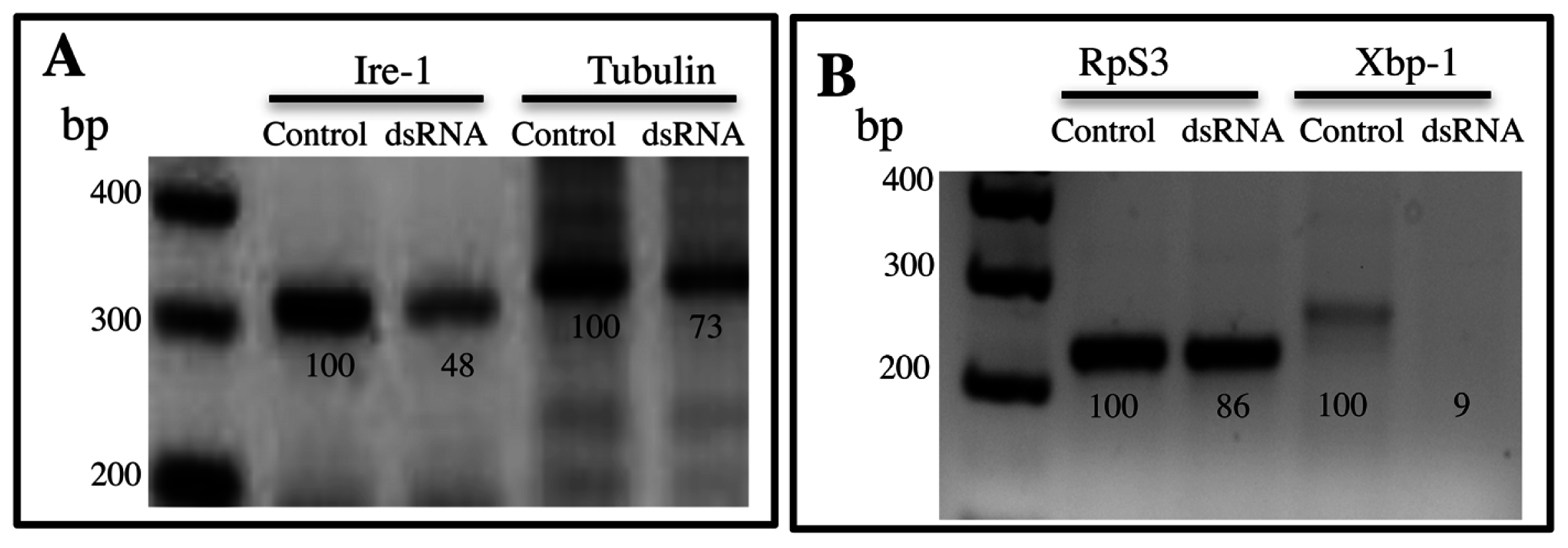
4.2. RNA Interference (RNAi) Assays
4.3. RT-PCR and Quantitative Real Time PCR
4.4. Insect Bioassays
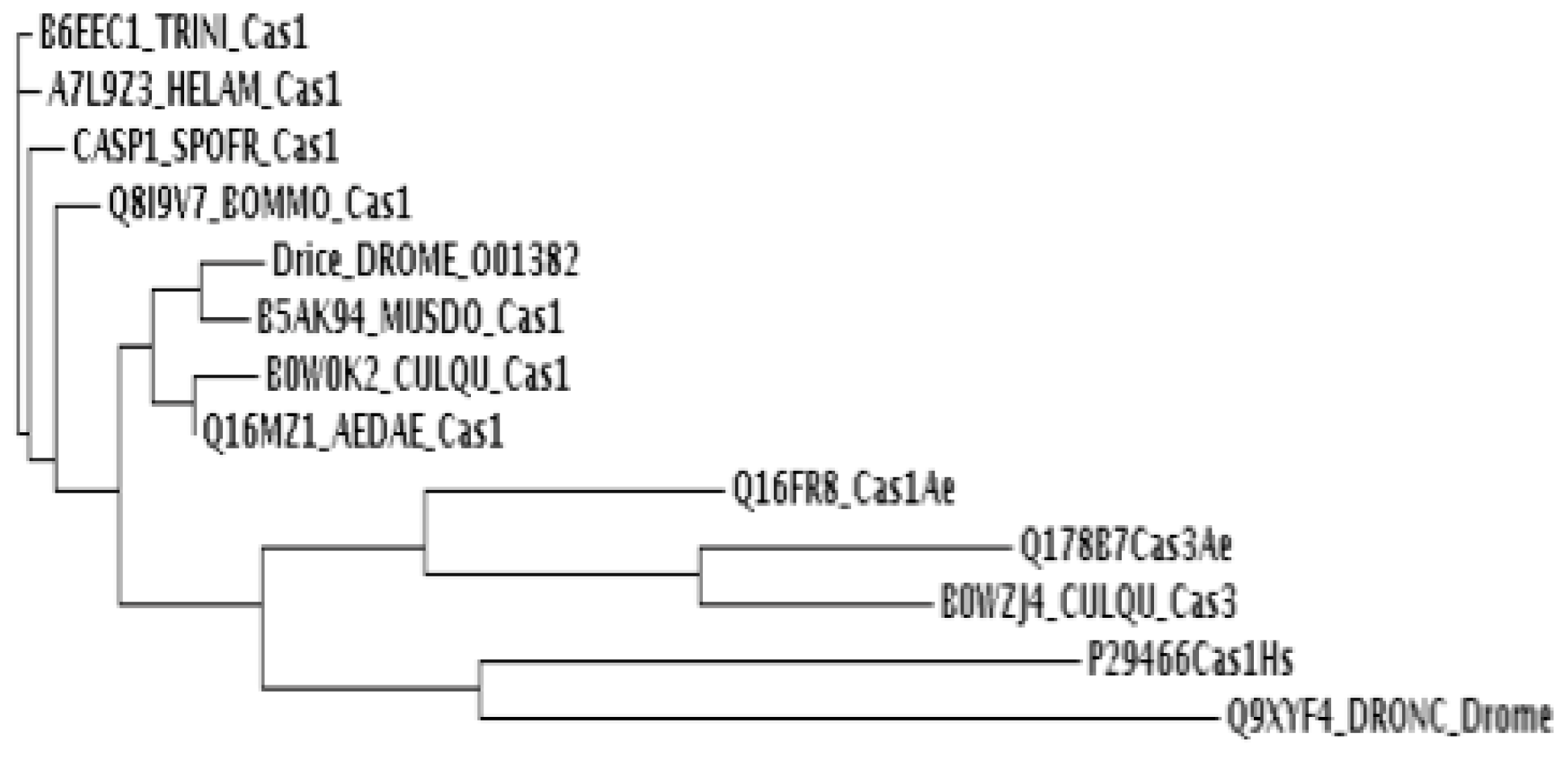
4.5. Phylogenetic Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev 2013, 37, 3–22. [Google Scholar]
- James, C. Global Status of Commercialized Biotech/GM Crops 2011; The International Service for the Acquisition of Agri-biotech Applications (ISAAA): Metro Manila, Philippines, 2011. [Google Scholar]
- Aroian, R.; van der Goot, F.G. Pore forming toxins and cellular non-immune defenses (CNIDs). Curr. Opin. Microbiol 2007, 10, 57–61. [Google Scholar]
- Cancino-Rodezno, A.; Alexander, C.; Villaseñor, R.; Pacheco, S.; Porta, H.; Pauchet, Y.; Gill, S.S.; Soberón, M.; Bravo, A. The mitogen-activated protein kinase p38 is involved in insect defense against Cry toxins from Bacillus thuringiensis. Insect. Biochem. Mol. Biol 2010, 40, 58–63. [Google Scholar]
- Bischof, L.J.; Kao, Ch-Y.; Los, F.C.O.; Gonzalez, M.R.; Shen, Z.; Briggs, S.P.; van der Goot, F.G.; Aroian, RV. Activation of the unfolded protein response is required for defenses against bacterial pore forming toxin in vivo. PLoS Pathog 2008, 4, e1000176. [Google Scholar]
- Kao, Ch.-Y.; Los, F.C.O.; Huffman, D.L.; Wachi, S.; Kloft, N.; Husmann, M.; Karabrahimi, V.; Schwartz, J.-L.; Bellier, A.; Ha, Ch.; et al. Global functional analyses of cellular responses to pore-forming toxins. PLoS Pathog. 2011, 7, e1001314. [Google Scholar]
- Gurcel, L.; Abrami, L.; Girardin, S.; Tschopp, J.; van der Goot, F.G. Caspase-1 Activation of lipid metabolic pathways in response to bacterial pore-forming toxins promotes cell survival. Cell 2006, 126, 1135–1145. [Google Scholar]
- Cancino-Rodezno, A.; Porta, H.; Soberón, M.; Bravo, A. Defense and death responses to pore forming toxins. Biotechnol. Gen. Eng. Rev 2009, 26, 65–94. [Google Scholar]
- Gonzalez, M.R.; Bischofberger, M.; Freche, B.; Ho, S.; Parton, R.G.; van der Goot, F.G. Pore forming toxins induce multiple cellular responses promoting survival. Cell Microbiol 2011, 13, 1026–1043. [Google Scholar]
- Gutierrez, M.G.; Saka, H.A.; Chinen, I.; Zoppino, F.C.M.; Yoshimori, T.; Bocco, J.L.; Colombo, M.I. Protective role of autophagy against Vibrio cholerae cytolysin, a pore-forming toxin from V. cholerae. Proc. Natl. Acad. Sci. USA 2007, 104, 1829–1834. [Google Scholar]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar]
- Brown, M.S.; Goldstein, J.L. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar]
- Calfon, M.; Zeng, H.; Urano, F.; Till, J.H.; Hubbard, S.R.; Harding, H.P.; Clark, S.G.; Ron, D. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature 2002, 415, 92–96. [Google Scholar]
- Amarneh, B.; Matthews, K.A.; Rawson, R.B. Activation of Sterol regulatory element-binding protein by the caspase Drice in Drosophila larvae. J. Biol. Chem 2009, 284, 9674–9682. [Google Scholar]
- Mori, K. Signaling pathways in the unfolded protein response: Development from yeast to mammals. J. Biochem 2009, 146, 743–750. [Google Scholar]
- Reimold, A.M.; Iwakoshi, N.N.; Manis, J.; Vallabhajosyula, P.; Szomolanyi-Tsuda, E.; Gravallese, E.M.; Friend, D.; Grusby, M.J.; Alt, F.; Glimcher, L.H. Plasma cell differentiation requires the transcription factor XBP-1. Nature 2001, 412, 300–307. [Google Scholar]
- Sriburi, R.; Jackowski, S.; Mori, K.; Brewer, J.W. XBP1: A link between unfolded protein response, lipid biosynthesis and biogenesis of the endoplasmatic reticulum. J. Cell Biol 2004, 167, 35–41. [Google Scholar]
- Theopold, U.; Ekengren, S.; Hultmark, D. HLH106, a Drosophila transcription factor with similarity to the vertebrate sterol responsive element binding protein. Proc. Natl. Acad. Sci. USA 1996, 93, 1195–1199. [Google Scholar]
- Seegmiller, A.C.; Dobrostskaya, I.; Goldstein, J.L.; Ho, Y.K.; Brown, M.S.; Rawson, R.B. The SREBP pathway in Drosophila: Regulation by palmitate, not sterols. Dev. Cell 2002, 2, 229–238. [Google Scholar]
- Kunte, A.S.; Matthews, K.A.; Rawson, R.B. Fatty acid auxotrophy in Drosophila larvae lacking SREBP. Cell Metab 2006, 3, 439–448. [Google Scholar]
- Matthews, K.A.; Ozdemir, C.; Rawson, R.B. Activation of sterol regulatory element binding proteins in the absence of Scap in Drosophila melanogaster. Genetics 2010, 185, 189–198. [Google Scholar]
- Chang, C.; Yu, Y.M.; Dai, S.M.; Law, S.K.; Gill, S.S. High-level cryIVD and cytA gene expression in Bacillus thuringiensis does not require the 20-kilodalton protein, and the coexpressed gene products are synergistic in their toxicity to mosquitoes. Appl. Environ. Microbiol 1993, 59, 815–821. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acid Res 2004, 32, 1792–1797. [Google Scholar]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol 2010, 59, 307–321. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Silenced protein | Larval survival % b | LC50 value (95% fiducial limits) | Fold increase in susceptibility |
|---|---|---|---|
| Control | 95 | 555 (408–725) | - |
| IRE-1 | 80 | 214 (137–306) | 2.6 |
| XBP-1 | 83 | 176.4 (127.1–222.6) | 3.1 |
| CASP1(Q16MZ1) | 98 | 392.7 (230.6–542.4) | NS a |
| SCAP | 6 | 683 (475–1160) | NS |
| S2P | 87 | 355 (137–533) | NS |
| Protein (Accession number) | Percentage of identity (Accession number) | ||
|---|---|---|---|
| Aedes aegypti | Culex quinquefasciatus | Anopheles gambiae | Drosophila melanogaster |
| Ire-1 (XM_001655187.1) | 77% (XP_001843113) | 61% (XP_562694) | 55% (NP_001097839) |
| Xbp-1 (XM_001651044.1) | 66% (XP-001847153) | 61% (XP-310116) | 46% (NP-524722) |
| Cas-1 (XM_001655826.1) | 94% (XP-001842236) | 79% (XP-316795) | 74% (CAA72937) |
| Scap (XM_001651241.1) | 89% (XP_001863686) | 74% (XP_309314) | 54% (AAM20923) |
| S2P (XM_001663618.1) | 84% (XP_001663668) | 68% (XP_320696) | 46% (NP_610705) |
| Gene | Oligonucleotide sequence | PCR product size (bp) |
|---|---|---|
| S2P | 5′-CCG GAA TTC AAC ATT CGG AAG GTG ATT GA-3′ 5′-CCC AAG CTT GGT GGC CAA TGT AGA TAA CG-3′ | 172 |
| SCAP | 5′-CCG GAA TTC GTG GGA TAA GTC GTT CGA TG-3′ 5′-CCC AAG CTT TCA TGA AGC CTC TTT GGA AG-3′ | 192 |
| CASP1Q16 | 5′-CCG GAA TTC TAT CTG TAT GCA AAG GA-3′ 5′-CCC AAG CTT ATG AGT AGA ATC CCG GAA TG-3′ | 268 |
| XBP-1 | 5′-CCG GAA TTC TCA ACG ATC TTC AGC AGC AC-3′ 5′-CCC AAG CTT TGT AGA GCA GGC AGA GAG CA-3′ | 264 |
| IRE-1 | 5′-CCG GAA TTC TGC TGT TGC AAA AGA TGA GG-3′ 5′-CCC AAG CTT CTC AGG ATT CCG GTA CGT GT- 3′ | 220 |
| TUBULIN | 5′-CTA CGG CAA GAA GTC CAA GC-3′ 5′-GAA GCG GTG ATC GAA GAG AC-3′ | 243 |
| RPS3 | 5′-TTC TCG GCG TAC AGC TCG ACG-3′ 5′-GGC ATG TTC CGT GCT GAA TTG AAC G-3′ | 239 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bedoya-Pérez, L.P.; Cancino-Rodezno, A.; Flores-Escobar, B.; Soberón, M.; Bravo, A. Role of UPR Pathway in Defense Response of Aedes aegypti against Cry11Aa Toxin from Bacillus thuringiensis. Int. J. Mol. Sci. 2013, 14, 8467-8478. https://doi.org/10.3390/ijms14048467
Bedoya-Pérez LP, Cancino-Rodezno A, Flores-Escobar B, Soberón M, Bravo A. Role of UPR Pathway in Defense Response of Aedes aegypti against Cry11Aa Toxin from Bacillus thuringiensis. International Journal of Molecular Sciences. 2013; 14(4):8467-8478. https://doi.org/10.3390/ijms14048467
Chicago/Turabian StyleBedoya-Pérez, Leidy P., Angeles Cancino-Rodezno, Biviana Flores-Escobar, Mario Soberón, and Alejandra Bravo. 2013. "Role of UPR Pathway in Defense Response of Aedes aegypti against Cry11Aa Toxin from Bacillus thuringiensis" International Journal of Molecular Sciences 14, no. 4: 8467-8478. https://doi.org/10.3390/ijms14048467
APA StyleBedoya-Pérez, L. P., Cancino-Rodezno, A., Flores-Escobar, B., Soberón, M., & Bravo, A. (2013). Role of UPR Pathway in Defense Response of Aedes aegypti against Cry11Aa Toxin from Bacillus thuringiensis. International Journal of Molecular Sciences, 14(4), 8467-8478. https://doi.org/10.3390/ijms14048467