Off-Target Effect of Endogenous siRNA Derived from RMRP in Human Cells

Abstract
:1. Introduction
2. Results and Discussion
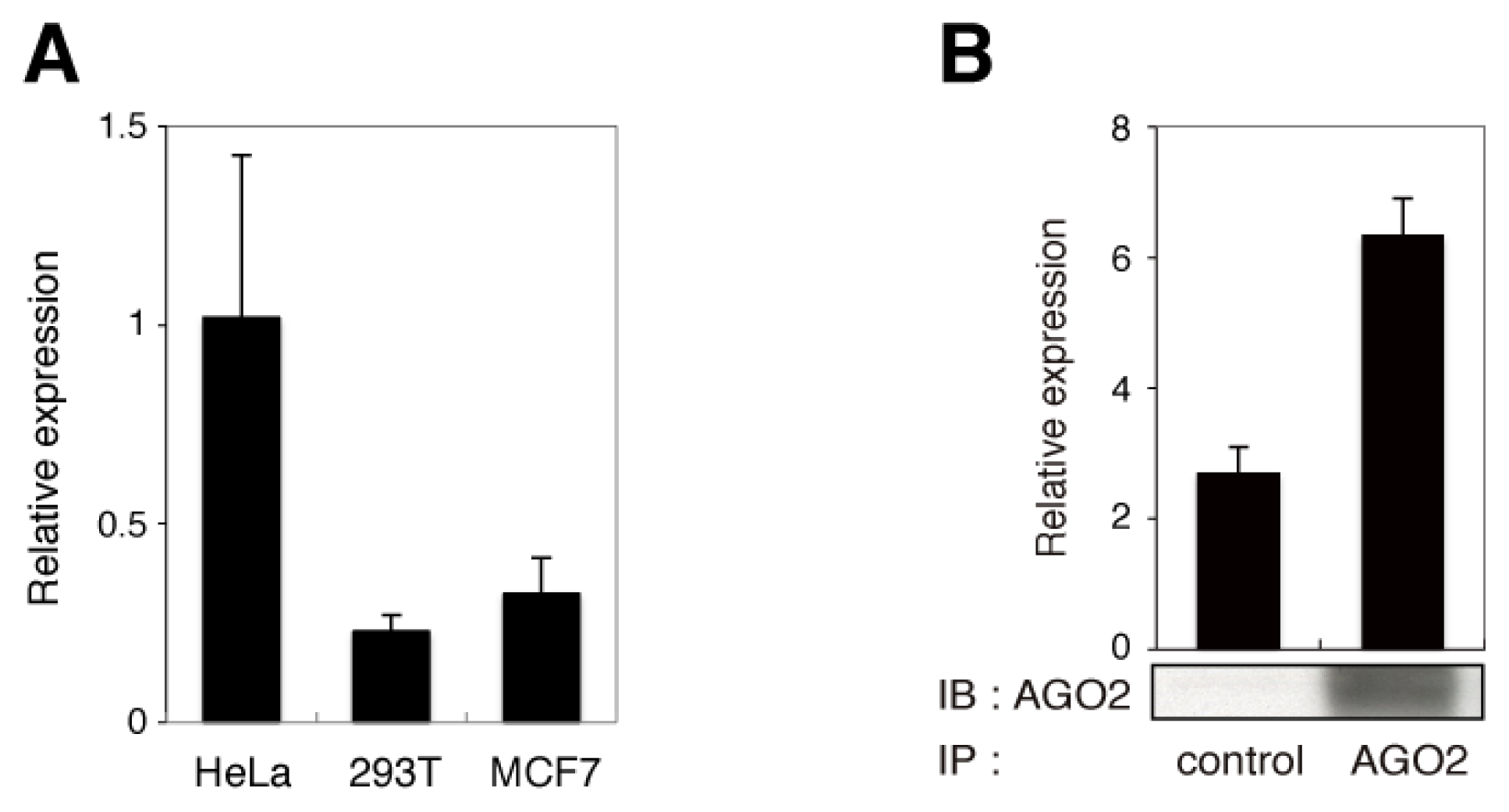
2.1. The Endo-siRNA Sequence Originates from Nucleotides 22–43 of RMRP
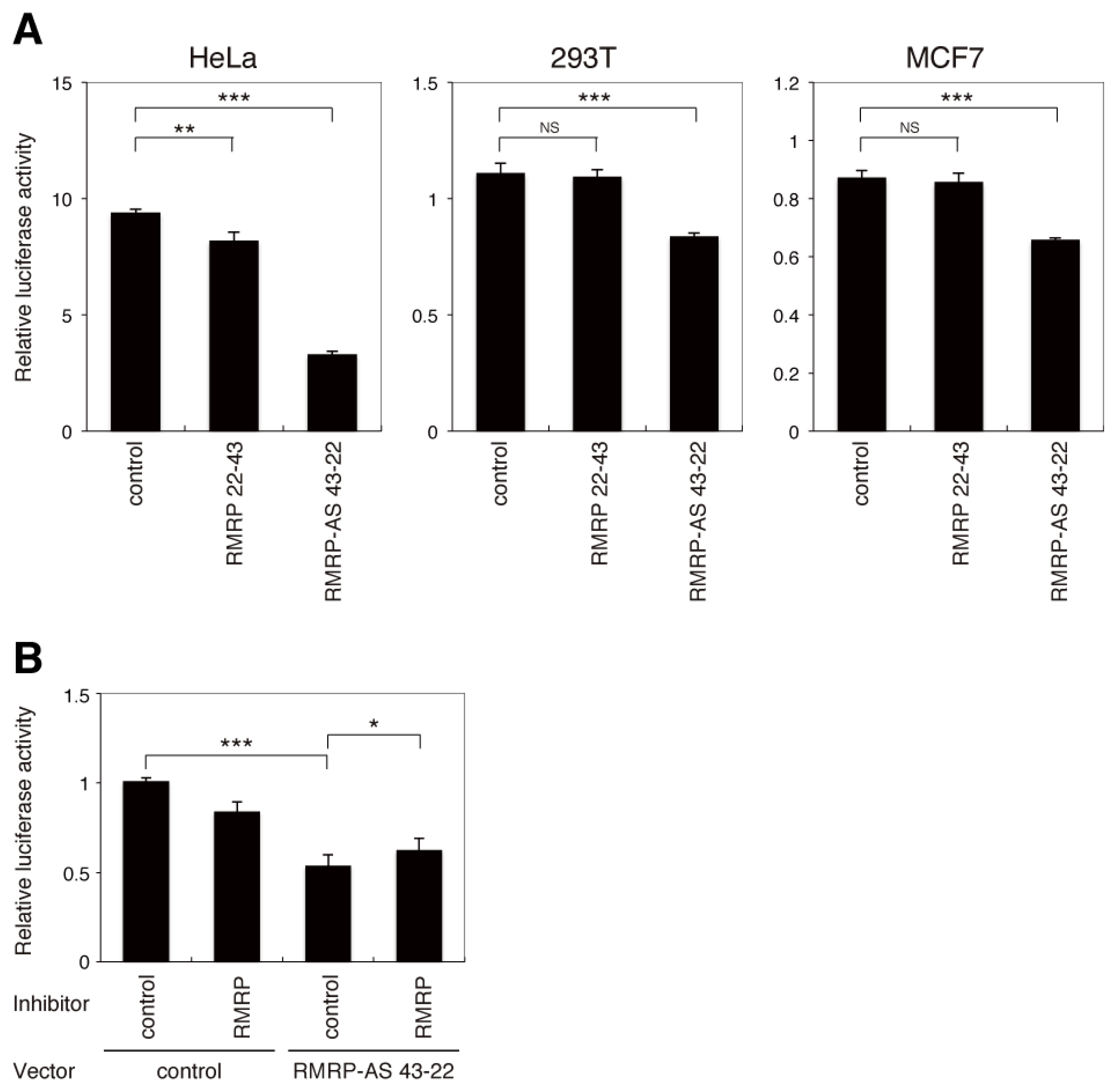
2.2. The Endo-siRNA Derived from RMRP Mediates Gene Silencing
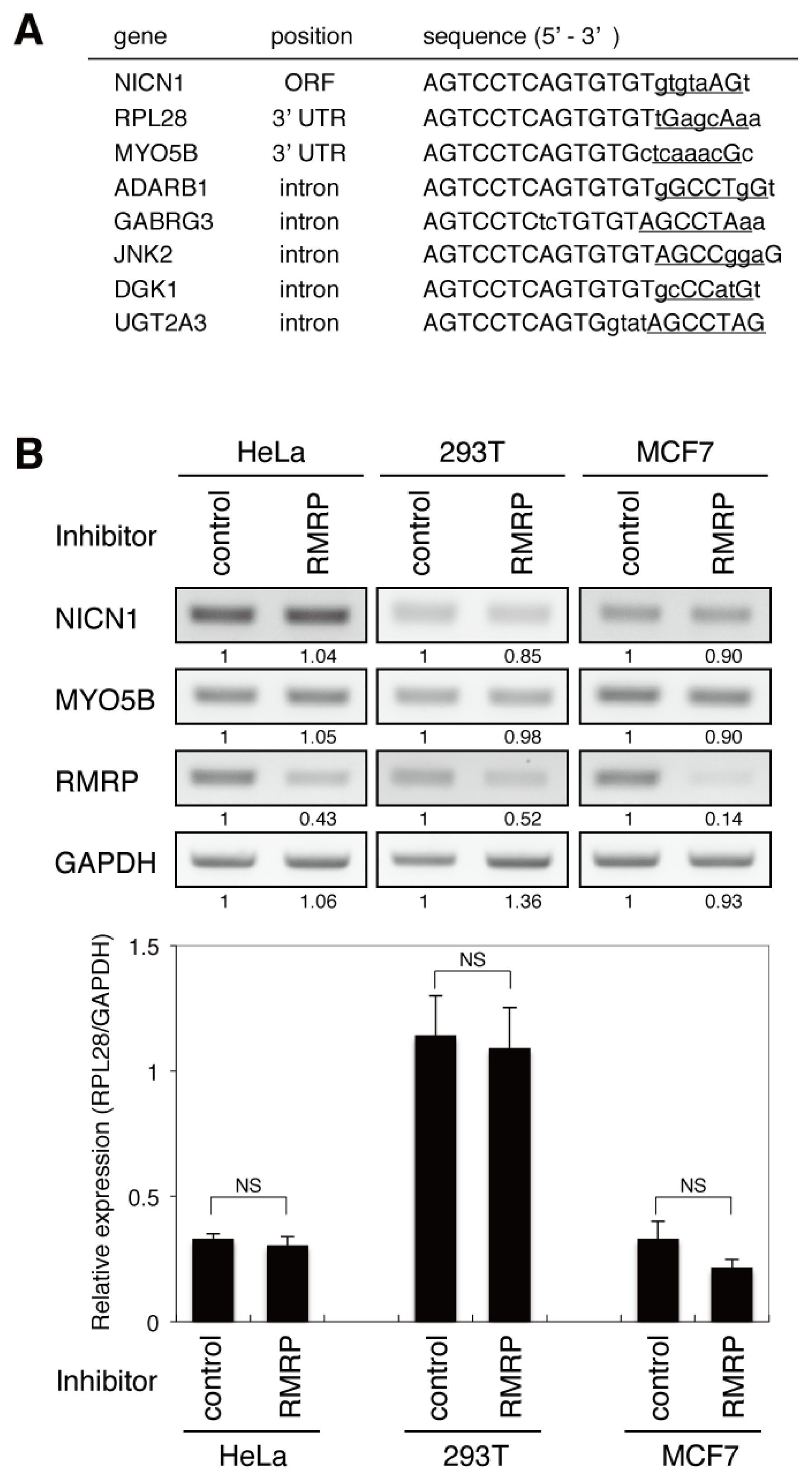
2.3. Overall Sequence Analogy Does Not Accurately Predict Unintended Targets of the Endo-siRNA Derived from RMRP
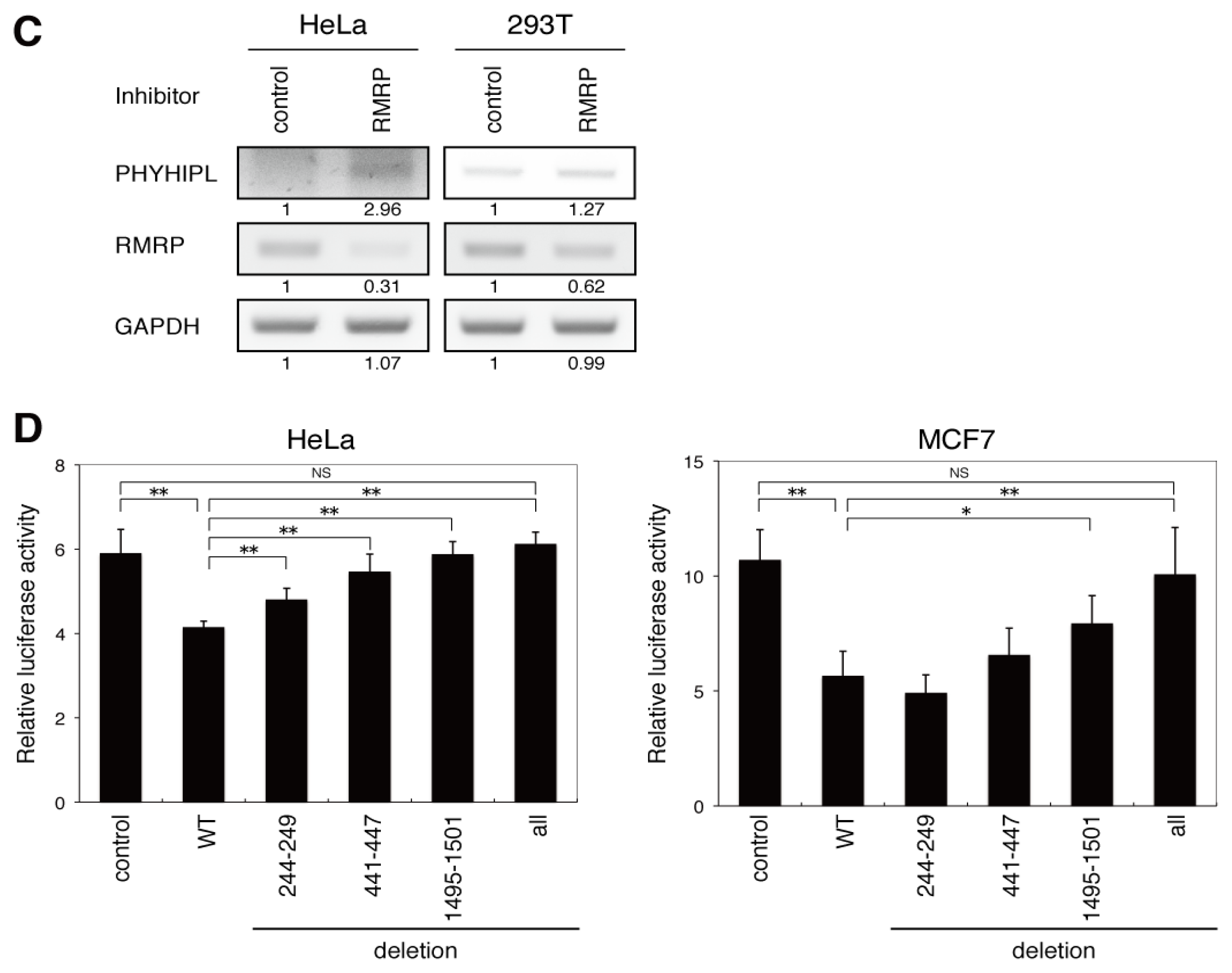
2.4. The Endo-siRNA Derived from RMRP Mediates Off-Targeting by a Mechanism Similar to That of miRNAs
3. Experimental Section
3.1. Cell Culture
3.2. Plasmids
3.3. Immunoprecipitation of Human AGO2 Complexes
3.4. Sequence-Specific Inhibition of the RMRP-Derived Endo-siRNA
3.5. RT-PCR and qRT–PCR
3.6. Luciferase Assay
3.7. Statistics
4. Conclusions
Supplementary Information
ijms-14-09305-s001.pdfAcknowledgments
Conflict of Interest
References
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet 2009, 10, 94–108. [Google Scholar]
- Siomi, H.; Siomi, M.C. On the road to reading the RNA-interference code. Nature 2009, 457, 396–404. [Google Scholar]
- Maida, Y.; Yasukawa, M.; Furuuchi, M.; Lassmann, T.; Possemato, R.; Okamoto, N.; Kasim, V.; Hayashizaki, Y.; Hahn, W.C.; Masutomi, K. An RNA-dependent RNA polymerase formed by TERT and the RMRP RNA. Nature 2009, 461, 230–235. [Google Scholar]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet 2008, 9, 102–114. [Google Scholar]
- Jackson, A.L.; Linsley, P.S. Recognizing and avoiding siRNA off-target effects for target identification and therapeutic application. Nat. Rev. Drug Discov 2010, 9, 57–67. [Google Scholar]
- Jackson, A.L.; Bartz, S.R.; Schelter, J.; Kobayashi, S.V.; Burchard, J.; Mao, M.; Li, B.; Cavet, G.; Linsley, P.S. Expression profiling reveals off-target gene regulation by RNAi. Nat. Biotechnol 2003, 21, 635–637. [Google Scholar]
- Birmingham, A.; Anderson, E.; Sullivan, K.; Reynolds, A.; Boese, Q.; Leake, D.; Karpilow, J.; Khvorova, A. A protocol for designing siRNAs with high functionality and specificity. Nat. Protoc 2007, 2, 2068–2078. [Google Scholar]
- Jackson, A.L.; Burchard, J.; Schelter, J.; Chau, B.N.; Cleary, M.; Lim, L.; Linsley, P.S. Widespread siRNA “Off-target” transcript silencing mediated by seed region sequence complementarity. RNA 2006, 12, 1179–1187. [Google Scholar]
- Birmingham, A.; Anderson, E.M.; Reynolds, A.; Ilsley-Tyree, D.; Leake, D.; Fedorov, Y.; Baskerville, S.; Maksimova, E.; Robinson, K.; Karpilow, J.; et al. 3′ UTR seed matches, but not overall identity, are associated with RNAi off-targets. Nat. Methods 2006, 3, 199–204. [Google Scholar]
- Doench, J.G.; Petersen, C.P.; Sharp, P.A. siRNAs can function as miRNAs. Genes Dev 2003, 17, 438–442. [Google Scholar]
- Grimson, A.; Farh, K.K.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA targeting specificity in mammals: Determinants beyond seed pairing. Mol. Cell 2007, 27, 91–105. [Google Scholar]
- Nielsen, C.B.; Shomron, N.; Sandberg, R.; Hornstein, E.; Kitzman, J.; Burge, C.B. Determinants of targeting by endogenous and exogenous microRNAs and siRNAs. RNA 2007, 13, 1894–1910. [Google Scholar]
- Tan Gana, N.H.; Victoriano, A.F.; Okamoto, T. Evaluation of online miRNA resources for biomedical applications. Genes Cells 2012, 17, 11–27. [Google Scholar]
- DIANA-microT-CDS v.4. Available online: http://www.microrna.gr/microT-CDS/ (accessed on 30 September 2011).
- TargetRank. Available online: http://genes.mit.edu/targetrank/ (accessed on 25 November 2011).
- TargetScanHuman, Release 5.2. Available online: http://www.targetscan.org/ (accessed on 30 September 2011).
- microRNA.org-Targets and Expression. Available online: http://www.microrna.org/ (accessed on 30 September 2011).
- Karlsen, T.A.; Brinchmann, J.E. Liposome delivery of microRNA-145 to mesenchymal stem cells leads to immunological off-target effects mediated by RIG-I. Mol. Ther. 2013. [Google Scholar] [CrossRef]
- Olejniczak, M.; Galka, P.; Krzyzosiak, W.J. Sequence-non-specific effects of RNA interference triggers and microRNA regulators. Nucleic Acids Res 2010, 38, 1–16. [Google Scholar]
- Ender, C.; Krek, A.; Friedlander, M.R.; Beitzinger, M.; Weinmann, L.; Chen, W.; Pfeffer, S.; Rajewsky, N.; Meister, G. A human snoRNA with microRNA-like functions. Mol. Cell 2008, 32, 519–528. [Google Scholar]
- Rother, S.; Meister, G. Small RNAs derived from longer non-coding RNAs. Biochimie 2011, 93, 1905–1915. [Google Scholar]
- Brameier, M.; Herwig, A.; Reinhardt, R.; Walter, L.; Gruber, J. Human box C/D snoRNAs with miRNA like functions: Expanding the range of regulatory RNAs. Nucleic Acids Res 2011, 39, 675–686. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed sequence | Nucleotide position of RMRP | Number of reads | |
|---|---|---|---|
| 5′ end | 3′ end | ||
| CTGAAGGCCTGTATCCTAGGCT | 8 | 29 | 1 |
| CCTAGGCTACACACTGAGGACT | 22 | 43 | 1 |
| CCTAGGCTACACACTGAGGAC | 22 | 42 | 1 |
| ACTGAGGACTCTGTTCCTCCCC | 34 | 55 | 1 |
| ACTGAGGACTCTGTTCCTCCC | 34 | 54 | 1 |
| TGAGGACTCTGTTCCTCCCCTT | 36 | 57 | 1 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maida, Y.; Kyo, S.; Lassmann, T.; Hayashizaki, Y.; Masutomi, K. Off-Target Effect of Endogenous siRNA Derived from RMRP in Human Cells. Int. J. Mol. Sci. 2013, 14, 9305-9318. https://doi.org/10.3390/ijms14059305
Maida Y, Kyo S, Lassmann T, Hayashizaki Y, Masutomi K. Off-Target Effect of Endogenous siRNA Derived from RMRP in Human Cells. International Journal of Molecular Sciences. 2013; 14(5):9305-9318. https://doi.org/10.3390/ijms14059305
Chicago/Turabian StyleMaida, Yoshiko, Satoru Kyo, Timo Lassmann, Yoshihide Hayashizaki, and Kenkichi Masutomi. 2013. "Off-Target Effect of Endogenous siRNA Derived from RMRP in Human Cells" International Journal of Molecular Sciences 14, no. 5: 9305-9318. https://doi.org/10.3390/ijms14059305
APA StyleMaida, Y., Kyo, S., Lassmann, T., Hayashizaki, Y., & Masutomi, K. (2013). Off-Target Effect of Endogenous siRNA Derived from RMRP in Human Cells. International Journal of Molecular Sciences, 14(5), 9305-9318. https://doi.org/10.3390/ijms14059305