D-pinitol Inhibits Prostate Cancer Metastasis through Inhibition of αVβ3 Integrin by Modulating FAK, c-Src and NF-κB Pathways

 and
and 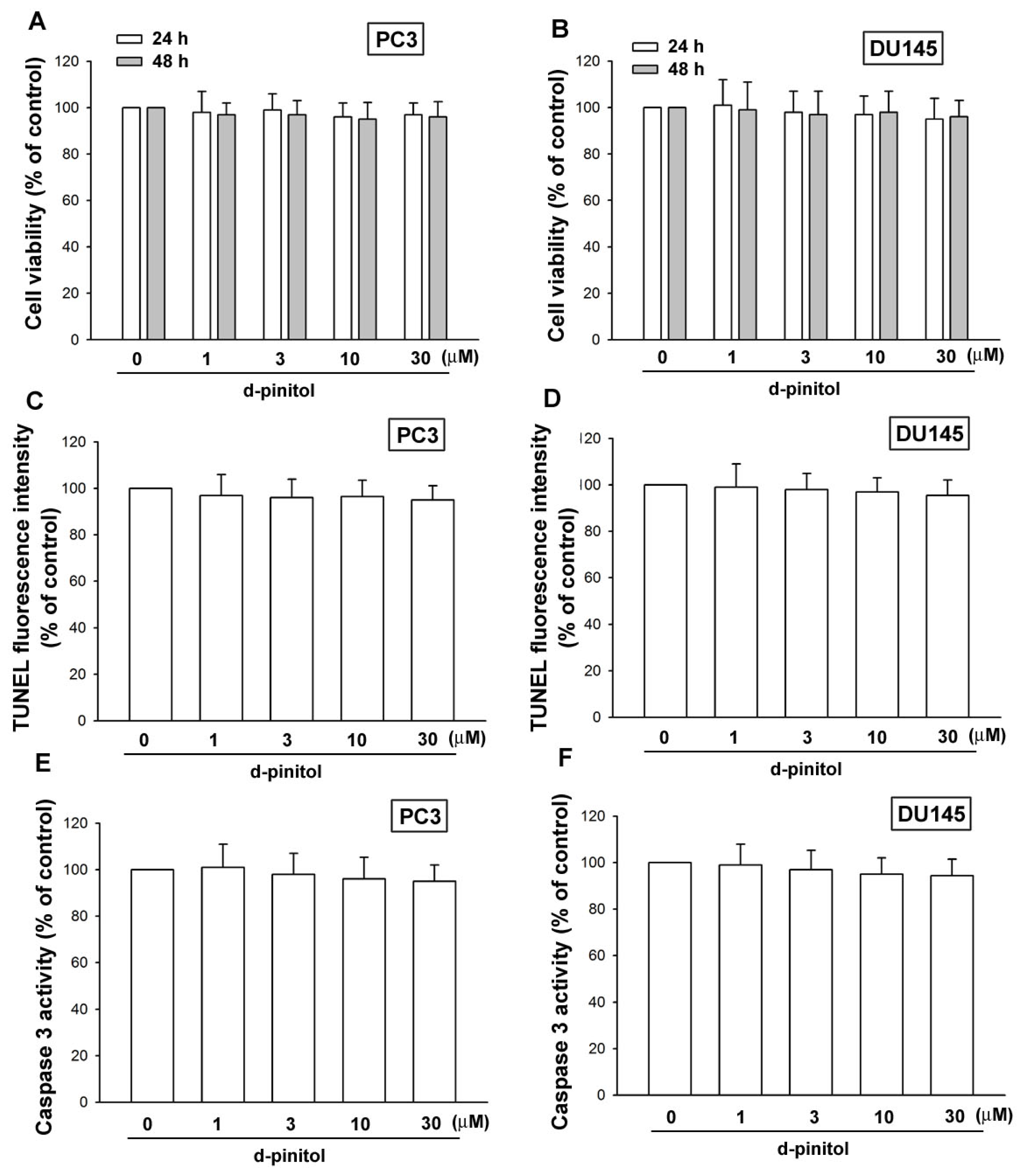{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. D-pinitol Does Not Induce Cell Death in Human Prostate Cancer Cells
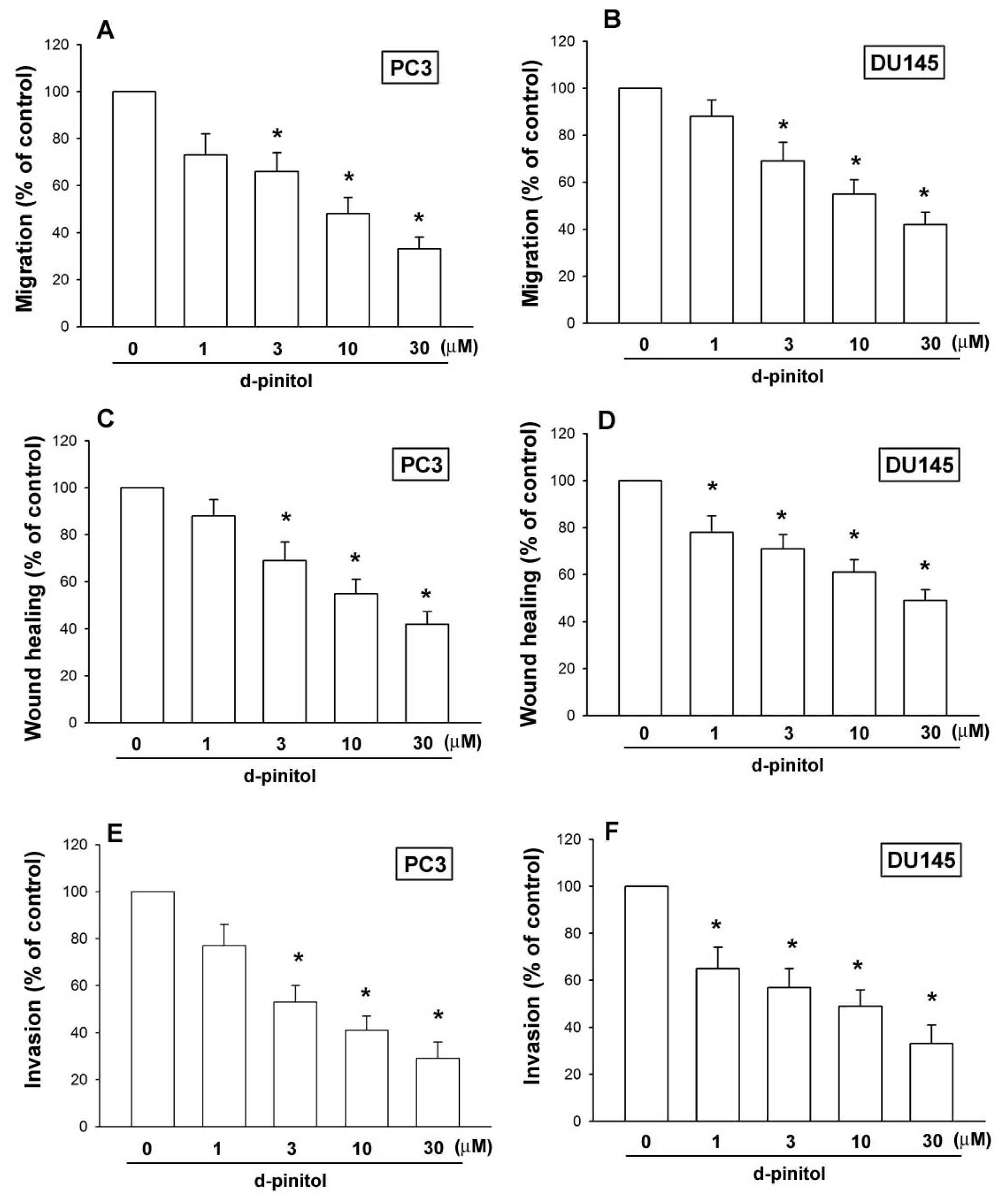
2.2. D-pinitol Reduces Cell Migration, Wound-Healing Migration and Invasion of Human Prostate Cancer Cells
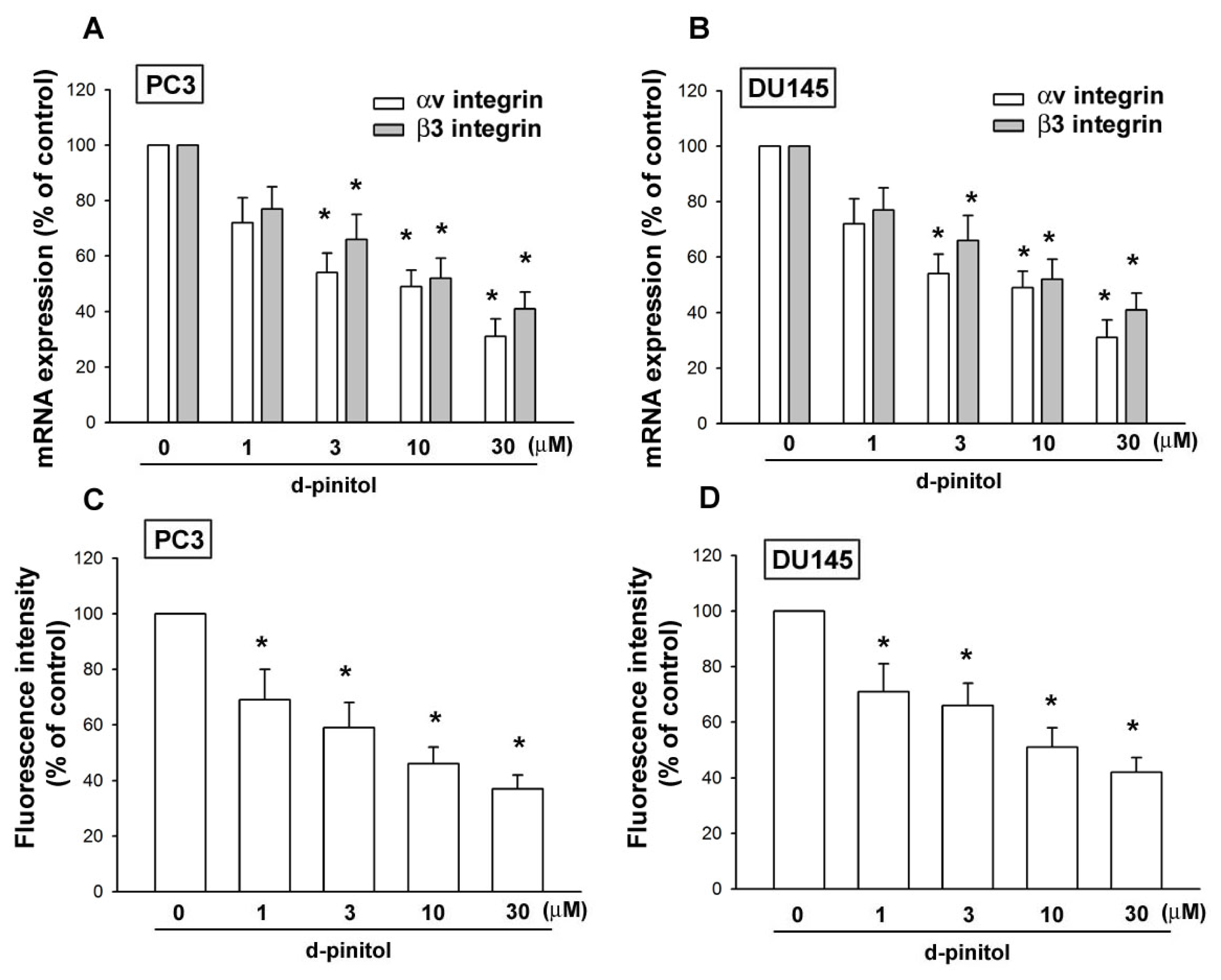
2.3. Involvement of αvβ3 Integrin Downregulation in D-pinitol-Reduced Cell Migration of Prostate Cancer Cells
2.4. The Effect of D-pinitol on FAK and c-Src Signaling Pathways
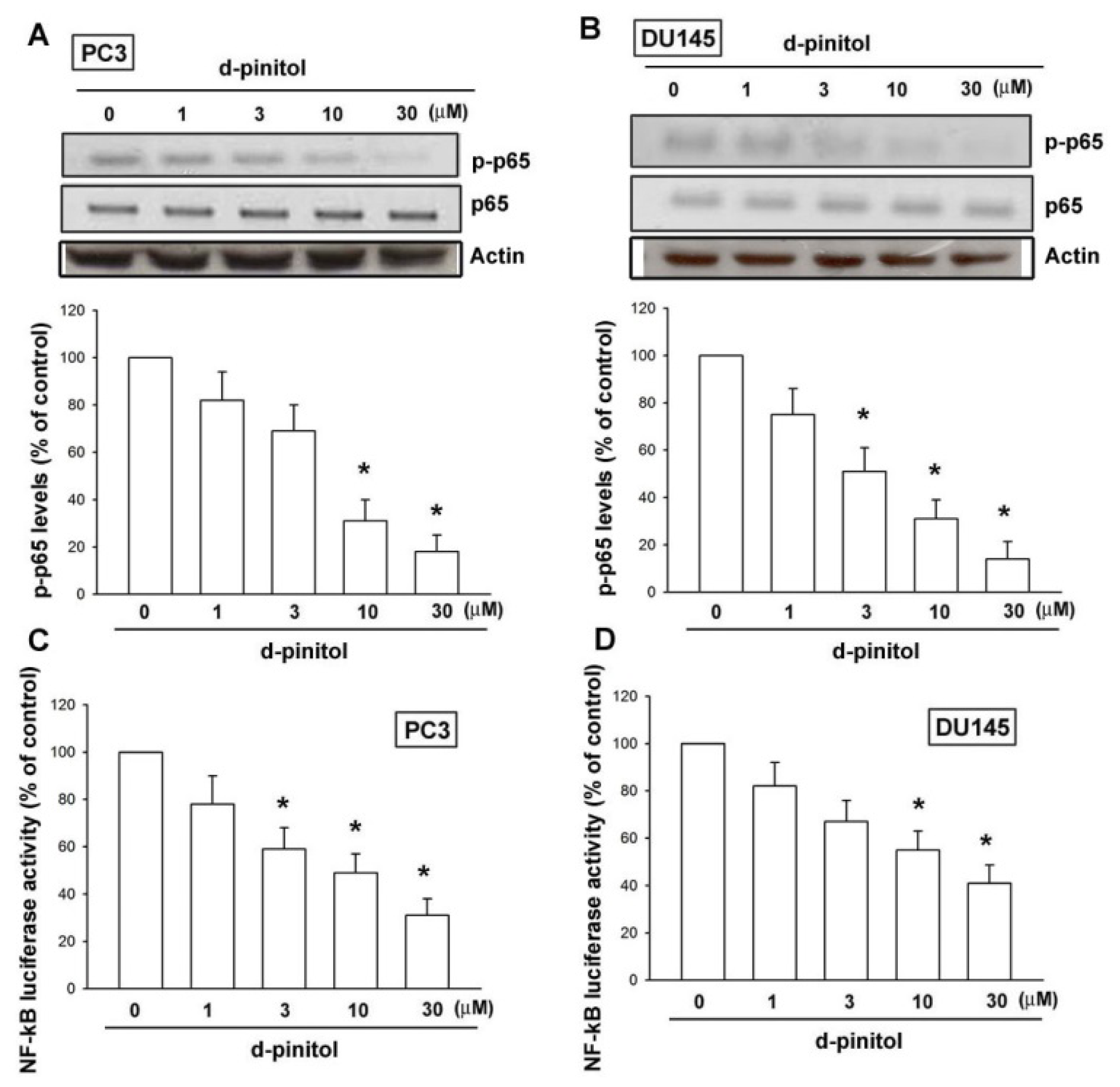
2.5. NF-κB Is Involved in D-pinitol-Reduced Cell Migration in Prostate Cancers
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Cell Culture
4.3. MTT Assay
4.4. TUNEL Assay
4.5. Caspase 3 Activity Assay
4.6. Migration and Invasion Assay
4.7. Wound-Healing Migration Assay
4.8. Flow Cytometric Analysis
4.9. Western Blot Analysis
4.10. Kinase Activity Assay
4.11. Quantitative Real-Time PCR
4.12. Reporter Gene Assay
4.13. Statistical Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Bryant, R.J.; Hamdy, F.C. Screening for prostate cancer: An update. Eur. Urol 2008, 53, 37–44. [Google Scholar]
- Ernst, D.S.; Hanson, J.; Venner, P.M. Analysis of prognostic factors in men with metastatic prostate cancer. Uro-Oncology Group of Northern Alberta. J. Urol 1991, 146, 372–376. [Google Scholar]
- Makrilia, N.; Kollias, A.; Manolopoulos, L.; Syrigos, K. Cell adhesion molecules: Role and clinical significance in cancer. Cancer Invest 2009, 27, 1023–1037. [Google Scholar]
- Giancotti, F.G.; Tarone, G. Positional control of cell fate through joint integrin/receptor protein kinase signaling. Annu. Rev. Cell Dev. Biol 2003, 19, 173–206. [Google Scholar]
- Wang, Y.; Shenouda, S.; Baranwal, S.; Rathinam, R.; Jain, P.; Bao, L.; Hazari, S.; Dash, S.; Alahari, S.K. Integrin subunits alpha5 and alpha6 regulate cell cycle by modulating the chk1 and Rb/E2F pathways to affect breast cancer metastasis. Mol. Cancer 2011, 10, 84. [Google Scholar]
- Shattil, S.J.; Kim, C.; Ginsberg, M.H. The final steps of integrin activation: The end game. Nat. Rev. Mol. Cell Biol 2010, 11, 288–300. [Google Scholar]
- Wang, Z.; Bryan, J.; Franz, C.; Havlioglu, N.; Sandell, L.J. Type IIB procollagen NH(2)-propeptide induces death of tumor cells via interaction with integrins alpha(V)beta(3) and alpha(V)beta(5). J. Biol. Chem 2010, 285, 20806–20817. [Google Scholar]
- Huang, C.Y.; Fong, Y.C.; Lee, C.Y.; Chen, M.Y.; Tsai, H.C.; Hsu, H.C.; Tang, C.H. CCL5 increases lung cancer migration via PI3K, Akt and NF-kappaB pathways. Biochem. Pharmacol 2009, 77, 794–803. [Google Scholar]
- Lu, D.Y.; Chang, C.S.; Yeh, W.L.; Tang, C.H.; Cheung, C.W.; Leung, Y.M.; Liu, J.F.; Wong, K.L. The novel phloroglucinol derivative BFP induces apoptosis of glioma cancer through reactive oxygen species and endoplasmic reticulum stress pathways. Phytomedicine 2012, 19, 1093–1100. [Google Scholar]
- Huang, C.Y.; Yu, H.S.; Lai, T.Y.; Yeh, Y.L.; Su, C.C.; Hsu, H.H.; Tsai, F.J.; Tsai, C.H.; Wu, H.C.; Tang, C.H. Leptin increases motility and integrin up-regulation in human prostate cancer cells. J. Cell Physiol 2011, 226, 1274–1282. [Google Scholar]
- Cooper, C.R.; Chay, C.H.; Pienta, K.J. The role of alpha(v)beta(3) in prostate cancer progression. Neoplasia 2002, 4, 191–194. [Google Scholar]
- Kumar, C.C. Integrin alpha v beta 3 as a therapeutic target for blocking tumor-induced angiogenesis. Curr. Drug Targets 2003, 4, 123–131. [Google Scholar]
- Streeter, J.G. Carbohydrates in Soybean Nodules: II. Distribution of compounds in seedlings during the onset of nitrogen fixation. Plant Physiol 1980, 66, 471–476. [Google Scholar]
- Numata, A.; Takahashi, C.; Fujiki, R.; Kitano, E.; Kitajima, A.; Takemura, T. Plant constituents biologically active to insects. VI. Antifeedants for larvae of the yellow butterfly, Eurema hecabe mandarina, in Osmunda japonica. (2). Chem. Pharm. Bull 1990, 38, 2862–2865. [Google Scholar]
- Singh, R.K.; Pandey, B.L.; Tripathi, M.; Pandey, V.B. Anti-inflammatory effect of (+)-pinitol. Fitoterapia 2001, 72, 168–170. [Google Scholar]
- Geethan, P.K.; Prince, P.S. Antihyperlipidemic effect of D-pinitol on streptozotocin-induced diabetic Wistar rats. J. Biochem. Mol. Toxicol 2008, 22, 220–224. [Google Scholar]
- Zhan, T.; Lou, H. Synthesis of azole nucleoside analogues of D-pinitol as potential antitumor agents. Carbohydr. Res 2007, 342, 865–869. [Google Scholar]
- Rengarajan, T.; Nandakumar, N.; Balasubramanian, M.P. D-pinitol attenuates 7, 12 dimethylbenz [a] anthracene induced hazards through modulating protein bound carbohydrates, adenosine triphosphatases and lysosomal enzymes during experimental mammary carcinogenesis. J. Exp. Ther. Oncol 2012, 10, 39–49. [Google Scholar]
- Sethi, G.; Ahn, K.S.; Sung, B.; Aggarwal, B.B. Pinitol targets nuclear factor-kappaB activation pathway leading to inhibition of gene products associated with proliferation, apoptosis, invasion, and angiogenesis. Mol. Cancer Ther 2008, 7, 1604–1614. [Google Scholar]
- Hsieh, M.T.; Hsieh, C.L.; Lin, L.W.; Wu, C.R.; Huang, G.S. Differential gene expression of scopolamine-treated rat hippocampus-application of cDNA microarray technology. Life Sci 2003, 73, 1007–1016. [Google Scholar]
- Hwangbo, C.; Kim, J.; Lee, J.J.; Lee, J.H. Activation of the integrin effector kinase focal adhesion kinase in cancer cells is regulated by crosstalk between protein kinase Calpha and the PDZ adapter protein mda-9/Syntenin. Cancer Res 2010, 70, 1645–1655. [Google Scholar]
- Boukerche, H.; Su, Z.Z.; Prevot, C.; Sarkar, D.; Fisher, P.B. mda-9/Syntenin promotes metastasis in human melanoma cells by activating c-Src. Proc. Natl. Acad. Sci. USA 2008, 105, 15914–15919. [Google Scholar]
- Park, J.; Singha, K.; Son, S.; Kim, J.; Namgung, R.; Yun, C.O.; Kim, W.J. A review of RGD-functionalized nonviral gene delivery vectors for cancer therapy. Cancer Gene Ther 2012, 19, 741–748. [Google Scholar]
- Deb, M.; Sengupta, D.; Patra, S.K. Integrin-epigenetics: Asystem with imperative impact on cancer. Cancer Metastasis Rev 2012, 31, 221–234. [Google Scholar]
- Liang, C.H.; Chiu, S.Y.; Hsu, I.L.; Wu, Y.Y.; Tsai, Y.T.; Ke, J.Y.; Pan, S.H.; Hsu, Y.C.; Li, K.C.; Yang, P.C.; et al. alpha-Catulin drives metastasis by activating ILK and driving an alphavbeta3 integrin signaling axis. Cancer Res 2013, 73, 428–438. [Google Scholar]
- Mundy, G.R. Metastasis to bone: causes, consequences and therapeutic opportunities. Nat. Rev. Cancer 2002, 2, 584–593. [Google Scholar]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev 2004, 25, 276–308. [Google Scholar]
- Lechertier, T.; Hodivala-Dilke, K. Focal adhesion kinase and tumour angiogenesis. J. Pathol 2012, 226, 404–412. [Google Scholar]
- Sliva, D. Signaling pathways responsible for cancer cell invasion as targets for cancer therapy. Curr. Cancer Drug Targets 2004, 4, 327–336. [Google Scholar]
- Fang, S.H.; Hou, Y.C.; Chang, W.C.; Hsiu, S.L.; Chao, P.D.; Chiang, B.L. Morin sulfates/ glucuronides exert anti-inflammatory activity on activated macrophages and decreased the incidence of septic shock. Life Sci 2003, 74, 743–756. [Google Scholar]
- Wang, Y.C.; Lee, P.J.; Shih, C.M.; Chen, H.Y.; Lee, C.C.; Chang, Y.Y.; Hsu, Y.T.; Liang, Y.J.; Wang, L.Y.; Han, W.H. Damage formation and repair efficiency in the p53 gene of cell lines and blood lymphocytes assayed by multiplex long quantitative polymerase chain reaction. Anal. Biochem 2003, 319, 206–215. [Google Scholar]
- Huang, H.C.; Shi, G.Y.; Jiang, S.J.; Shi, C.S.; Wu, C.M.; Yang, H.Y.; Wu, H.L. Thrombomodulin-mediated cell adhesion: involvement of its lectin-like domain. J. Biol. Chem 2003, 278, 46750–46759. [Google Scholar]
- Tseng, C.P.; Huang, C.L.; Huang, C.H.; Cheng, J.C.; Stern, A.; Tseng, C.H.; Chiu, D.T. Disabled-2 small interfering RNA modulates cellular adhesive function and MAPK activity during megakaryocytic differentiation of K562 cells. FEBS Lett 2003, 541, 21–27. [Google Scholar]
- Pezzuto, J.M. Plant-derived anticancer agents. Biochem. Pharmacol 1997, 53, 121–133. [Google Scholar]





© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lin, T.-H.; Tan, T.-W.; Tsai, T.-H.; Chen, C.-C.; Hsieh, T.-F.; Lee, S.-S.; Liu, H.-H.; Chen, W.-C.; Tang, C.-H. D-pinitol Inhibits Prostate Cancer Metastasis through Inhibition of αVβ3 Integrin by Modulating FAK, c-Src and NF-κB Pathways. Int. J. Mol. Sci. 2013, 14, 9790-9802. https://doi.org/10.3390/ijms14059790
Lin T-H, Tan T-W, Tsai T-H, Chen C-C, Hsieh T-F, Lee S-S, Liu H-H, Chen W-C, Tang C-H. D-pinitol Inhibits Prostate Cancer Metastasis through Inhibition of αVβ3 Integrin by Modulating FAK, c-Src and NF-κB Pathways. International Journal of Molecular Sciences. 2013; 14(5):9790-9802. https://doi.org/10.3390/ijms14059790
Chicago/Turabian StyleLin, Tien-Huang, Tzu-Wei Tan, Tsung-Hsun Tsai, Chi-Cheng Chen, Teng-Fu Hsieh, Shang-Sen Lee, Hsin-Ho Liu, Wen-Chi Chen, and Chih-Hsin Tang. 2013. "D-pinitol Inhibits Prostate Cancer Metastasis through Inhibition of αVβ3 Integrin by Modulating FAK, c-Src and NF-κB Pathways" International Journal of Molecular Sciences 14, no. 5: 9790-9802. https://doi.org/10.3390/ijms14059790
APA StyleLin, T.-H., Tan, T.-W., Tsai, T.-H., Chen, C.-C., Hsieh, T.-F., Lee, S.-S., Liu, H.-H., Chen, W.-C., & Tang, C.-H. (2013). D-pinitol Inhibits Prostate Cancer Metastasis through Inhibition of αVβ3 Integrin by Modulating FAK, c-Src and NF-κB Pathways. International Journal of Molecular Sciences, 14(5), 9790-9802. https://doi.org/10.3390/ijms14059790