Epigenetic Mechanisms in Penile Carcinoma

Abstract
:1. Introduction
2. Risk Factors: Human Papillomavirus as One of the Main Actors
3. Histopathological Data: Involvement of Lymph Nodes as the Major Predictive Factor of Poor Prognosis in Penile Cancer
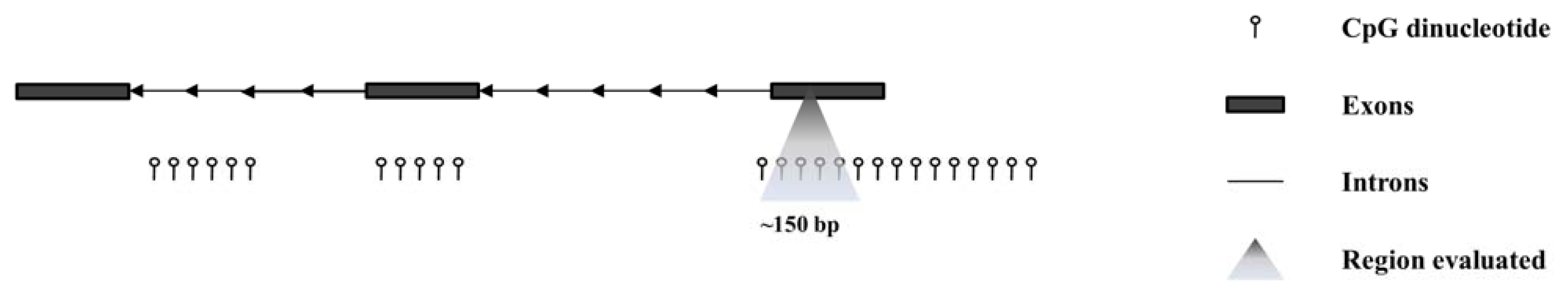
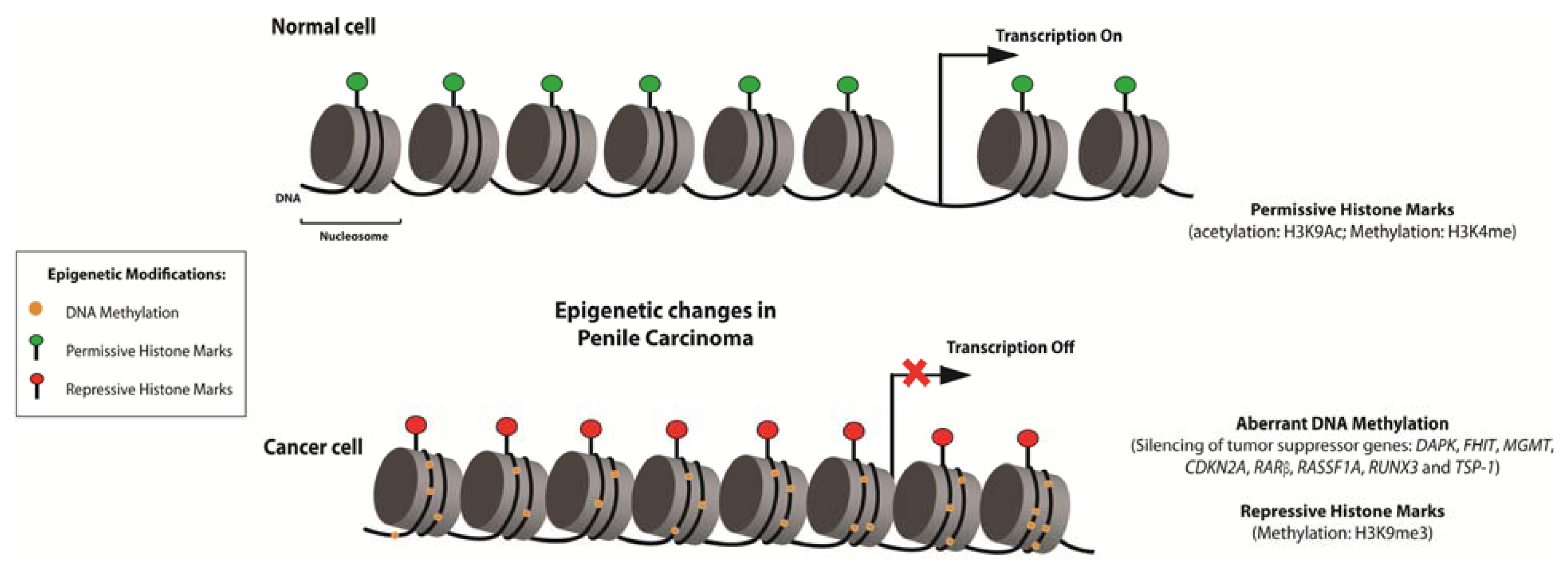
4. Epigenetic Alterations and Cancer: Emerging Potential Markers of Diagnosis, Prognosis, and Therapy
5. Epigenetics Studies in PeCa
6. Future Perspectives and Direction
Acknowledgments
Conflict of Interest
References
- Parkin, D.; Whelan, S.; Ferlay, J.; Teppo, L.; Thomas, D. Cancer Incidence in Five Continents; IARC Scientific Publications: Lyon, France, 2002; Volume VIII. [Google Scholar]
- Barnholtz-Sloan, J.S.; Maldonado, J.L.; Pow-sang, J.; Giuliano, A.R.; Guiliano, A.R. Incidence trends in primary malignant penile cancer. Urol. Oncol 2007, 25, 361–367. [Google Scholar]
- Misra, S.; Chaturvedi, A.; Misra, N.C. Penile carcinoma: A challenge for the developing world. Lancet Oncol 2004, 5, 240–247. [Google Scholar]
- Solsona, E.; Algaba, F.; Horenblas, S.; Pizzocaro, G.; Windahl, T.; Urology, E.A.O. Eau guidelines on penile cancer. Eur. Urol 2004, 46, 1–8. [Google Scholar]
- Favorito, L.A.; Nardi, A.C.; Ronalsa, M.; Zequi, S.C.; Sampaio, F.J.; Glina, S. Epidemiologic study on penile cancer in Brazil. Int. Braz. J. Urol 2008, 34, 587–591. [Google Scholar]
- Daling, J.R.; Madeleine, M.M.; Johnson, L.G.; Schwartz, S.M.; Shera, K.A.; Wurscher, M.A.; Carter, J.J.; Porter, P.L.; Galloway, D.A.; McDougall, J.K.; et al. Penile cancer: Importance of circumcision, human papillomavirus and smoking in in situ and invasive disease. Int. J. Cancer 2005, 116, 606–616. [Google Scholar]
- Ornellas, A.A.; Kinchin, E.W.; Nóbrega, B.L.; Wisnescky, A.; Koifman, N.; Quirino, R. Surgical treatment of invasive squamous cell carcinoma of the penis: Brazilian national cancer institute long-term experience. J. Surg. Oncol 2008, 97, 487–495. [Google Scholar]
- Calmon, M.F.; Tasso Mota, M.; Vassallo, J.; Rahal, P. Penile carcinoma: Risk factors and molecular alterations. ScientificWorldJournal 2011, 11, 269–282. [Google Scholar]
- Ficarra, V.; Akduman, B.; Bouchot, O.; Palou, J.; Tobias-Machado, M. Prognostic factors in penile cancer. Urology 2010, 76, S66–S73. [Google Scholar]
- Lopes, A.; Hidalgo, G.S.; Kowalski, L.P.; Torloni, H.; Rossi, B.M.; Fonseca, F.P. Prognostic factors in carcinoma of the penis: Multivariate analysis of 145 patients treated with amputation and lymphadenectomy. J. Urol 1996, 156, 1637–1642. [Google Scholar]
- Copeland, R.A.; Olhava, E.J.; Scott, M.P. Targeting epigenetic enzymes for drug discovery. Curr. Opin. Chem. Biol 2010, 14, 505–510. [Google Scholar]
- Nebbioso, A.; Carafa, V.; Benedetti, R.; Altucci, L. Trials with “epigenetic” drugs: An update. Mol. Oncol 2012, 6, 657–682. [Google Scholar]
- Dillner, J.; von Krogh, G.; Horenblas, S.; Meijer, C.J. Etiology of squamous cell carcinoma of the penis. Scand. J. Urol. Nephrol. Suppl 2000, 205, 189–193. [Google Scholar]
- Kochen, M.; McCurdy, S. Circumcision and the risk of cancer of the penis. A life-table analysis. Am. J. Dis. Child 1980, 134, 484–486. [Google Scholar]
- Harish, K.; Ravi, R. The role of tobacco in penile carcinoma. Br. J. Urol 1995, 75, 375–377. [Google Scholar]
- Miralles-Guri, C.; Bruni, L.; Cubilla, A.L.; Castellsagué, X.; Bosch, F.X.; de Sanjosé, S. Human papillomavirus prevalence and type distribution in penile carcinoma. J. Clin. Pathol 2009, 62, 870–878. [Google Scholar]
- Tsen, H.F.; Morgenstern, H.; Mack, T.; Peters, R.K. Risk factors for penile cancer: Results of a population-based case-control study in los angeles county (united states). Cancer Causes Control 2001, 12, 267–277. [Google Scholar]
- Madsen, B.S.; van den Brule, A.J.; Jensen, H.L.; Wohlfahrt, J.; Frisch, M. Risk factors for squamous cell carcinoma of the penis—Population-based case-control study in Denmark. Cancer Epidemiol. Biomark. Prev 2008, 17, 2683–2691. [Google Scholar]
- Schoen, E.J.; Oehrli, M.; Colby, C.; Machin, G. The highly protective effect of newborn circumcision against invasive penile cancer. Pediatrics 2000, 105, E36. [Google Scholar]
- Gross, G.; Pfister, H. Role of human papillomavirus in penile cancer, penile intraepithelial squamous cell neoplasias and in genital warts. Med. Microbiol. Immunol 2004, 193, 35–44. [Google Scholar]
- Zur Hausen, H. Papillomaviruses and cancer: From basic studies to clinical application. Nat. Rev. Cancer 2002, 2, 342–350. [Google Scholar]
- Rubin, M.A.; Kleter, B.; Zhou, M.; Ayala, G.; Cubilla, A.L.; Quint, W.G.; Pirog, E.C. Detection and typing of human papillomavirus dna in penile carcinoma: Evidence for multiple independent pathways of penile carcinogenesis. Am. J. Pathol 2001, 159, 1211–1218. [Google Scholar]
- Chaux, A.; Cubilla, A.L. The role of human papillomavirus infection in the pathogenesis of penile squamous cell carcinomas. Semin. Diagn. Pathol 2012, 29, 67–71. [Google Scholar]
- Pascual, A.; Pariente, M.; Godínez, J.M.; Sánchez-Prieto, R.; Atienzar, M.; Segura, M.; Poblet, E. High prevalence of human papillomavirus 16 in penile carcinoma. Histol. Histopathol 2007, 22, 177–183. [Google Scholar]
- Backes, D.M.; Kurman, R.J.; Pimenta, J.M.; Smith, J.S. Systematic review of human papillomavirus prevalence in invasive penile cancer. Cancer Causes Control 2009, 20, 449–457. [Google Scholar]
- Schottenfeld, D.; Beebe-Dimmer, J. Alleviating the burden of cancer: A perspective on advances, challenges, and future directions. Cancer Epidemiol. Biomarkers Prev 2006, 15, 2049–2055. [Google Scholar]
- Rombaldi, R.L.; Serafini, E.P.; Villa, L.L.; Vanni, A.C.; Baréa, F.; Frassini, R.; Xavier, M.; Paesi, S. Infection with human papillomaviruses of sexual partners of women having cervical intraepithelial neoplasia. Braz. J. Med. Biol. Res 2006, 39, 177–187. [Google Scholar]
- Gillison, M.L.; Chaturvedi, A.K.; Lowy, D.R. Hpv prophylactic vaccines and the potential prevention of noncervical cancers in both men and women. Cancer 2008, 113, 3036–3046. [Google Scholar]
- Al-Daraji, W.I.; Smith, J.H. Infection and cervical neoplasia: Facts and fiction. Int. J. Clin. Exp. Pathol 2009, 2, 48–64. [Google Scholar]
- Sasagawa, T.; Shimakage, M.; Nakamura, M.; Sakaike, J.; Ishikawa, H.; Inoue, M. Epstein-barr virus (ebv) genes expression in cervical intraepithelial neoplasia and invasive cervical cancer: A comparative study with human papillomavirus (hpv) infection. Hum. Pathol 2000, 31, 318–326. [Google Scholar]
- Afonso, L.A.; Moyses, N.; Alves, G.; Ornellas, A.A.; Passos, M.R.; Oliveira, L.O.H.; Cavalcanti, S.M. Prevalence of human papillomavirus and epstein-barr virus dna in penile cancer cases from brazil. Mem. Inst. Oswaldo. Cruz 2012, 107, 18–23. [Google Scholar]
- Buechner, S.A. Common skin disorders of the penis. BJU Int 2002, 90, 498–506. [Google Scholar]
- Schellhammer, P.F.; Jordan, G.H.; Robey, E.L.; Spaulding, J.T. Premalignant lesions and nonsquamous malignancy of the penis and carcinoma of the scrotum. Urol. Clin. N. Am 1992, 19, 131–142. [Google Scholar]
- Wieland, U.; Jurk, S.; Weissenborn, S.; Krieg, T.; Pfister, H.; Ritzkowsky, A. Erythroplasia of queyrat: Coinfection with cutaneous carcinogenic human papillomavirus type 8 and genital papillomaviruses in a carcinoma in situ. J. Invest. Dermatol 2000, 115, 396–401. [Google Scholar]
- Val, I.; Almeida, G. An overview of lichen sclerosus. Clin. Obstet. Gynecol 2005, 48, 808–817. [Google Scholar]
- Tasker, G.L.; Wojnarowska, F. Lichen sclerosus. Clin. Exp. Dermatol 2003, 28, 128–133. [Google Scholar]
- Sufrin, G.; Huber, R. Benign and Malignant Lesions of the Penis; J.Y. Gillenwater: St. Louis, MO, USA, 1991; pp. 1997–2042. [Google Scholar]
- Mosconi, A.M.; Roila, F.; Gatta, G.; Theodore, C. Cancer of the penis. Crit. Rev. Oncol. Hematol 2005, 53, 165–177. [Google Scholar]
- Chaux, A.; Velazquez, E.F.; Algaba, F.; Ayala, G.; Cubilla, A.L. Developments in the pathology of penile squamous cell carcinomas. Urology 2010, 76, S7–S14. [Google Scholar]
- Pizzocaro, G.; Algaba, F.; Horenblas, S.; Solsona, E.; Tana, S.; van der Poel, H.; Watkin, N.A. Eau penile cancer guidelines 2009. Eur. Urol 2010, 57, 1002–1012. [Google Scholar]
- Cubilla, A.L.; Velazquez, E.F.; Young, R.H. Epithelial lesions associated with invasive penile squamous cell carcinoma: A pathologic study of 288 cases. Int. J. Surg. Pathol 2004, 12, 351–364. [Google Scholar]
- Cubilla, A.L.; Barreto, J.; Caballero, C.; Ayala, G.; Riveros, M. Pathologic features of epidermoid carcinoma of the penis. A prospective study of 66 cases. Am. J. Surg. Pathol 1993, 17, 753–763. [Google Scholar]
- Lucia, M.S.; Miller, G.J. Histopathology of malignant lesions of the penis. Urol. Clin. N. Am 1992, 19, 227–246. [Google Scholar]
- Jackson, S.M. The treatment of carcinoma of the penis. Br. J. Surg 1966, 53, 33–35. [Google Scholar]
- Sobin, L.; Wittekind, C. TNM Classification of Malignant Tumours, 6th ed.; Wiley-Liss: New York, NY, USA, 2002. [Google Scholar]
- Kroon, B.K.; Horenblas, S.; Lont, A.P.; Tanis, P.J.; Gallee, M.P.; Nieweg, O.E. Patients with penile carcinoma benefit from immediate resection of clinically occult lymph node metastases. J. Urol 2005, 173, 816–819. [Google Scholar]
- Ravi, R. Morbidity following groin dissection for penile carcinoma. Br. J. Urol 1993, 72, 941–945. [Google Scholar]
- Britto, C.A.; Rebouças, R.B.; Lopes, T.R.; Silva da Costa, T.; Leite, R.E.C.; de Carvalho, P.S. Video-assisted left inguinal lymphadenectomy for penile cancer. Int. Braz. J. Urol 2012, 38, 289. [Google Scholar]
- Canter, D.J.; Dobbs, R.W.; Jafri, S.M.; Herrel, L.A.; Ogan, K.; Delman, K.A.; Master, V.A. Functional, oncologic, and technical outcomes after endoscopic groin dissection for penile carcinoma. Can. J. Urol 2012, 19, 6395–6400. [Google Scholar]
- Kattan, M.W.; Ficarra, V.; Artibani, W.; Cunico, S.C.; Fandella, A.; Martignoni, G.; Novara, G.; Galetti, T.P.; Zattoni, F.; Members, G.P.C.P. Nomogram predictive of cancer specific survival in patients undergoing partial or total amputation for squamous cell carcinoma of the penis. J. Urol 2006, 175, 2103–2108. [Google Scholar]
- Ficarra, V.; Zattoni, F.; Artibani, W.; Fandella, A.; Martignoni, G.; Novara, G.; Galetti, T.P.; Zambolin, T.; Kattan, M.W. Penile Cancer Project Members. Nomogram predictive of pathological inguinal lymph node involvement in patients with squamous cell carcinoma of the penis. J. Urol. 2006, 175, 1700–1704. [Google Scholar]
- Gregoire, L.; Cubilla, A.L.; Reuter, V.E.; Haas, G.P.; Lancaster, W.D. Preferential association of human papillomavirus with high-grade histologic variants of penile-invasive squamous cell carcinoma. J. Natl. Cancer Inst 1995, 87, 1705–1709. [Google Scholar]
- Burger, R.A.; Monk, B.J.; Kurosaki, T.; Anton-Culver, H.; Vasilev, S.A.; Berman, M.L.; Wilczynski, S.P. Human papillomavirus type 18: Association with poor prognosis in early stage cervical cancer. J. Natl. Cancer Inst 1996, 88, 1361–1368. [Google Scholar]
- Bezerra, A.L.; Lopes, A.; Santiago, G.H.; Ribeiro, K.C.; Latorre, M.R.; Villa, L.L. Human papillomavirus as a prognostic factor in carcinoma of the penis: Analysis of 82 patients treated with amputation and bilateral lymphadenectomy. Cancer 2001, 91, 2315–2321. [Google Scholar]
- Kirrander, P.; Kolaric, A.; Helenius, G.; Windahl, T.; Andrén, O.; Stark, J.R.; Lillsunde-Larsson, G.; Elgh, F.; Karlsson, M. Human papillomavirus prevalence, distribution and correlation to histopathological parameters in a large swedish cohort of men with penile carcinoma. BJU Int 2011, 108, 355–359. [Google Scholar]
- Zhai, J.P.; Li, M.; Wang, Q.Y.; Wei, D.; Xu, K.X. Correlation of cd82 and htert expressions and hpv infection with penile cancer. Zhonghua Nan Ke Xue 2011, 17, 817–822. [Google Scholar]
- Lont, A.P.; Kroon, B.K.; Horenblas, S.; Gallee, M.P.; Berkhof, J.; Meijer, C.J.; Snijders, P.J. Presence of high-risk human papillomavirus dna in penile carcinoma predicts favorable outcome in survival. Int. J. Cancer 2006, 119, 1078–1081. [Google Scholar]
- Herceg, Z.; Hernandez-Vargas, H. New concepts of old epigenetic phenomena and their implications for selecting specific cell populations for epigenomic research. Epigenomics 2011, 3, 383–386. [Google Scholar]
- Feinberg, A.P.; Ohlsson, R.; Henikoff, S. The epigenetic progenitor origin of human cancer. Nat. Rev. Genet 2006, 7, 21–33. [Google Scholar]
- Jones, P.A.; Baylin, S.B. The epigenomics of cancer. Cell 2007, 128, 683–692. [Google Scholar]
- Feinberg, A.P. Phenotypic plasticity and the epigenetics of human disease. Nature 2007, 447, 433–440. [Google Scholar]
- Willard, S.S.; Koochekpour, S. Regulators of gene expression as biomarkers for prostate cancer. Am. J. Cancer Res 2012, 2, 620–657. [Google Scholar]
- Jovanovic, J.; Rønneberg, J.A.; Tost, J.; Kristensen, V. The epigenetics of breast cancer. Mol. Oncol 2010, 4, 242–254. [Google Scholar]
- Sandoval, J.; Esteller, M. Cancer epigenomics: Beyond genomics. Curr. Opin. Genet. Dev 2012, 22, 50–55. [Google Scholar]
- Bestor, T.H. The DNA methyltransferases of mammals. Hum. Mol. Genet 2000, 9, 2395–2402. [Google Scholar]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev 2002, 16, 6–21. [Google Scholar]
- Takai, D.; Jones, P.A. Comprehensive analysis of CPG islands in human chromosomes 21 and 22. Proc. Natl. Acad. Sci. USA 2002, 99, 3740–3745. [Google Scholar]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet 2008, 9, 465–476. [Google Scholar]
- Wang, Y.; Leung, F.C. An evaluation of new criteria for CPG islands in the human genome as gene markers. Bioinformatics 2004, 20, 1170–1177. [Google Scholar]
- Rodriguez, J.; Frigola, J.; Vendrell, E.; Risques, R.A.; Fraga, M.F.; Morales, C.; Moreno, V.; Esteller, M.; Capellà, G.; Ribas, M.; et al. Chromosomal instability correlates with genome-wide dna demethylation in human primary colorectal cancers. Cancer Res 2006, 66, 8462–9468. [Google Scholar]
- Dahl, C.; Guldberg, P. DNA methylation analysis techniques. Biogerontology 2003, 4, 233–250. [Google Scholar]
- Herman, J.G.; Graff, J.R.; Myöhänen, S.; Nelkin, B.D.; Baylin, S.B. Methylation-specific PCR: A novel PCR assay for methylation status of CPG islands. Proc. Natl. Acad. Sci. USA 1996, 93, 9821–9826. [Google Scholar]
- Eads, C.A.; Danenberg, K.D.; Kawakami, K.; Saltz, L.B.; Blake, C.; Shibata, D.; Danenberg, P.V.; Laird, P.W. Methylight: A high-throughput assay to measure DNA methylation. Nucleic Acids Res 2000, 28, E32. [Google Scholar]
- Xiong, Z.; Laird, P.W. Cobra: A sensitive and quantitative DNA methylation assay. Nucleic Acids Res 1997, 25, 2532–2534. [Google Scholar]
- Gonzalgo, M.L.; Jones, P.A. Rapid quantitation of methylation differences at specific sites using methylation-sensitive single nucleotide primer extension (ms-snupe). Nucleic Acids Res 1997, 25, 2529–2531. [Google Scholar]
- Wojdacz, T.K.; Dobrovic, A. Methylation-sensitive high resolution melting (ms-hrm): A new approach for sensitive and high-throughput assessment of methylation. Nucleic Acids Res 2007, 35, e41. [Google Scholar]
- Tost, J.; Gut, I.G. DNA methylation analysis by pyrosequencing. Nat. Protoc 2007, 2, 2265–2275. [Google Scholar]
- Mohn, F.; Weber, M.; Schübeler, D.; Roloff, T.C. Methylated DNA immunoprecipitation (medip). Methods Mol. Biol 2009, 507, 55–64. [Google Scholar]
- Irizarry, R.A.; Ladd-Acosta, C.; Carvalho, B.; Wu, H.; Brandenburg, S.A.; Jeddeloh, J.A.; Wen, B.; Feinberg, A.P. Comprehensive high-throughput arrays for relative methylation (charm). Genome Res 2008, 18, 780–790. [Google Scholar]
- Serre, D.; Lee, B.H.; Ting, A.H. Mbd-isolated genome sequencing provides a high-throughput and comprehensive survey of DNA methylation in the human genome. Nucleic Acids Res 2010, 38, 391–399. [Google Scholar]
- Karimi, M.; Johansson, S.; Ekström, T.J. Using luma: A luminometric-based assay for global DNA-methylation. Epigenetics 2006, 1, 45–48. [Google Scholar]
- Oda, M.; Greally, J.M. The help assay. Methods Mol. Biol 2009, 507, 77–87. [Google Scholar]
- Sonnet, M.; Baer, C.; Rehli, M.; Weichenhan, D.; Plass, C. Enrichment of methylated DNA by methyl-CPG immunoprecipitation. Methods Mol. Biol 2013, 971, 201–212. [Google Scholar]
- Ammerpohl, O.; Martín-Subero, J.I.; Richter, J.; Vater, I.; Siebert, R. Hunting for the 5th base: Techniques for analyzing DNA methylation. Biochim. Biophys. Acta 2009, 1790, 847–862. [Google Scholar]
- Brebi-Mieville, P.; Ili-Gangas, C.; Leal-Rojas, P.; Noordhuis, M.G.; Soudry, E.; Perez, J.; Roa, J.C.; Sidransky, D.; Guerrero-Preston, R. Clinical and public health research using methylated dna immunoprecipitation (medip): A comparison of commercially available kits to examine differential DNA methylation across the genome. Epigenetics 2012, 7, 106–112. [Google Scholar]
- Wei, S.H.; Chen, C.M.; Strathdee, G.; Harnsomburana, J.; Shyu, C.R.; Rahmatpanah, F.; Shi, H.; Ng, S.W.; Yan, P.S.; Nephew, K.P.; et al. Methylation microarray analysis of late-stage ovarian carcinomas distinguishes progression-free survival in patients and identifies candidate epigenetic markers. Clin. Cancer Res 2002, 8, 2246–2252. [Google Scholar]
- Vincent, A.; Omura, N.; Hong, S.M.; Jaffe, A.; Eshleman, J.; Goggins, M. Genome-wide analysis of promoter methylation associated with gene expression profile in pancreatic adenocarcinoma. Clin. Cancer Res 2011, 17, 4341–4354. [Google Scholar]
- De Capoa, A.; Musolino, A.; Della Rosa, S.; Caiafa, P.; Mariani, L.; del Nonno, F.; Vocaturo, A.; Donnorso, R.P.; Niveleau, A.; Grappelli, C. DNA demethylation is directly related to tumour progression: Evidence in normal, pre-malignant and malignant cells from uterine cervix samples. Oncol. Rep 2003, 10, 545–549. [Google Scholar]
- Hernandez-Vargas, H.; Lambert, M.P.; le Calvez-Kelm, F.; Gouysse, G.; McKay-Chopin, S.; Tavtigian, S.V.; Scoazec, J.Y.; Herceg, Z. Hepatocellular carcinoma displays distinct DNA methylation signatures with potential as clinical predictors. PLoS One 2010, 5, e9749. [Google Scholar]
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 a resolution. Nature 1997, 389, 251–260. [Google Scholar]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar]
- Huang, Y.; Nayak, S.; Jankowitz, R.; Davidson, N.E.; Oesterreich, S. Epigenetics in breast cancer: What’s new? Breast Cancer Res 2011, 13, 225. [Google Scholar]
- Linggi, B.E.; Brandt, S.J.; Sun, Z.W.; Hiebert, S.W. Translating the histone code into leukemia. J. Cell Biochem 2005, 96, 938–950. [Google Scholar]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microrna biogenesis, function and decay. Nat. Rev. Genet 2010, 11, 597–610. [Google Scholar]
- Lopez-Serra, P.; Esteller, M. DNA methylation-associated silencing of tumor-suppressor micrornas in cancer. Oncogene 2012, 31, 1609–1622. [Google Scholar]
- Calin, G.A.; Croce, C.M. Microrna-cancer connection: The beginning of a new tale. Cancer Res 2006, 66, 7390–7394. [Google Scholar]
- Thomson, J.M.; Newman, M.; Parker, J.S.; Morin-Kensicki, E.M.; Wright, T.; Hammond, S.M. Extensive post-transcriptional regulation of micrornas and its implications for cancer. Genes Dev 2006, 20, 2202–2207. [Google Scholar]
- Chang, T.C.; Yu, D.; Lee, Y.S.; Wentzel, E.A.; Arking, D.E.; West, K.M.; Dang, C.V.; Thomas-Tikhonenko, A.; Mendell, J.T. Widespread microrna repression by MYC contributes to tumorigenesis. Nat. Genet 2008, 40, 43–50. [Google Scholar]
- Melo, S.A.; Kalluri, R. Molecular pathways: Micrornas as cancer therapeutics. Clin. Cancer Res 2012, 18, 4234–4239. [Google Scholar]
- Yamamoto, H.; Adachi, Y.; Taniguchi, H.; Kunimoto, H.; Nosho, K.; Suzuki, H.; Shinomura, Y. Interrelationship between microsatellite instability and microrna in gastrointestinal cancer. World J. Gastroenterol 2012, 18, 2745–2755. [Google Scholar]
- Cho, W.C. Great potential of mirnas as predictive and prognostic markers for cancer. Expert Rev. Mol. Diagn 2012, 12, 315–318. [Google Scholar]
- Kelly, T.K.; de Carvalho, D.D.; Jones, P.A. Epigenetic modifications as therapeutic targets. Nat. Biotechnol 2010, 28, 1069–1078. [Google Scholar]
- Huang, Z.; Jiang, H.; Liu, X.; Chen, Y.; Wong, J.; Wang, Q.; Huang, W.; Shi, T.; Zhang, J. Hemd: An integrated tool of human epigenetic enzymes and chemical modulators for therapeutics. PLoS One 2012, 7, e39917. [Google Scholar]
- Ferreux, E.; Lont, A.P.; Horenblas, S.; Gallee, M.P.; Raaphorst, F.M.; von Knebel Doeberitz, M.; Meijer, C.J.; Snijders, P.J. Evidence for at least three alternative mechanisms targeting the p16ink4a/cyclin d/rb pathway in penile carcinoma, one of which is mediated by high-risk human papillomavirus. J. Pathol 2003, 201, 109–118. [Google Scholar]
- Soufir, N.; Queille, S.; Liboutet, M.; Thibaudeau, O.; Bachelier, F.; Delestaing, G.; Balloy, B.C.; Breuer, J.; Janin, A.; Dubertret, L.; et al. Inactivation of the cdkn2a and the p53 tumour suppressor genes in external genital carcinomas and their precursors. Br. J. Dermatol 2007, 156, 448–453. [Google Scholar]
- Kalantari, M.; Villa, L.L.; Calleja-Macias, I.E.; Bernard, H.U. Human papillomavirus-16 and -18 in penile carcinomas: DNA methylation, chromosomal recombination and genomic variation. Int. J. Cancer 2008, 123, 1832–1840. [Google Scholar]
- Guerrero, D.; Guarch, R.; Ojer, A.; Casas, J.M.; Ropero, S.; Mancha, A.; Pesce, C.; Lloveras, B.; Garcia-Bragado, F.; Puras, A. Hypermethylation of the thrombospondin-1 gene is associated with poor prognosis in penile squamous cell carcinoma. BJU Int 2008, 102, 747–755. [Google Scholar]
- Yanagawa, N.; Osakabe, M.; Hayashi, M.; Tamura, G.; Motoyama, T. Detection of HPV-DNA, p53 alterations, and methylation in penile squamous cell carcinoma in japanese men. Pathol. Int 2008, 58, 477–482. [Google Scholar]
- Yanagawa, N.; Osakabe, M.; Hayashi, M.; Tamura, G.; Motoyama, T. Frequent epigenetic silencing of the FHIT gene in penile squamous cell carcinomas. Virchows. Arch 2008, 452, 377–382. [Google Scholar]
- Rogenhofer, S.; Miersch, H.; Göke, F.; Kahl, P.; Wieland, W.F.; Hofstädter, F.; Kristiansen, G.; von Ruecker, A.; Müller, S.C.; Ellinger, J. Histone methylation defines an epigenetic entity in penile squamous cell carcinoma. J. Urol 2012, 189, 1117–1122. [Google Scholar]
- Poetsch, M.; Hemmerich, M.; Kakies, C.; Kleist, B.; Wolf, E.; vom Dorp, F.; Hakenberg, O.W.; Protzel, C. Alterations in the tumor suppressor gene p16(ink4a) are associated with aggressive behavior of penile carcinomas. Virchows. Arch 2011, 458, 221–229. [Google Scholar]
- Larsen, C.J. Contribution of the dual coding capacity of the p16ink4a/mts1/cdkn2 locus to human malignancies. Prog. Cell Cycle Res 1997, 3, 109–124. [Google Scholar]
- Kim, W.Y.; Sharpless, N.E. The regulation of ink4/arf in cancer and aging. Cell 2006, 127, 265–275. [Google Scholar]
- Jacobs, J.J.; Kieboom, K.; Marino, S.; DePinho, R.A.; van Lohuizen, M. The oncogene and polycomb-group gene bmi-1 regulates cell proliferation and senescence through the ink4a locus. Nature 1999, 397, 164–168. [Google Scholar]
- Deng, Z.; Hasegawa, M.; Yamashita, Y.; Matayoshi, S.; Kiyuna, A.; Agena, S.; Uehara, T.; Maeda, H.; Suzuki, M. Prognostic value of human papillomavirus and squamous cell carcinoma antigen in head and neck squamous cell carcinoma. Cancer Sci 2012, 103, 2127–2134. [Google Scholar]
- Huang, H.; Zhang, B.; Chen, W.; Zhou, S.M.; Zhang, Y.X.; Gao, L.; Xu, Z.G.; Qiao, Y.L.; Tang, P.Z. Human papillomavirus infection and prognostic predictors in patients with oropharyngeal squamous cell carcinoma. Asian Pac. J. Cancer Prev 2012, 13, 891–896. [Google Scholar]
- Bornstein, P. Thrombospondins as matricellular modulators of cell function. J. Clin. Invest 2001, 107, 929–934. [Google Scholar]
- Sonpavde, G.; Pagliaro, L.C.; Buonerba, C.; Dorff, T.B.; Lee, R.J.; di Lorenzo, G. Penile cancer: Current therapy and future directions. Ann. Oncol 2013, 24, 1179–1189. [Google Scholar]



{kind=link}
{kind=link}
| References | Number of samples | Method | Gene studied | % methylation | HPV infection | HPV 16 |
|---|---|---|---|---|---|---|
| Ferreux et al. [105] | 53 | Methylation-specific PCR | CDKN2A | 9 (17%) | 20 (38%) | 15 (28%) |
| Poetsch et al. [112] | 52 | Methylation-specific PCR | CDKN2A | 22 (42%) | 20 (38%) | 18 (35%) |
| Soufir et al. [106] | 3 | Methylation-specific PCR | CDKN2A | 0 (0%) | 2 (66.3%) | 2 (100%) |
| Guerreto et al. [108] | 24 | Methylation-specific PCR | CDKN2A RASSF1A TSP-1 | 9 (38%) 10 (42%) 11 (46%) | 11 (46%) | 10 (42%) |
| Yanagawa et al. [109] | 26 | Methylation-specific PCR | DAPK FHIT MGMT CDKN2A (p16INK4A) CDKN2A (p14ARF) RARβ RASSF1A RUNX3 | 7 (26.9%) 23 (88.4%) 5 (19.2%) 1 (3.8%) 6 (23.1%) 6 (23.1%) 3 (11.5%) 11 (42.3%) | 3 (11.5%) | 3 (11.5%) |
| Yanagawa et al. [110] | 25 | Methylation-specific PCR | DAPK FHIT MGMT CDKN2A (p16INK4A) CDKN2A (p14ARF) RARβ RASSF1A RUNX3 | 7 (28%) 23 (92%) 5(20%) 1 (4%) 6 (24%) 6 (24%) 3 (12%) 11 (44%) | 3 (12%) | 3 (12%) |
| Kalantari et al. [107] | 24 | DNA sequencing | L1 HPV16 LCR HPV16 | 58% 22% | 24 (100%) | 19 (79%) |
| Rogenhofer et al. [111] | 65 | Immunohistochemical | H3K4 H3K9 H3K27 | H3K4me1 Decrease H3K9me1 Decrease H3K9me2 Decrease H3K27me2 Decrease H3K27me3 Decrease H3K9me3 Increase |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kuasne, H.; Marchi, F.A.; Rogatto, S.R.; De Syllos Cólus, I.M. Epigenetic Mechanisms in Penile Carcinoma. Int. J. Mol. Sci. 2013, 14, 10791-10808. https://doi.org/10.3390/ijms140610791
Kuasne H, Marchi FA, Rogatto SR, De Syllos Cólus IM. Epigenetic Mechanisms in Penile Carcinoma. International Journal of Molecular Sciences. 2013; 14(6):10791-10808. https://doi.org/10.3390/ijms140610791
Chicago/Turabian StyleKuasne, Hellen, Fabio Albuquerque Marchi, Silvia Regina Rogatto, and Ilce Mara De Syllos Cólus. 2013. "Epigenetic Mechanisms in Penile Carcinoma" International Journal of Molecular Sciences 14, no. 6: 10791-10808. https://doi.org/10.3390/ijms140610791