Early Exercise Protects the Blood-Brain Barrier from Ischemic Brain Injury via the Regulation of MMP-9 and Occludin in Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Physiological Variables
2.2. Early Exercise Improved Neurological Function
2.3. Early Exercise Reduced Infarct Volume
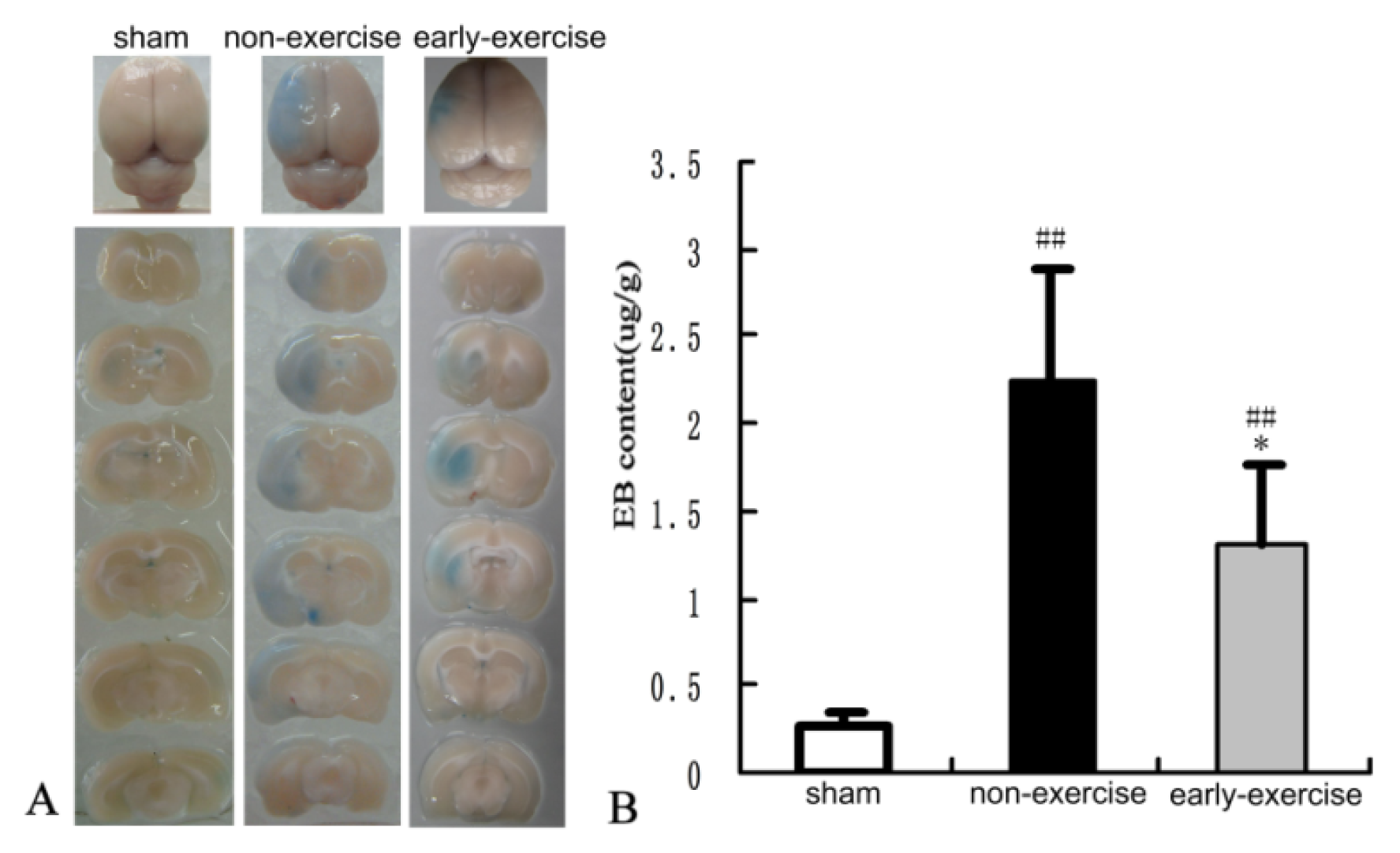
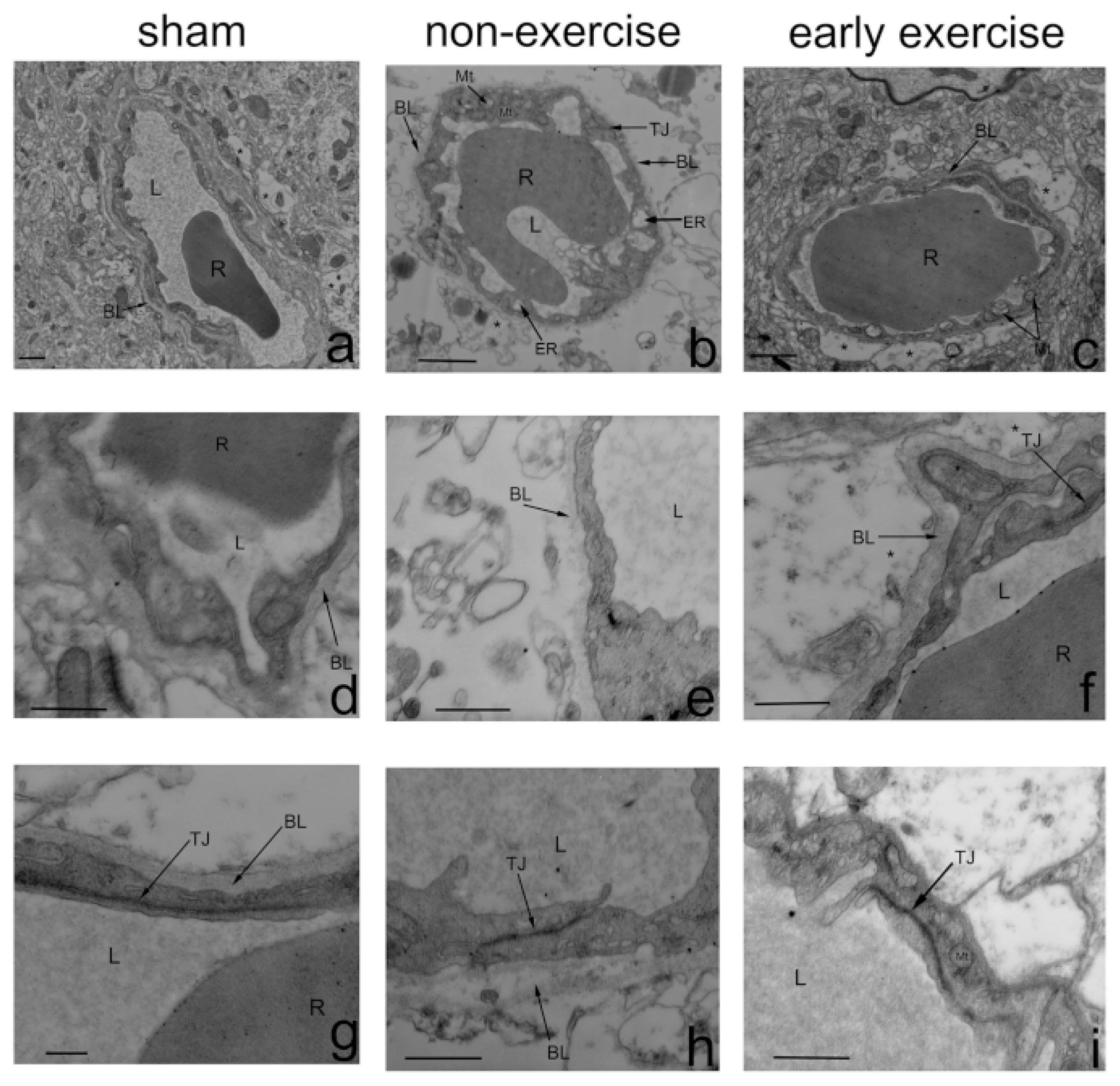
2.4. Early Exercise Preserved BBB Integrity
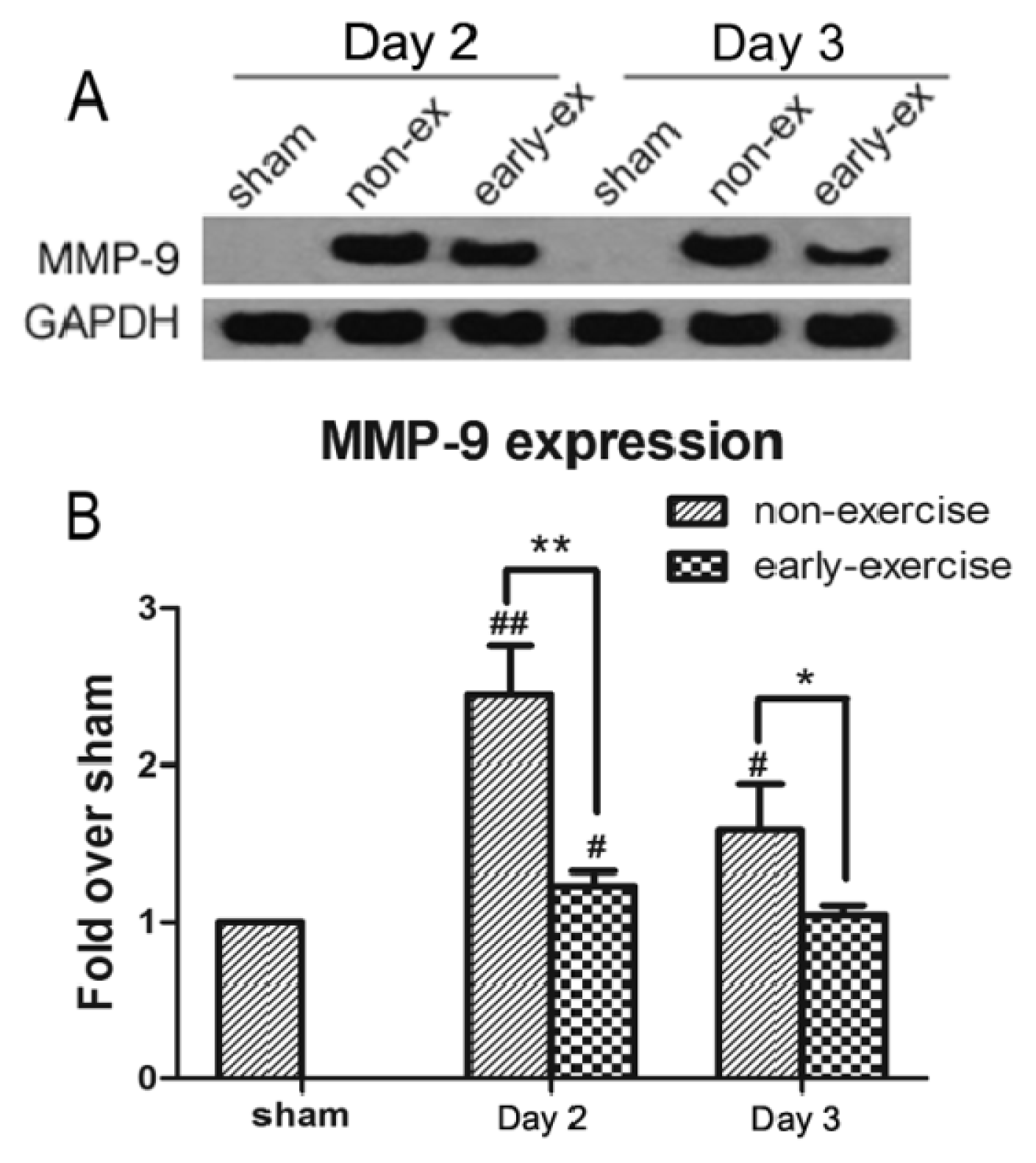
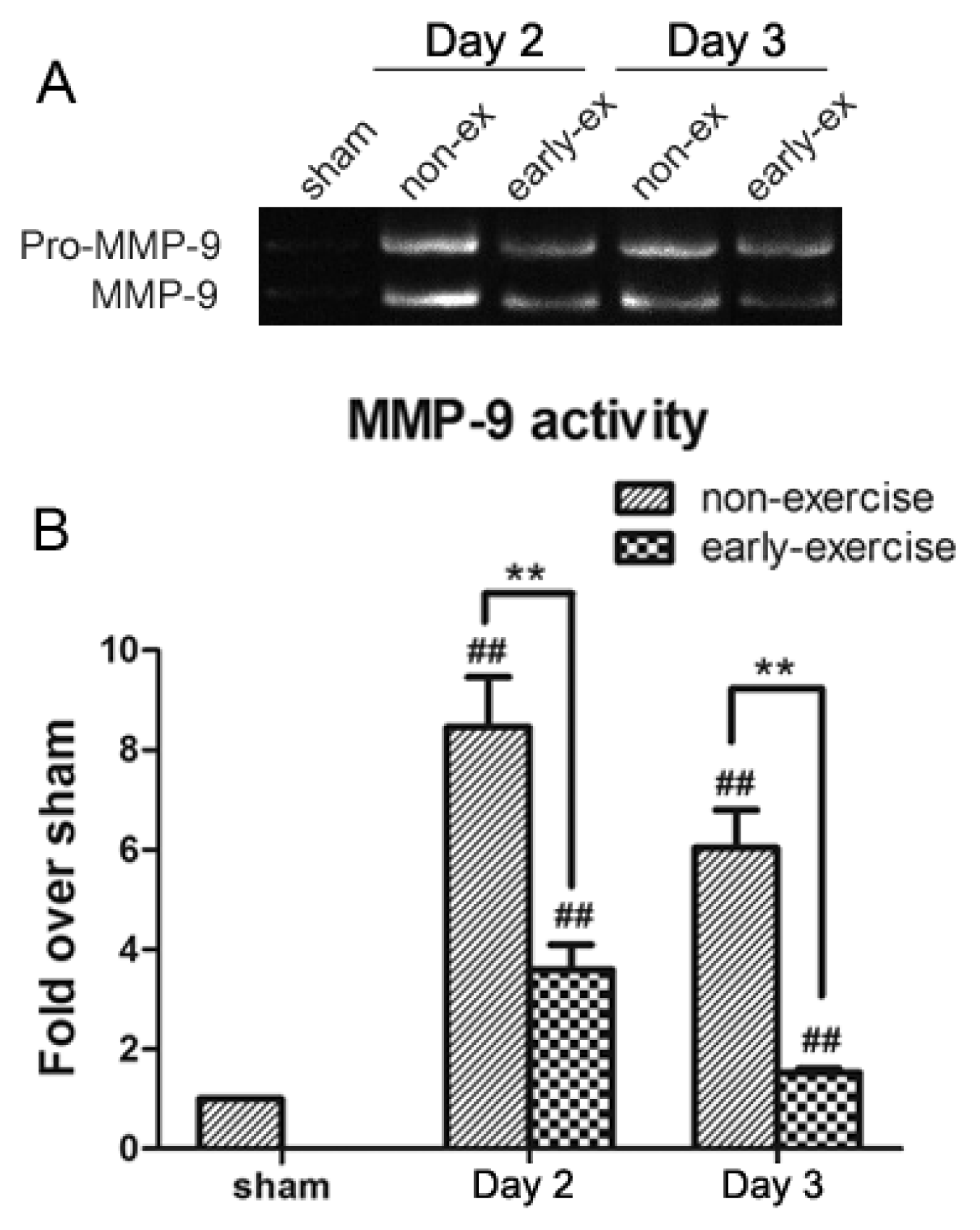
2.5. Early Exercise Inhibited the Increased Expression of MMP-9 Induced by MCAO
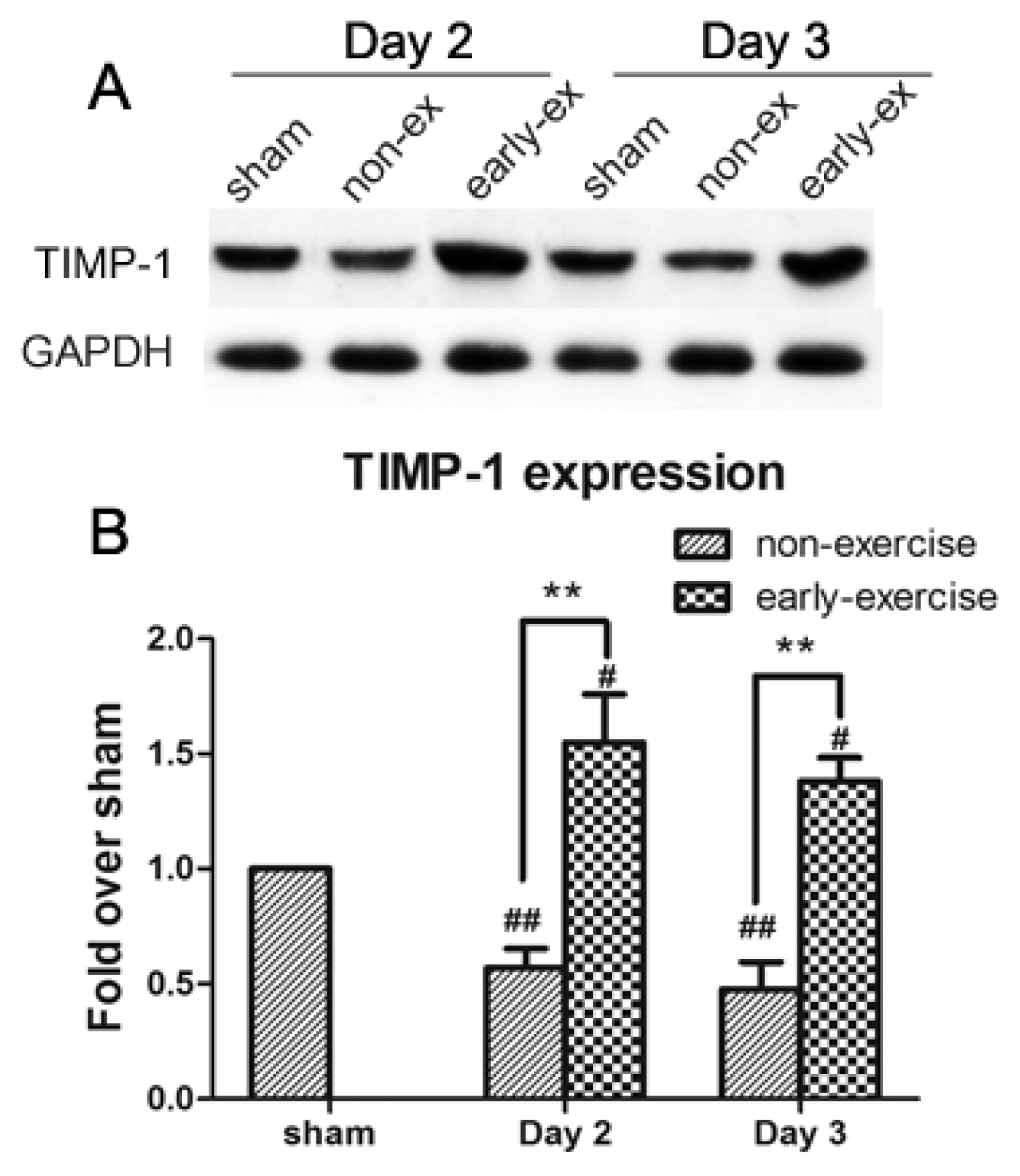
2.6. Early Exercise Promoted the Upregulation of TIMP-1 Expression
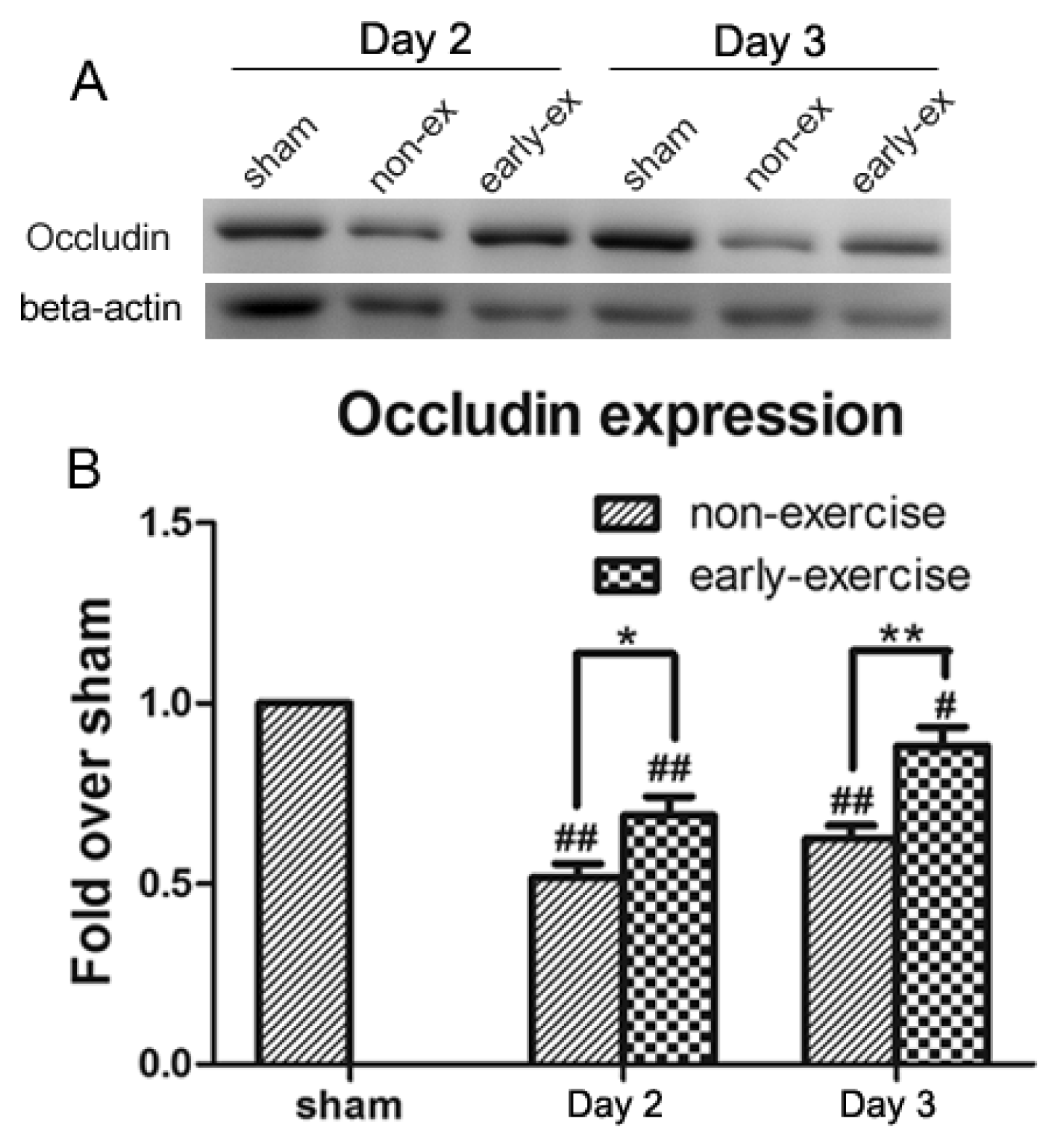
2.7. Early Exercise Suppressed Decreases in Occludin
2.8. Discussion
3. Experimental Section
3.1. Animals and Experimental Groups
3.2. Middle Cerebral Artery Occlusion (MCAO)
3.3. Treadmill Training
3.4. Assessment of Neurological Deficits
3.5. Measurement of Infarct Volume
3.6. Evaluation of Blood-Brain Barrier Permeability
3.7. Electron Microscopy
3.8. Western Blotting
3.9. Gelatin Zymography
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Yang, D.; Li, S.Y.; Yeung, C.M.; Chang, R.C.; So, K.F.; Wong, D.; Lo, A.C. Lycium barbarum extracts protect the brain from blood-brain barrier disruption and cerebral edema in experimental stroke. PLoS One 2012, 7, e33596. [Google Scholar]
- Ding, Y.H.; Ding, Y.; Li, J.; Bessert, D.A.; Rafols, J.A. Exercise pre-conditioning strengthens brain microvascular integrity in a rat stroke model. Neurol. Res 2006, 28, 184–189. [Google Scholar]
- Jin, R.; Yang, G.; Li, G. Molecular insights and therapeutic targets for blood-brain barrier disruption in ischemic stroke: Critical role of matrix metalloproteinases and tissue-type plasminogen activator. Neurobiol. Dis 2010, 38, 376–385. [Google Scholar]
- Perez de la Ossa, N.; Davalos, A. Neuroprotection in cerebral infarction: The opportunity of new studies. Cerebrovasc. Dis 2007, 24, 153–156. [Google Scholar]
- Huang, P.; Zhou, C.M.; Qin, H.; Liu, Y.Y.; Hu, B.H.; Chang, X.; Zhao, X.R.; Xu, X.S.; Li, Q.; Wei, X.H.; et al. Cerebralcare Granule(R) attenuates blood-brain barrier disruption after middle cerebral artery occlusion in rats. Exp. Neurol 2012, 237, 453–463. [Google Scholar]
- Wang, Z.; Leng, Y.; Tsai, L.K.; Leeds, P.; Chuang, D.M. Valproic acid attenuates blood-brain barrier disruption in a rat model of transient focal cerebral ischemia: The roles of HDAC and MMP-9 inhibition. J. Cereb. Blood Flow Metab 2011, 31, 52–57. [Google Scholar]
- Spatz, M. Past and recent BBB studies with particular emphasis on changes in ischemic brain edema. Acta. Neurochir. Suppl 2010, 106, 21–27. [Google Scholar]
- Abbott, N.J.; Friedman, A. Overview and introduction: The blood-brain barrier in health and disease. Epilepsia 2012, 53, 1–6. [Google Scholar]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J. Cereb. Blood Flow Metab 2007, 27, 697–709. [Google Scholar]
- Sandoval, K.E.; Witt, K.A. Blood-brain barrier tight junction permeability and ischemic stroke. Neurobiol. Dis 2008, 32, 200–219. [Google Scholar]
- Rosenberg, G.A.; Estrada, E.Y.; Dencoff, J.E. Matrix metalloproteinases and TIMPs are associated with blood-brain barrier opening after reperfusion in rat brain. Stroke 1998, 29, 2189–2195. [Google Scholar]
- Purvis, T.; Cadilhac, D.; Donnan, G.; Bernhardt, J. Systematic review of process indicators: Including early rehabilitation interventions used to measure quality of acute stroke care. Int. J. Stroke 2009, 4, 72–80. [Google Scholar]
- Zhang, F.; Wu, Y.; Jia, J. Exercise preconditioning and brain ischemic tolerance. Neuroscience 2011, 177, 170–176. [Google Scholar]
- Van Praag, H. Exercise and the brain: Something to chew on. Trends Neurosci 2009, 32, 283–290. [Google Scholar]
- Galvin, R.; Cusack, T.; O’Grady, E.; Murphy, T.B.; Stokes, E. Family-mediated exercise intervention (FAME): Evaluation of a novel form of exercise delivery after stroke. Stroke 2011, 42, 681–686. [Google Scholar]
- Matsuda, F.; Sakakima, H.; Yoshida, Y. The effects of early exercise on brain damage and recovery after focal cerebral infarction in rats. Acta Physiol 2010, 201, 275–287. [Google Scholar]
- Yang, Y.R.; Wang, R.Y.; Wang, P.S. Early and late treadmill training after focal brain ischemia in rats. Neurosci. Lett 2003, 339, 91–94. [Google Scholar]
- Chang, H.C.; Yang, Y.R.; Wang, S.G.; Wang, R.Y. Effects of treadmill training on motor performance and extracellular glutamate level in striatum in rats with or without transient middle cerebral artery occlusion. Behav. Brain Res 2009, 205, 450–455. [Google Scholar]
- Berchtold, N.C.; Castello, N.; Cotman, C.W. Exercise and time-dependent benefits to learning and memory. Neuroscience 2010, 167, 588–597. [Google Scholar]
- Zhang, P.; Zhang, Q.; Pu, H.; Wu, Y.; Bai, Y.; Vosler, P.S.; Chen, J.; Shi, H.; Gao, Y.; Hu, Y. Very early-initiated physical rehabilitation protects against ischemic brain injury. Front. Biosci 2012, 4, 2476–2489. [Google Scholar]
- Zheng, Q.; Zhu, D.; Bai, Y.; Wu, Y.; Jia, J.; Hu, Y. Exercise improves recovery after ischemic brain injury by inducing the expression of angiopoietin-1 and Tie-2 in rats. Tohoku J. Exp. Med 2011, 224, 221–228. [Google Scholar]
- Zhang, Q.; Wu, Y.; Zhang, P.; Sha, H.; Jia, J.; Hu, Y.; Zhu, J. Exercise induces mitochondrial biogenesis after brain ischemia in rats. Neuroscience 2011, 205, 10–17. [Google Scholar]
- Zhang, Q.; Wu, Y.; Sha, H.; Zhang, P.; Jia, J.; Hu, Y.; Zhu, J. Early exercise affects mitochondrial transcription factors expression after cerebral ischemia in rats. Int. J. Mol. Sci 2012, 13, 1670–1679. [Google Scholar]
- Zhang, P.; Yu, H.; Zhou, N.; Zhang, J.; Wu, Y.; Zhang, Y.; Bai, Y.; Jia, J.; Zhang, Q.; Tian, S.; et al. Early exercise improves cerebral blood flow through increased angiogenesis in experimental stroke rat model. J. Neuroeng. Rehabil 2013, 10, 43, :1–43:10.. [Google Scholar]
- Zhang, P.; Zhang, Y.; Zhang, J.; Wu, Y.; Jia, J.; Wu, J.; Hu, Y. Early exercise protects against cerebral ischemic injury through inhibiting neuron apoptosis in cortex in rats. Int. J. Mol. Sci 2013, 14, 6074–6089. [Google Scholar]
- Sood, R.R.; Taheri, S.; Candelario-Jalil, E.; Estrada, E.Y.; Rosenberg, G.A. Early beneficial effect of matrix metalloproteinase inhibition on blood-brain barrier permeability as measured by magnetic resonance imaging countered by impaired long-term recovery after stroke in rat brain. J. Cereb. Blood Flow Metab 2008, 28, 431–438. [Google Scholar]
- Tavelin, S.; Hashimoto, K.; Malkinson, J.; Lazorova, L.; Toth, I.; Artursson, P. A new principle for tight junction modulation based on occludin peptides. Mol. Pharmacol 2003, 64, 1530–1540. [Google Scholar]
- Enzinger, C.; Dawes, H.; Johansen-Berg, H.; Wade, D.; Bogdanovic, M.; Collett, J.; Guy, C.; Kischka, U.; Ropele, S.; Fazekas, F.; et al. Brain activity changes associated with treadmill training after stroke. Stroke 2009, 40, 2460–2467. [Google Scholar]
- Liu, N.; Huang, H.; Lin, F.; Chen, A.; Zhang, Y.; Chen, R.; Du, H. Effects of treadmill exercise on the expression of netrin-1 and its receptors in rat brain after cerebral ischemia. Neuroscience 2011, 194, 349–358. [Google Scholar]
- Quirie, A.; Hervieu, M.; Garnier, P.; Demougeot, C.; Mossiat, C.; Bertrand, N.; Martin, A.; Marie, C.; Prigent-Tessier, A. Comparative effect of treadmill exercise on mature BDNF production in control versus stroke rats. PLoS One 2012, 7, e44218. [Google Scholar]
- Langhammer, B.; Lindmark, B. Functional exercise and physical fitness post stroke: The importance of exercise maintenance for motor control and physical fitness after stroke. Stroke Res. Treat 2012, 2012, 864835. [Google Scholar]
- Brethour, M.K.; Nystrom, K.V.; Broughton, S.; Kiernan, T.E.; Perez, A.; Handler, D.; Swatzell, V.; Yang, J.J.; Starr, M.; Seagraves, K.B.; et al. Controversies in acute stroke treatment. AACN Adv. Crit. Care 2012, 23, 158–172, ; quiz 173–174.. [Google Scholar]
- Risedal, A.; Zeng, J.; Johansson, B.B. Early training may exacerbate brain damage after focal brain ischemia in the rat. J. Cereb. Blood Flow Metab 1999, 19, 997–1003. [Google Scholar]
- Dromerick, A.W.; Lang, C.E.; Birkenmeier, R.L.; Wagner, J.M.; Miller, J.P.; Videen, T.O.; Powers, W.J.; Wolf, S.L.; Edwards, D.F. Very early constraint-induced movement during stroke rehabilitation (VECTORS): A single-center RCT. Neurology 2009, 73, 195–201. [Google Scholar]
- Bland, S.T.; Pillai, R.N.; Aronowski, J.; Grotta, J.C.; Schallert, T. Early overuse and disuse of the affected forelimb after moderately severe intraluminal suture occlusion of the middle cerebral artery in rats. Behav. Brain Res 2001, 126, 33–41. [Google Scholar]
- Bland, S.T.; Schallert, T.; Strong, R.; Aronowski, J.; Grotta, J.C.; Feeney, D.M. Early exclusive use of the affected forelimb after moderate transient focal ischemia in rats: Functional and anatomic outcome. Stroke 2000, 31, 1144–1152. [Google Scholar]
- Lee, S.U.; Kim, D.Y.; Park, S.H.; Choi, D.H.; Park, H.W.; Han, T.R. Mild to moderate early exercise promotes recovery from cerebral ischemia in rats. Can. J. Neurol. Sci 2009, 36, 443–449. [Google Scholar]
- Yang, Y.R.; Chang, H.C.; Wang, P.S.; Wang, R.Y. Motor performance improved by exercises in cerebral ischemic rats. J. Mot. Behav 2012, 44, 97–103. [Google Scholar]
- Bernhardt, J.; Dewey, H.; Thrift, A.; Collier, J.; Donnan, G. A very early rehabilitation trial for stroke (AVERT): Phase II safety and feasibility. Stroke 2008, 39, 390–396. [Google Scholar]
- Bernhardt, J.; Indredavik, B.; Langhorne, P. When should rehabilitation begin after stroke? Int. J. Stroke 2013, 8, 5–7. [Google Scholar]
- Tian, S.; Bai, Y.; Yang, L.; Wang, X.; Wu, Y.; Jia, J.; Zhu, Y.; Cheng, Y.; Zhang, P.; Wu, J.; et al. Shear stress inhibits apoptosis of ischemic brain microvascular endothelial cells. Int. J. Mol. Sci 2013, 14, 1412–1427. [Google Scholar]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis 2010, 37, 13–25. [Google Scholar]
- Khan, M.; Dhammu, T.S.; Sakakima, H.; Shunmugavel, A.; Gilg, A.G.; Singh, A.K.; Singh, I. The inhibitory effect of S-nitrosoglutathione on blood-brain barrier disruption and peroxynitrite formation in a rat model of experimental stroke. J. Neurochem 2012, 123, 86–97. [Google Scholar]
- Cunningham, L.A.; Wetzel, M.; Rosenberg, G.A. Multiple roles for MMPs and TIMPs in cerebral ischemia. Glia 2005, 50, 329–339. [Google Scholar]
- Barr, T.L.; Latour, L.L.; Lee, K.Y.; Schaewe, T.J.; Luby, M.; Chang, G.S.; El-Zammar, Z.; Alam, S.; Hallenbeck, J.M.; Kidwell, C.S.; et al. Blood-brain barrier disruption in humans is independently associated with increased matrix metalloproteinase-9. Stroke 2010, 41, e123–e128. [Google Scholar]
- Cojocarui, I.M.; Cojocaru, M.; Sapira, V.; Socoliuc, G.; Hertea, C.; Paveliu, S. Changes in plasma matrix metalloproteinase-9 levels in patients with acute ischemic stroke. Rom. J. Intern. Med 2012, 50, 155–158. [Google Scholar]
- Guo, M.; Cox, B.; Mahale, S.; Davis, W.; Carranza, A.; Hayes, K.; Sprague, S.; Jimenez, D.; Ding, Y. Pre-ischemic exercise reduces matrix metalloproteinase-9 expression and ameliorates blood-brain barrier dysfunction in stroke. Neuroscience 2008, 151, 340–351. [Google Scholar]
- Yang, Y.; Rosenberg, G.A. Blood-brain barrier breakdown in acute and chronic cerebrovascular disease. Stroke 2011, 42, 3323–3328. [Google Scholar]
- Cheng, T.; Petraglia, A.L.; Li, Z.; Thiyagarajan, M.; Zhong, Z.; Wu, Z.; Liu, D.; Maggirwar, S.B.; Deane, R.; Fernandez, J.A.; et al. Activated protein C inhibits tissue plasminogen activator-induced brain hemorrhage. Nat. Med 2006, 12, 1278–1285. [Google Scholar]
- Zhang, G.S.; Tian, Y.; Huang, J.Y.; Tao, R.R.; Liao, M.H.; Lu, Y.M.; Ye, W.F.; Wang, R.; Fukunaga, K.; Lou, Y.J.; et al. The gamma-secretase blocker DAPT reduces the permeability of the blood-brain barrier by decreasing the ubiquitination and degradation of occludin during permanent brain ischemia. CNS Neurosci. Ther 2012, 19, 53–60. [Google Scholar]
- Persidsky, Y.; Ramirez, S.H.; Haorah, J.; Kanmogne, G.D. Blood-brain barrier: Structural components and function under physiologic and pathologic conditions. J. Neuroimmune. Pharmacol 2006, 1, 223–236. [Google Scholar]
- Liu, W.; Hendren, J.; Qin, X.J.; Shen, J.; Liu, K.J. Normobaric hyperoxia attenuates early blood-brain barrier disruption by inhibiting MMP-9-mediated occludin degradation in focal cerebral ischemia. J. Neurochem 2009, 108, 811–820. [Google Scholar]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar]
- Rogers, D.C.; Campbell, C.A.; Stretton, J.L.; Mackay, K.B. Correlation between motor impairment and infarct volume after permanent and transient middle cerebral artery occlusion in the rat. Stroke 1997, 28, 2060–2065, ; discussion 2066.. [Google Scholar]
- Yu, Q.; Chu, M.; Wang, H.; Lu, S.; Gao, H.; Li, P.; Gan, Y.; Shi, H.; Liang, W.; Chen, J.; et al. Sevoflurane preconditioning protects blood-brain-barrier against brain ischemia. Front. Biosci 2011, 3, 978–988. [Google Scholar]








© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Y.; Zhang, P.; Shen, X.; Tian, S.; Wu, Y.; Zhu, Y.; Jia, J.; Wu, J.; Hu, Y. Early Exercise Protects the Blood-Brain Barrier from Ischemic Brain Injury via the Regulation of MMP-9 and Occludin in Rats. Int. J. Mol. Sci. 2013, 14, 11096-11112. https://doi.org/10.3390/ijms140611096
Zhang Y, Zhang P, Shen X, Tian S, Wu Y, Zhu Y, Jia J, Wu J, Hu Y. Early Exercise Protects the Blood-Brain Barrier from Ischemic Brain Injury via the Regulation of MMP-9 and Occludin in Rats. International Journal of Molecular Sciences. 2013; 14(6):11096-11112. https://doi.org/10.3390/ijms140611096
Chicago/Turabian StyleZhang, Yuling, Pengyue Zhang, Xiafeng Shen, Shan Tian, Yi Wu, Yulian Zhu, Jie Jia, Junfa Wu, and Yongshan Hu. 2013. "Early Exercise Protects the Blood-Brain Barrier from Ischemic Brain Injury via the Regulation of MMP-9 and Occludin in Rats" International Journal of Molecular Sciences 14, no. 6: 11096-11112. https://doi.org/10.3390/ijms140611096
APA StyleZhang, Y., Zhang, P., Shen, X., Tian, S., Wu, Y., Zhu, Y., Jia, J., Wu, J., & Hu, Y. (2013). Early Exercise Protects the Blood-Brain Barrier from Ischemic Brain Injury via the Regulation of MMP-9 and Occludin in Rats. International Journal of Molecular Sciences, 14(6), 11096-11112. https://doi.org/10.3390/ijms140611096