Enhancement of Immune Activation Activities of Spirulina maxima Grown in Deep-Sea Water

Abstract
:1. Introduction
2. Results and Discussion
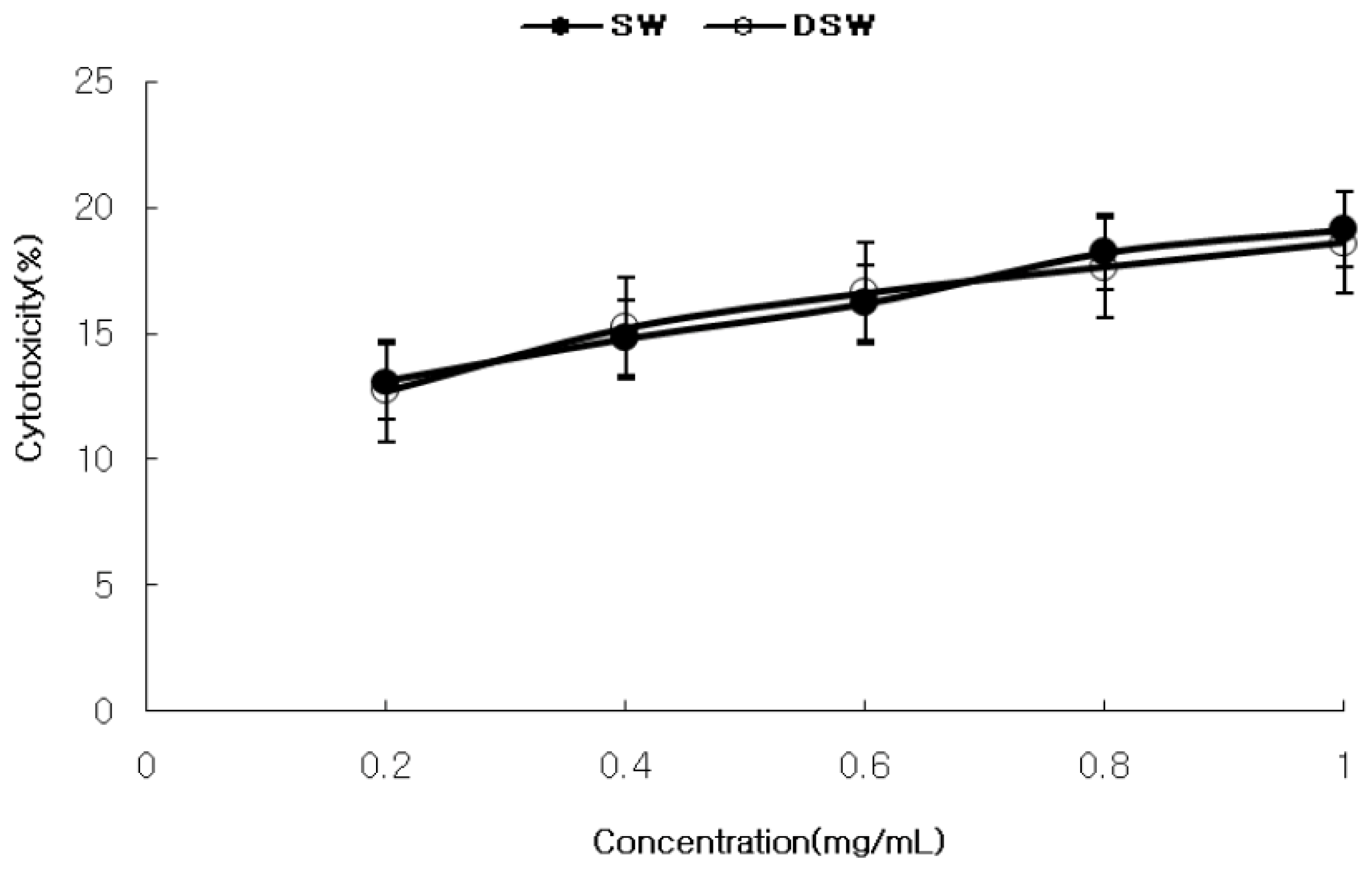
2.1. Measurement of Cytotoxicity and Anticancer Activity
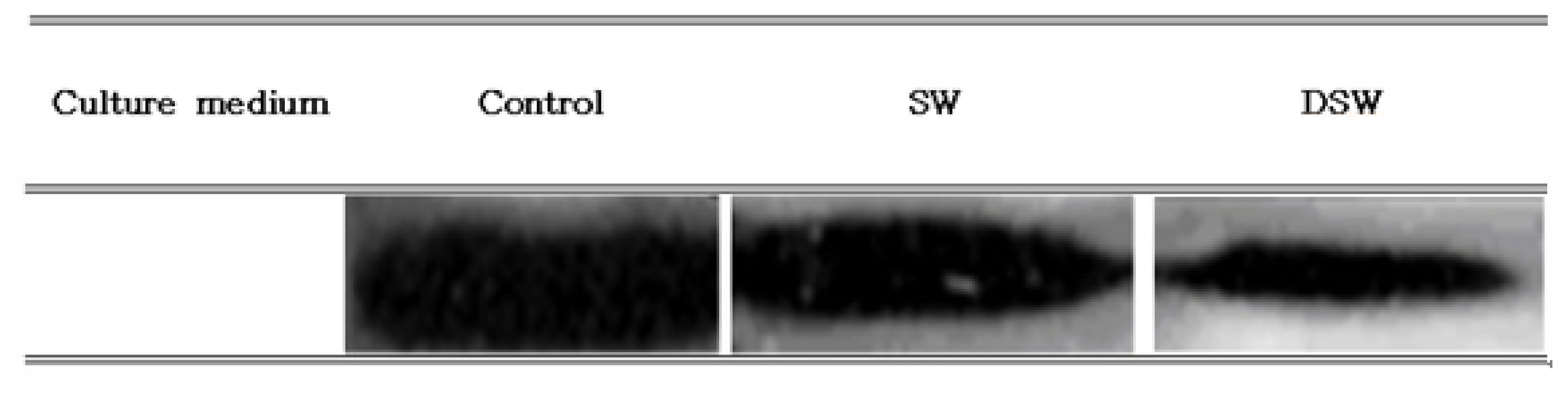
2.2. Measurement of Bcl-2 Expression in Human Cancer Cell
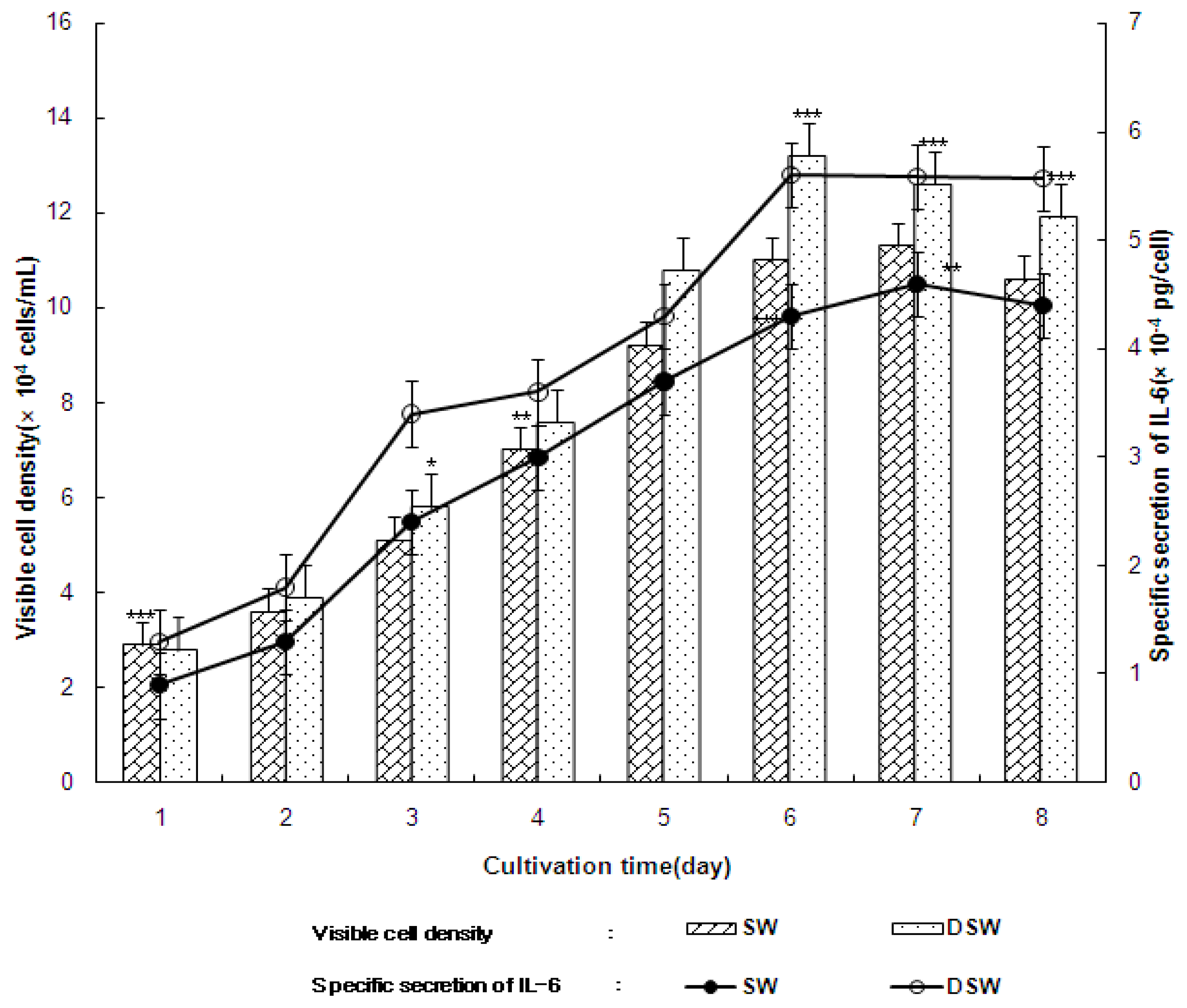
2.3. Secretion of Cytokine, Interleukin-6 (IL-6), and Tumor Nucreosis Factor-α (TNF-α) from the Growth of Human B and T Cells
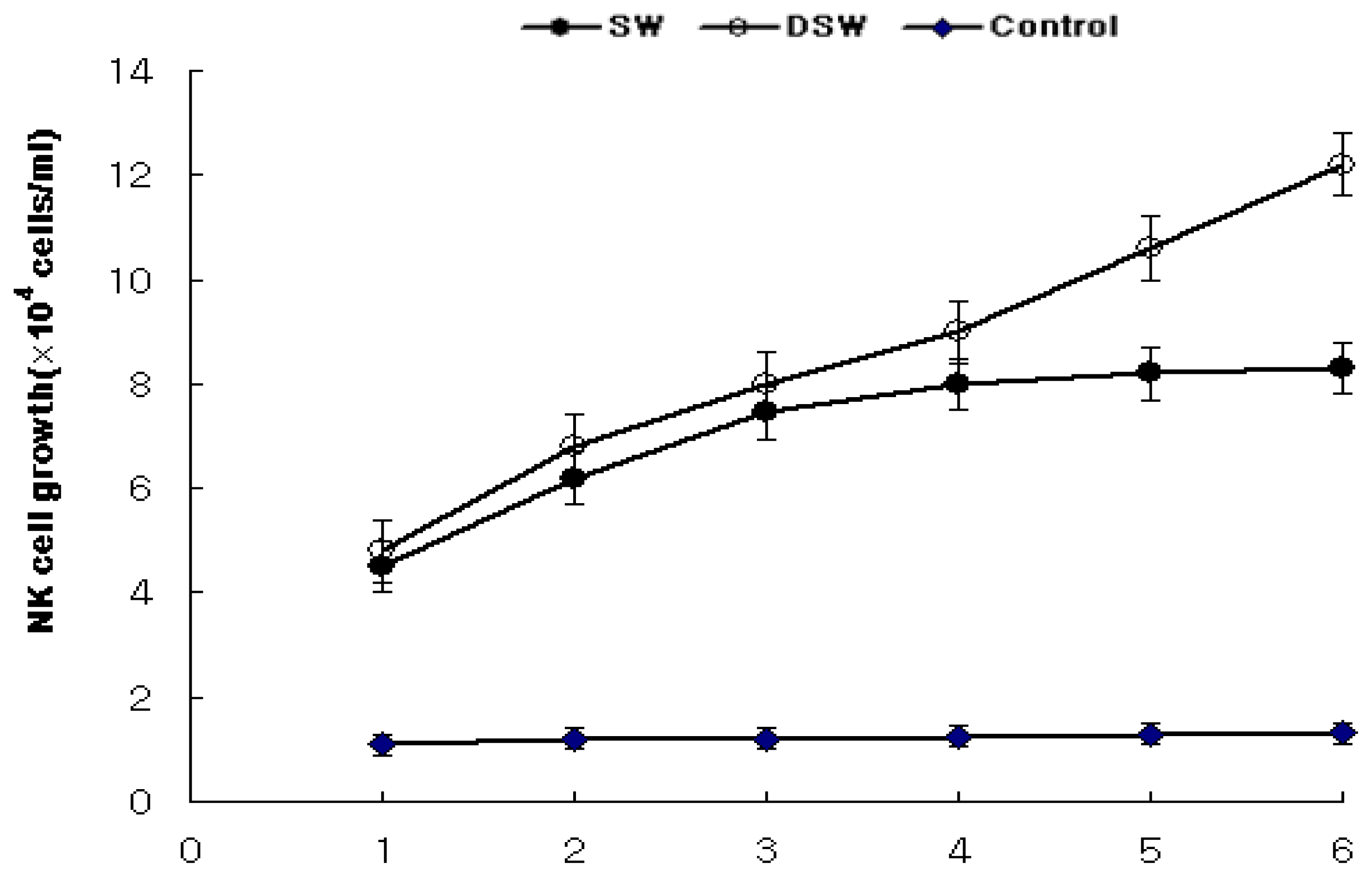
2.4. Enhancement of NK Cells Associated with Human T Cell Growth
2.5. HPLC Analysis
2.6. Discussion
3. Experimental Section
3.1. Cells and Chemicals
3.2. Culture Condition and Extraction of S. maxima
3.3. Measurement of Cytotoxicity and Anticancer Activity
3.4. Measurement of Bcl2 Protein Level
3.5. Measurement of Human T and B Cell Growth and Secretion of Cytokines
3.6. Enhancement of Human Natural Killer (NK) Cell Growth
3.7. HPLC Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Oh, S.H.; Han, J.G.; Ha, J.H.; Kim, Y.; Jeong, M.H.; Kim, S.S.; Jeong, H.S.; Choi, G.P.; Park, U.Y.; Kang, D.H.; et al. Enhancement of immune activity of Spirulina maxima by low temperature ultrasonification extraction. Korean J. Food Sci. Technol 2009, 41, 313–319. [Google Scholar]
- Jassby, A. Spirulina: A Model for Microalgae as Human Food. In Algae and Human Affair; Cambridge University Press: Cambridge, UK, 1998; pp. 65–67. [Google Scholar]
- Ayehunie, S.; Belay, A.; Hu, Y.; Baba, T.; Ruprecht, R. Inhibition of HIV-1 replication by an aqueous extract of Spirulina platensis (Arthrospira platensis). J. Acquir. Immune. Defic. Syndr. Hum. Retrovirol 1996, 18, 7–12. [Google Scholar]
- Babu, M. Evaluation of chemoprevention of oral cancer with Spirulina fusiformis. Nutr. Cancer 1995, 24, 197–202. [Google Scholar]
- Kim, H.S.; Kim, C.H.; Kwon, M.C.; Song, Y.K.; Cho, J.H.; Gwak, H.G.; Hwang, B.Y.; Kim, J.C.; Lee, H.Y. Anticancer activity of ultrasonified extracts from seawater-based culture of the microalga Spirulina platensis. Korean J. Fish. Soc 2006, 39, 318–325. [Google Scholar]
- Tomohiro, H.; Misako, M.; Kaoru, H.; Yoshiko, S.; Michio, U.; Tsukasa, S. Activation of the human innate immune system by Spirulina: Augmentation of interferon production and NK cytotoxicity by oral administration of hot water extract of Spirulina platensis. Int. Immunopharmacol 2002, 2, 423–434. [Google Scholar]
- Mohamed, F.I.; Doaa, A.A.; Augusta, F.; Mohamed, E.A.; Rajiv, L.G.; Wael, M.I.; Madhwa, H.R.; Allal, O. Chemoprevention of rat liver toxicity and carcinogenesis by Spirulina. Int. J. Biol. Sci 2009, 5, 377–387. [Google Scholar]
- Rathore, N.K.; Singh, S.; Singh, V. Spirulina as immuno-enhancer and biomodulator. J. Immunol. Immunopathol 2004, 6, 1–10. [Google Scholar]
- Mazo, V.K.; Gmoshinshii, I.V.; Zilova, I.S. Microalgae Spirulina in human nutrition. Vopr. Pitan 2004, 73, 45–53. [Google Scholar]
- Kay, R.A. Microalgae as food and supplement. Crit. Rev. Food Sci. Nutr 1991, 30, 555–573. [Google Scholar]
- Yang, H.N.; Lee, E.H.; Kim, H.M. Spirulina platensis inhibits anaphylactic reaction. Life Sci 1997, 61, 1237–1244. [Google Scholar]
- Hernandez, A.C.; Nieves, I.; Meckes, M.; Chamorro, G.; Barron, B.L. Antiviral activity of Spirulina maxima against Herpes simplex virus type 2. Antiviral Res 2002, 56, 297–285. [Google Scholar]
- Jeong, S.W.; Lee, N.K.; Kim, S.J.; Han, D.S. Screening of tyrosinase inhibitor from plants. Korean J. Food Sci. Technol 1995, 27, 891–896. [Google Scholar]
- Kim, I.S; Cho, Z.S. Modulation of human fibroblast proliferation and collegan production by Prostaglandin E2. Korean Biochem. J 1993, 26, 40–52. [Google Scholar]
- Cho, J.H.; Lee, K.M.; Kim, N.S; Kang, W.H. The effects of whitening components on human melanocytes in vitro. Korean Cosmet. Sci 1997, 23, 115–121. [Google Scholar]
- Fukami, K.; Nishijima, T.; Hata, Y. Availability of deep-sea water and effects of bacteria isolated from deep-sea water on the mass culture of food microalga Chaetoceros ceratosporum. Nippon Suisan Gakkaishi 1992, 58, 931–936. [Google Scholar]
- Nakasone, T.; Akeda, S. The application of deep-sea water in Japan. Nakasone Akeda. 1997, 69–75. [Google Scholar]
- Fukami, K.; Nishimura, S.; Ogusa, M.; Asada, M.; Nishijima, T. Continuous culture with deep-sea water of a benthic food diatom Nitzschia sp. Hydrobiologia 1997, 358, 245–249. [Google Scholar]
- Roger, H. Chemical exchange between sea water and deep ocean basalts. Earth Planet. Sci. Lett 1970, 9, 269–279. [Google Scholar]
- Quinby-Hunt, M.S.; Turehian, K.K. Distribution of elements in sea water. Eos Trans. Am. Geophys. Union 1983, 64, 130–135. [Google Scholar]
- Berger, W.H. Deep-sea carbonate: Pteropod distribution and the aragonite compensation depth. Deep Sea Res 1978, 25, 447–452. [Google Scholar]
- Hwang, H.S.; Kim, S.H.; Yoo, Y.G.; Chu, Y.S.; Shon, Y.H.; Nam, K.S.; Yun, J.W. Inhibitory effect of deep-sea water on differentiation of 3T3-L1 adipocytes. Mar. Biotechnol 2009, 11, 161–168. [Google Scholar]
- Hataguchi, Y.; Tai, H.; Nakajima, H.; Kimata, H. Drinking deep-sea water restores mineral imbalance in atopic eczema/dermatitis syndrome. Eur. J. Clin. Nutr 2005, 59, 1093–1096. [Google Scholar]
- Fusako, M.; Kyoko, M.; Taeko, N.; Nao, M.; Yuhsuke, S. Effects of red algae cultivated with deep-sea water on the oxidation-reduction status of liver, lung, brain, and bone metabolism in SAMP1 and SAMR1. Int. Congr. Ser 2003, 1260, 413–416. [Google Scholar]
- Lingling, Y.; Yao, W.; Qingjun, Z.; Peng, C.; Yiqiang, W.; Ye, W.; Ting, L.; Lixin, X. Inhibitory effects of polysaccharide extract from Spirulina platensis on corneal neovascularization. Mol. Vis 1991, 15, 1951–1961. [Google Scholar]
- Joel, S.; Gerald, S. Regression of experimental hamster cancer by beta carotene and algae extracts. J. Oral. Maxillofac. Surg 1987, 45, 510–515. [Google Scholar]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar]
- Youn, C.K.; Cho, H.J.; Kim, S.H.; Kim, H.B.; Kim, M.H.; Chang, I.Y.; Lee, J.S.; Chung, M.H.; Hahm, K.S.; You, H.J. Bcl-2 expression suppresses mismatch repair activity through inhibition of E2F transcriptional activity. Nat. Cell Biol 2005, 7, 137–147. [Google Scholar]
- Hwang, C.S.; Choi, J.O.; Kim, I.S. Detection of bcl-2 & bax Expression in the Head & Neck Cancers. Cancer Res. Treat 1995, 27, 570–577. [Google Scholar]
- Yim, J.H.; Son, E.; Pyo, S.; Lee, H.K. Novel sulfated polysaccharide derived from red-tide microalgae Gyrodinium impudicum strain KG03 with immunostimulating activity in vivo. Mar. Biotechnol 2005, 7, 1436–2228. [Google Scholar]
- Liu, R.; van-Kaer, L.; La-Cava, La.; Price, M.; Campagnolo, D.I.; Collins, M.; Young, D.A.; Vollmer, T.L.; Shp, F.D. Autoreactive T cells mediate NK cell degeneration in autoimmune disease. J. Immunol 2006, 176, 5247–5254. [Google Scholar]
- See, D.M.; Broumand, N.; Sahl, L.; Tilles, J.G. In vitro effects of echinacea and ginseng on natural killer and antibody-dependent cell cytotoxicity in healthy subjects and chronic fatigue syndrome or acquired immunodeficiency syndrome patients. Immunopharmacology 1997, 35, 229–235. [Google Scholar]
- Abd El-Baky, H.H.; El Baz, F.K.; El-Baroty, G.S. Spirulina Species as a source of carotenoids and α-tocopherol and its anticarcinoma factors. Biotechnol 2003, 2, 222–240. [Google Scholar]
- Abd El-Baky, H.H.; El Baz, F.K.; El-Baroty, G.S. Production of antioxidant by the green alga Dunaliella salina. Int. J. Agric. Biol 2004, 6, 49–57. [Google Scholar]
- Bendich, A. Non vitamin A activity of carotenoids: Immuno enhancement. Trends in Food Sci. Technol. 1991, 127–129. [Google Scholar]
- Aboul-Enein, A.M.; El Baz, F.K.; El-Baroty, G.S.; Youssef, A.M.; Abd El-Baky, H.H. Antioxidant activity of algal extracts on lipid peroxidation. J. Med. Sci 2003, 3, 87–98. [Google Scholar]
- Poppel, G.V.; Goldbohm, R.A. Epidemiological evidences for β-carotene and cancer prevention. Am. J. Clin. Nut 1995, 62, 1393S–1402S. [Google Scholar]
- Schwartz, J.L. The dual role of nutrients as antioxidants and peroxidants: Their effects on tumor cell growth. J. Nutr 1996, 126, 12215–12275. [Google Scholar]
- Packer, L.; Glazer, A.N. Methods in Enzymology. In Cyanobacteria; Academic Press: San Diego, CA, USA, 1988. [Google Scholar]
- Aristo, V.; Mamdooh, G. In vivo effect of ascorbic acid on enhancement of human natural killer cell activity. Nutr. Res 1993, 13, 753–764. [Google Scholar]
- Abd El-Baky, H.H.; El Baz, F.K.; El-Baroty, G.S. Production of carotenoids from marine microalgae and its evaluation as food colorant and lowering cholesterol agents. Am. Eurasian J. Agric. Environ. Sci 2007, 2, 792–800. [Google Scholar]
- Fukami, K.; Nishijima, T.; Ishida, Y. Stimulative and inhibitory effects of bacteria on the growth of microalgae. Hydrobiologia 1997, 358, 185–191. [Google Scholar]
- Henson, D.E.; Block, G.; Levine, M. Ascorbic acids: Biologic functions and relation to cancer. J. Natl. Cancer Inst 1991, 83, 547–550. [Google Scholar]
- Lee, J.J.; Shin, D.H.; Park, S.E.; Kim, W.I.; Park, D.I.; Choi, Y.H.; Hong, S.H. Euphorbiae humifusae sensitizes apoptosis of TRAIL-resistant human gastric adenocarcinoma AGS cells. J. Life Sci 2008, 18, 120–128. [Google Scholar]
- Thomas, W.D.; Hersey, P. TNF-related apoptosis-inducing ligand (TRAIL) induces apoptosis in Fas ligand-resistant melanoma cells and mediates CD4 T cell killing of target cells. J. Immunol 1998, 161, 2195–2200. [Google Scholar]
- Qadir, S.A.; Kwon, M.C.; Han, J.G.; Ha, J.H.; Chung, H.S.; Ahnm, J.; Lee, H.Y. Effect of different extraction protocols on anticancer and antioxidant activities of Berberis koreana bark extracts. J. Biosci. Bioeng 2009, 107, 331–338. [Google Scholar]
- Femenia, A.; Garcia-Marin, M.; Simal, S.; Rossello, C.; Blasco, M. Effects of supercritical carbon dioxide (SC-CO2) oil extraction on the cell wall composition of almond fruits. J. Agric. Food Chem 2001, 49, 5828–5834. [Google Scholar]
- Hamburger, M.; Baumann, D.; Adler, S. Supercritical carbon dioxide extraction of selected medical plants—Effects of high pressure and added ethanol on yield of extracted substances. Phytochem. Anal 2004, 15, 46–54. [Google Scholar]
- Kaufmann, B.; Christen, P. Recent extraction techniques for natural products: Microwave-assisted extraction and pressurised solvent extraction. Phytochem. Anal 2002, 13, 105–113. [Google Scholar]
- Sharma, U.K.; Sharma, K.; Sharma, N.; Sharma, A.; Singh, H.P.; Sinha, A.K. Microwave-assisted efficient extraction of different parts of Hippophae rhamnoides for the comparative evaluation of antioxidant activity and quantification of Its phenolic constituents by reverse-phase high-performance liquid chromatography (RP-HPLC). J. Agric. Food Chem 2008, 56, 374–379. [Google Scholar]
- Zhang, S.; Zhang, J.; Wang, C. Extraction of steroid saponins from Paris polyphylla Sm. var. yunnanensis using novel ultrahigh pressure extraction technology. Pharm. Chem. J 2007, 41, 424–429. [Google Scholar]
- Samson, R.; Leduy, A. Multistage continuous cultivation of blue-green alga Spirulina maxima in the flat tank photobioreactors with recycle. Can. J. Chem. Eng 1985, 63, 105–112. [Google Scholar]
- Abd El-Baky, H.H.; El Baz, F.K.; El-Baroty, G.S. Production of phenolic compounds from Spirulina maxima microalgae and its protective effects in vitro toward hepatotoxicity model. Afr. J. Pharm. Pharmacol 2009, 3, 133–139. [Google Scholar]
- Doll, R.; Peto, R. The causes of cancer, quantitative estimates of avoidable risks of cancer in the United States today. J. Natl. Cancer Inst 1981, 66, 1192–1308. [Google Scholar]
- Li, H.L.; Chen, D.D.; Li, X.H.; Zhang, H.W.; Lu, Y.Q.; Ye, C.L.; Ren, X.D. Changes of NF-κB, p53, Bcl-2 and caspase in apoptosis induced by JTE-522 in human gastric adenocarcinoma cell line AGS cells: role of reactive oxygen species. World J. Gastroenterol 2002, 8, 431–435. [Google Scholar]
- Laemmli, U.K. Cleavage if structural proteins during assembly of the head of bacteriophasfe T4. Nature 1970, 227, 680–685. [Google Scholar]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide fels to nitrocellulose sheets: Procedures and some applications. Proc. Natl. Acad. Sci. USA 1976, 76, 4350–4354. [Google Scholar]
- Lee, S.H.; Lee, H.S.; Park, Y.S.; Hwang, B.; Kim, J.H.; Lee, H.Y. Screening of immune activation activities in the leaves of Dendropanax morbifera Lev. Hanguk Yakyong Changmul Hakhoe Chi 2002, 10, 109–115. [Google Scholar]
- Ha, J.H.; Kwon, M.C.; Han, J.G.; Jin, L.; Jeong, H.S.; Choi, G.P.; Park, U.Y.; You, S.G.; Lee, H.Y. Enhancement of immunomodulatory activities of low molecular weight fucoidan isolated from Hizikia fusiforme. Korean J. Food Sci. Technol 2008, 40, 545–550. [Google Scholar]
- Yueran, Z.; Rui, S.; You, L.; Chunyi, G.; Zhigang, T. Expression of leptin receptors and response to leptin stimulation of human natural killer cell lines. Biochem. Biophys. Res. Commun 2003, 300, 247–252. [Google Scholar]
- Limdbolum, C.K. IL-2 receptor signaling through the Shb adapter protein in T and NK cells. Biochem. Biophys. Res. Commun 2002, 296, 929–936. [Google Scholar]
- Driskell, W.J.; Bashor, M.M.; Neese, J.W. Beta-carotene determined in serum by liquid chromatography with an internal standard. Clin. Chem 1983, 29, 1042–1044. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (mg/mL) | Seawater (SW) | Deep-sea water (DSW) | ||
|---|---|---|---|---|
| MCF-7 | Hep3B | MCF-7 | Hep3B | |
| 0.2 | 43.5 ± 0.22 | 44.2 ± 0.22 *** | 46.5 ± 0.23 *** | 45.3 ± 0.40 ** |
| 0.4 | 52.1 ± 0.23 * | 55.6 ± 0.24 | 56.1 ± 0.43 | 59.4 ± 0.47 |
| 0.6 | 61.7 ± 0.20 | 62.4 ± 0.23 * | 68.2 ± 0.26 ** | 74.4 ± 0.44 * |
| 0.8 | 73.4 ± 0.27 ** | 75.1 ± 0.28 | 77.5 ± 0.39 | 82.1 ± 0.41 *** |
| 1.0 | 80.5 ± 0.41 *** | 83.4 ± 0.31 ** | 89.6 ± 0.42 * | 93.5 ± 0.43 ** |
| Sample | Cultivation time (Day) | T cell | B cell | ||||
|---|---|---|---|---|---|---|---|
| Viable cell density (×104 cells/mL) | Cytokine secretion (×10−4 pg/cell) | Viable cell density (×104 cells/mL) | Cytokine secretion (×10−4 pg/cell) | ||||
| IL-6 | TNF-α | IL-6 | TNF-α | ||||
| DSW | 1 | 2.9 ± 0.02 * | 1.2 ± 0.16 *** | 1.3 ± 0.11 * | 2.8 ± 0.07 ** | 1.3 ± 0.01 ** | 1.4 ± 0.06 * |
| 2 | 4.2 ± 0.04 ** | 1.6 ± 0.07 *** | 1.5 ± 0.03 *** | 3.9 ± 0.06 * | 1.8 ± 0.15 *** | 1.7 ± 0.04 *** | |
| 3 | 6.5 ± 0.05 * | 3.2 ± 0.06 ** | 2.3 ± 0.07 ** | 5.8 ± 0.02 ** | 3.4 ± 0.23 ** | 2.6 ± 0.01 *** | |
| 4 | 8.8 ± 0.06 *** | 3.7 ± 0.03 * | 3.3 ± 0.05 * | 7.6 ± 0.10 * | 3.6 ± 0.16 * | 3.4 ± 0.08 * | |
| 5 | 16.0 ± 0.08 *** | 5.3 ± 0.04 *** | 4.8 ± 0.15 ** | 12.0 ± 0.06 * | 4.0 ± 0.07 ** | 5 ± 0.02 ** | |
| 6 | 24.0 ± 0.09 * | 6.4 ± 0.11 * | 5.6 ± 0.11 * | 18.0 ± 0.08 *** | 5.9 ± 0.18 ** | 5.7 ± 0.13 * | |
| SW | 1 | 3.1 ± 0.06 * | 1.1 ± 0.15 *** | 1.3 ± 0.16 ** | 2.9 ± 0.03 *** | 0.9 ± 0.12 ** | 1.1 ± 0.15 * |
| 2 | 3.8 ± 0.09 *** | 1.6 ± 0.20 * | 1.7 ± 0.03 *** | 3.6 ± 0.05 * | 1.3 ± 0.09 * | 1.9 ± 0.06 ** | |
| 3 | 5.4 ± 0.07 * | 2.6 ± 0.18 * | 2.6 ± 0.04 * | 5.1 ± 0.02 * | 2.4 ± 0.16 * | 2.6 ± 0.03 ** | |
| 4 | 7.3 ± 0.05 *** | 3.2 ± 0.07 *** | 3.7 ± 0.01 *** | 7.0 ± 0.12 *** | 3 ± 0.13 *** | 3.8 ± 0.11 *** | |
| 5 | 9.6 ± 0.10 * | 3.8 ± 0.06 * | 4.3 ± 0.20 * | 9.2 ± 0.09 * | 3.7 ± 0.02 ** | 4.2 ± 0.13 * | |
| 6 | 11.2 ± 0.09 ** | 5.3 ± 0.04 * | 5.6 ± 0.18 | 11.0 ± 0.08 ** | 5.6 ± 0.05 * | 5.7 ± 0.13 *** | |
| Composition | Sea water | Deep seawater | |
|---|---|---|---|
| mg/kg | mg/kg | ||
| Main element | K | - | 380 |
| Na | 8900 | 10780 | |
| Ca | 393 | 403 | |
| Mg | 1080 | 1320 | |
| SiO | 2.0 | 2.8 | |
| Cl− | 12000 | 19350 | |
| SO4−2 | 2648 | 898 | |
| F− | 1 | 1.3 | |
| Li | 0.16 | 0.18 | |
| Sr | 7.8 | 14.3 | |
| Trace element | Fe | 0.02 | 0.09 |
| Mn | 0.03 | 0.16 | |
| Cu | - | 0.26 | |
| Pb | - | 0.11 | |
| Zn | - | 0.45 | |
| Cd | - | 0.05 | |
| Ni | - | 0.36 | |
| Nutritive salts | NO3− | 0.04 | 0.28 |
| PO43− | 0.012 | 0.06 | |
| Si | 0.44 | 2.8 | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Choi, W.Y.; Kang, D.H.; Lee, H.Y. Enhancement of Immune Activation Activities of Spirulina maxima Grown in Deep-Sea Water. Int. J. Mol. Sci. 2013, 14, 12205-12221. https://doi.org/10.3390/ijms140612205
Choi WY, Kang DH, Lee HY. Enhancement of Immune Activation Activities of Spirulina maxima Grown in Deep-Sea Water. International Journal of Molecular Sciences. 2013; 14(6):12205-12221. https://doi.org/10.3390/ijms140612205
Chicago/Turabian StyleChoi, Woon Yong, Do Hyung Kang, and Hyeon Yong Lee. 2013. "Enhancement of Immune Activation Activities of Spirulina maxima Grown in Deep-Sea Water" International Journal of Molecular Sciences 14, no. 6: 12205-12221. https://doi.org/10.3390/ijms140612205
APA StyleChoi, W. Y., Kang, D. H., & Lee, H. Y. (2013). Enhancement of Immune Activation Activities of Spirulina maxima Grown in Deep-Sea Water. International Journal of Molecular Sciences, 14(6), 12205-12221. https://doi.org/10.3390/ijms140612205