The Association of IFI27 Expression and Fatigue Intensification during Localized Radiation Therapy: Implication of a Para-Inflammatory Bystander Response

Abstract
:
1. Introduction
2. Results
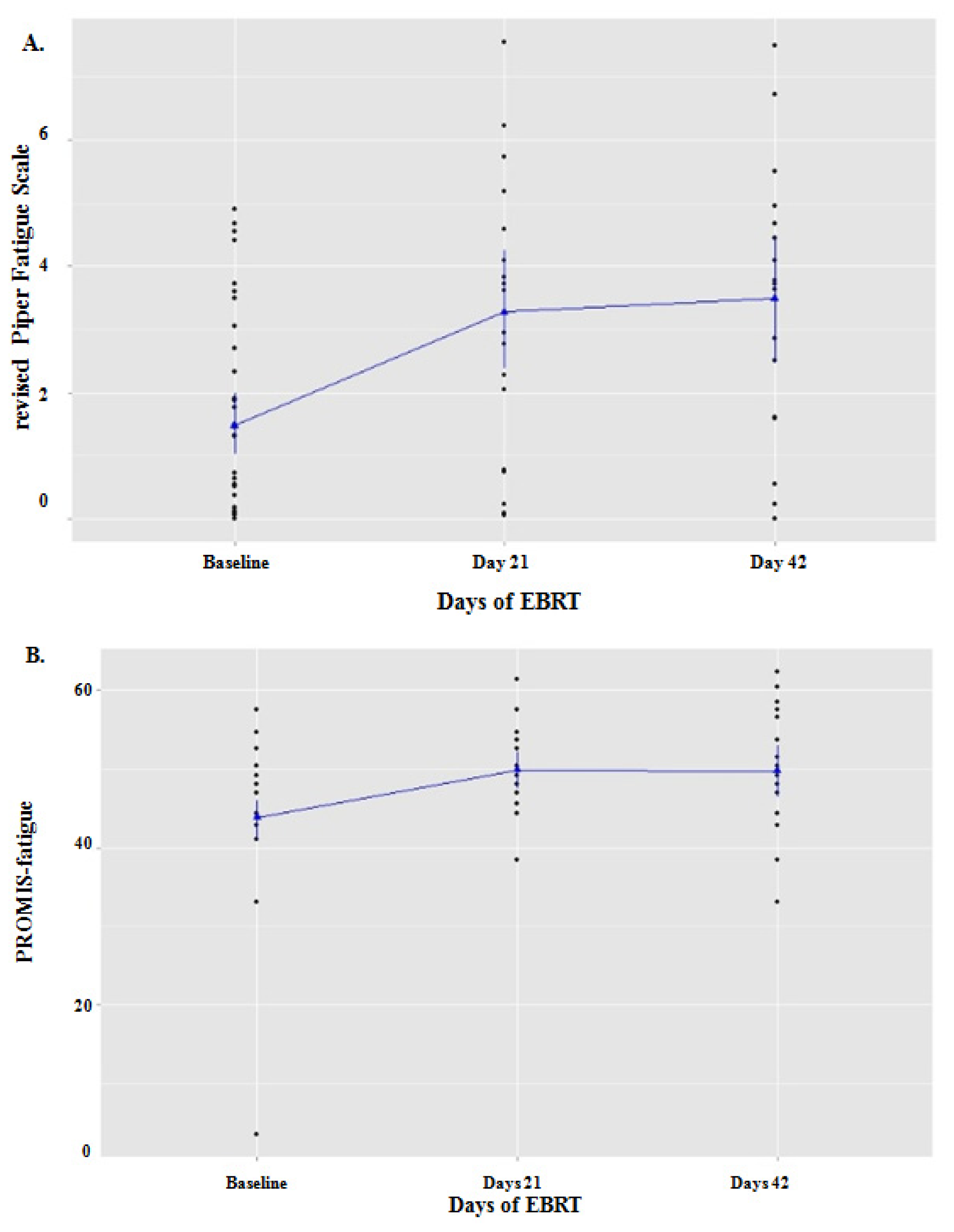
2.1. Fatigue during EBRT
2.2. Gene Expression by Microarray
2.3. Confirmation of IFI27 Expression during EBRT
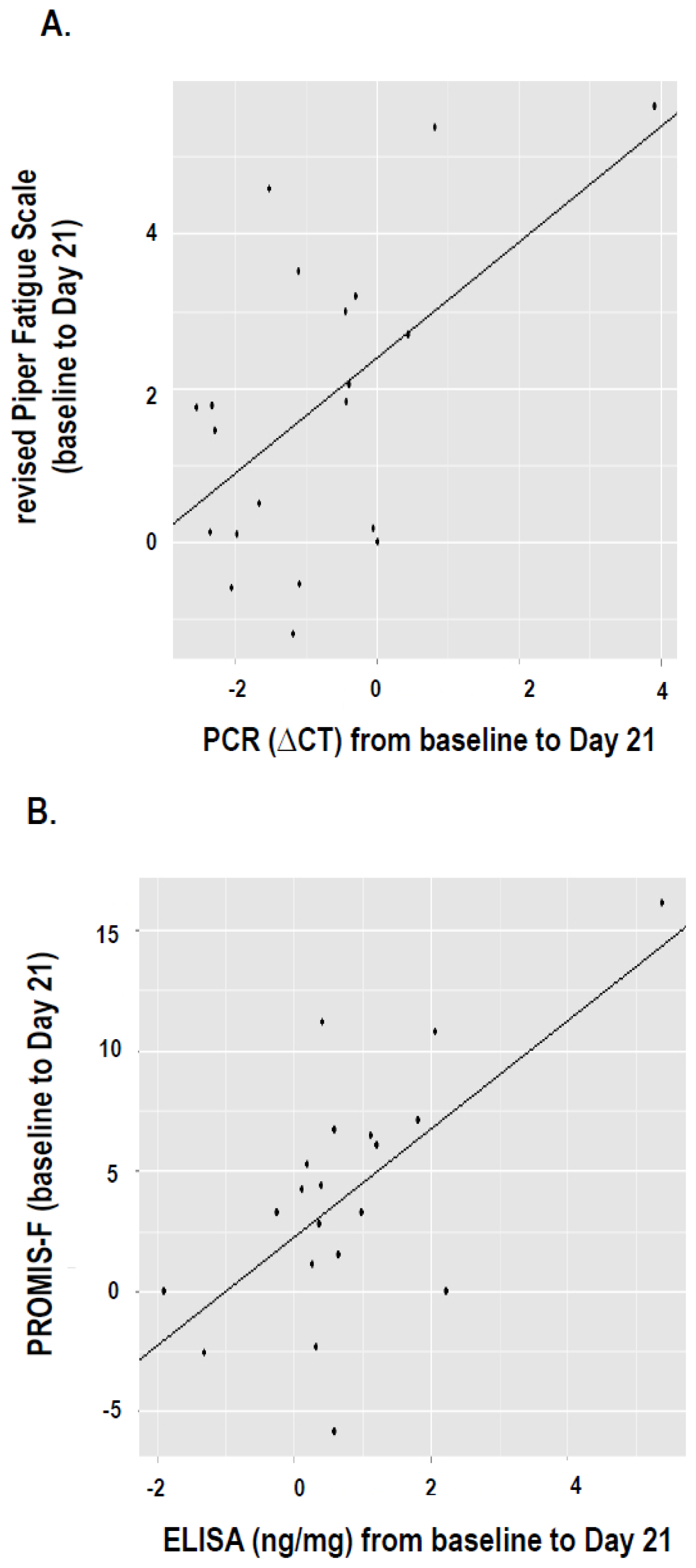
2.4. Correlation between Fatigue and IFI27 Expression
3. Discussion
4. Methods/Experimental Section
4.1. Gene Expression Chip Processing
4.2. Confirmatory Quantitative Real Time Polymerase Chain Reaction (qPCR)
4.3. Confirmation by Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Statistical Methods
5. Conclusions
Supplementary Information
ijms-14-16943-s001.pdfAcknowledgements
Conflicts of Interest
References
- Wong, W.W.; Vora, S.A.; Schild, S.E.; Ezzell, G.A.; Andrews, P.E.; Ferrigni, R.G.; Swanson, S.K. Radiation dose escalation for localized prostate cancer: Intensity-modulated radiotherapy versus permanent transperineal brachytherapy. Cancer 2009, 115, 5596–5606. [Google Scholar]
- Basivireddy, J.; Jacob, M.; Balasubramanian, K.A. Oral glutamine attenuates indomethacin-induced small intestinal damage. Clin. Sci. (Lond. ) 2004, 107, 281–289. [Google Scholar]
- Smets, E.M.; Visser, M.R.; Willems-Groot, A.F.; Garssen, B.; Oldenburger, F.; van Tienhoven, G.; de Haes, J.C. Fatigue and radiotherapy: A experience in patients undergoing treatment. Br. J. Cancer 1998, 78, 899–906. [Google Scholar]
- Miaskowski, C.; Paul, S.M.; Cooper, B.A.; Lee, K.; Dodd, M.; West, C.; Aouizerat, B.E.; Swift, P.S.; Wara, W. Trajectories of fatigue in men with prostate cancer before, during, and after radiation therapy. J. Pain Symptom Manage 2008, 35, 632–643. [Google Scholar]
- Saligan, L.N.; Kim, H.S. A systematic review of the association between immunogenomic markers and cancer-related fatigue. Brain. Behav. Immun 2012, 26, 830–848. [Google Scholar]
- Bower, J.E.; Ganz, P.A.; Tao, M.L.; Hu, W.; Belin, T.R.; Sepah, S.; Cole, S.; Aziz, N. Inflammatory biomarkers and fatigue during radiation therapy for breast and prostate cancer. Clin. Cancer Res 2009, 15, 5534–5540. [Google Scholar]
- Wratten, C.; Kilmurray, J.; Nash, S.; Seldon, M.; Hamilton, C.S.; O’Brien, P.C.; Denham, J.W. Fatigue during breast radiotherapy and its relationship to biological factors. Int. J. Radiat. Oncol. Biol. Phys 2004, 59, 160–167. [Google Scholar]
- Bower, J.E.; Ganz, P.A.; Irwin, M.R.; Kwan, L.; Breen, E.C.; Cole, S.W. Inflammation and behavioral symptoms after breast cancer treatment: do fatigue, depression, and sleep disturbance share a common underlying mechanism? J. Clin. Oncol 2011, 29, 3517–3522. [Google Scholar]
- Liu, L.; Mills, P.J.; Rissling, M.; Fiorentino, L.; Natarajan, L.; Dimsdale, J.E.; Sadler, G.R.; Parker, B.A.; Ancoli-Israel, S. Fatigue and sleep quality are associated with changes in inflammatory markers in breast cancer patients undergoing chemotherapy. Brain. Behav. Immun 2012, 26, 706–713. [Google Scholar]
- Aouizerat, B.E.; Dodd, M.; Lee, K.; West, C.; Paul, S.M.; Cooper, B.A.; Wara, W.; Swift, P.; Dunn, L.B.; Miaskowski, C. Preliminary Evidence of a Genetic Association Between Tumor Necrosis Factor Alpha and the Severity of Sleep Disturbance and Morning Fatigue. Biol. Res. Nurs 2009, 11, 27–41. [Google Scholar]
- Belloni, P.; Latini, P.; Palitti, F. Radiation-induced bystander effect in healthy G0 human lymphocytes: Biological and clinical significance. Mutat. Res. Fundam. Mol. Mech. Mutagenes 2011, 713, 32–38. [Google Scholar]
- Mariotti, L.G.; Bertolotti, A.; Ranza, E.; Babini, G.; Ottolenghi, A. Investigation of the mechanisms underpinning IL-6 cytokine release in bystander responses: the roles of radiation dose, radiation quality and specific ROS/RNS scavengers. Int. J. Radiat. Biol 2012, 88, 751–762. [Google Scholar]
- York, J.M.; Blevins, N.A.; Meling, D.D.; Peterlin, M.B.; Gridley, D.S.; Cengel, K.A.; Freund, G.G. The biobehavioral and neuroimmune impact of low-dose ionizing radiation. Brain. Behav. Immun 2012, 26, 218–227. [Google Scholar]
- Khayyal, M.T.; El-Ghazaly, M.A.; El-Hazek, R.M.; Nada, A.S. The effects of celecoxib, a COX-2 selective inhibitor, on acute inflammation induced in irradiated rats. Inflammopharmacology 2009, 17, 255–266. [Google Scholar]
- Donovan, K.; Jacobsen, P.; Andrykowski, M.; Winters, E.; Balducci, L.; Malik, U.; Kenady, D.; McGrath, P. Course of fatigue in women receiving chemotherapy and/or radiotherapy for early stage breast cancer. J. Pain Symptom Manage 2004, 28, 373–380. [Google Scholar]
- Wright, E.G. Manifestations and mechanisms of non-targeted effects of ionizing radiation. Mutat. Res. Fundam. Mol. Mech. Mutagenes 2010, 687, 28–33. [Google Scholar]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar]
- Wood, L.J.; Nail, L.M.; Perrin, N.A.; Elsea, C.R.; Fischer, A.; Druker, B.J. The Cancer Chemotherapy Drug Etoposide (VP-16) Induces Proinflammatory Cytokine Production and Sickness Behavior–like Symptoms in a Mouse Model of Cancer Chemotherapy–Related Symptoms. Biol.Res. Nurs 2006, 8, 157–169. [Google Scholar]
- Hsiao, C.P.; Wang, D.; Kaushal, A.; Saligan, L. Mitochondria-related gene expression changes are associated with fatigue in patients with nonmetastatic prostate cancer receiving external beam radiation therapy. Cancer Nurs 2012, 36, 189–197. [Google Scholar]
- Saligan, L.N.; Hsiao, C.P.; Wang, D.; Wang, X.M.; John, L.S.; Kaushal, A.; Citrin, D.; Barb, J.J.; Munson, P.J.; Dionne, R.A. Upregulation of alpha-synuclein during localized radiation therapy signals the association of cancer-related fatigue with the activation of inflammatory and neuroprotective pathways. Brain. Behav. Immun 2012, 27, 63–70. [Google Scholar]
- Yost, K.J.; Eton, D.T.; Garcia, S.F.; Cella, D. Minimally important differences were estimated for six Patient-Reported Outcomes Measurement Information System-Cancer scales in advanced-stage cancer patients. J. Clin. Epidemiol 2011, 64, 507–516. [Google Scholar]
- Burnette, B.C.; Liang, H.; Lee, Y.; Chlewicki, L.; Khodarev, N.N.; Weichselbaum, R.R.; Fu, Y.X.; Auh, S.L. The efficacy of radiotherapy relies upon induction of type I interferon-dependent innate and adaptive immunity. Cancer Res 2011, 71, 2488–2496. [Google Scholar]
- Rosebeck, S.; Leaman, D.W. Mitochondrial localization and pro-apoptotic effects of the interferon-inducible protein ISG12a. Apoptosis 2008, 13, 562–572. [Google Scholar]
- Rasmussen, U.B.; Wolf, C.; Mattei, M.-G.; Chenard, M.-P.; Bellocq, J.-P.; Chambon, P.; Rio, M.-C.; Basset, P. Identification of a New Interferon-α-inducible Gene (p27) on Human Chromosome 14q32 and Its Expression in Breast Carcinoma. Cancer Res 1993, 53, 4096–4101. [Google Scholar]
- Kelly, J.M.; Porter, A.C.; Chernajovsky, Y.; Gilbert, C.S.; Stark, G.R.; Kerr, I.M. Characterization of a human gene inducible by alpha- and beta-interferons and its expression in mouse cells. EMBO J 1986, 5, 1601–1606. [Google Scholar]
- Belardelli, F.; Ferrantini, M. Cytokines as a link between innate and adaptive antitumor immunity. Trends Immunol 2002, 23, 201–208. [Google Scholar]
- Liu, Y.J.; Kanzler, H.; Soumelis, V.; Gilliet, M. Dendritic cell lineage, plasticity and cross-regulation. Nat. Immunol 2001, 2, 585–589. [Google Scholar]
- Naing, A.; Reuben, J.M.; Camacho, L.H.; Gao, H.; Lee, B.N.; Cohen, E.N.; Verschraegen, C.; Stephen, S.; Aaron, J.; Hong, D.; et al. Phase I Dose Escalation Study of Sodium Stibogluconate (SSG), a Protein Tyrosine Phosphatase Inhibitor, Combined with Interferon Alpha for Patients with Solid Tumors. J. Cancer 2011, 2, 81–89. [Google Scholar]
- Maher, K.J.; Klimas, N.G.; Fletcher, M.A. Chronic fatigue syndrome is associated with diminished intracellular perforin. Clin. Exp. Immunol 2005, 142, 505–511. [Google Scholar]
- Bower, J.E.; Ganz, P.A.; Aziz, N.; Fahey, J.L. Fatigue and proinflammatory cytokine activity in breast cancer survivors. Psychosom. Med 2002, 64, 604–611. [Google Scholar]
- Collado-Hidalgo, A.; Bower, J.E.; Ganz, P.A.; Cole, S.W.; Irwin, M.R. Inflammatory biomarkers for persistent fatigue in breast cancer survivors. Clin. Cancer Res 2006, 12, 2759–2766. [Google Scholar]
- Bower, J.E.; Ganz, P.A.; Aziz, N.; Olmstead, R.; Irwin, M.R.; Cole, S.W. Inflammatory responses to psychological stress in fatigued breast cancer survivors: relationship to glucocorticoids. Brain. Behav. Immun 2007, 21, 251–258. [Google Scholar]
- Eggermont, A.M.; Suciu, S.; Santinami, M.; Testori, A.; Kruit, W.H.; Marsden, J.; Punt, C.J.; Sales, F.; Gore, M.; Mackie, R.; et al. Adjuvant therapy with pegylated interferon alfa-2b versus observation alone in resected stage III melanoma: final results of EORTC 18991, a randomised phase III trial. Lancet 2008, 372, 117–126. [Google Scholar]
- Groenewegen, G.; Walraven, M.; Vermaat, J.; de Gast, B.; Witteveen, E.; Giles, R.; Haanen, J.; Voest, E. Targeting the endothelin axis with atrasentan, in combination with IFN-alpha, in metastatic renal cell carcinoma. Br. J. Cancer 2012, 106, 284–289. [Google Scholar]
- Felger, J.C.; Alagbe, O.; Pace, T.W.; Woolwine, B.J.; Hu, F.; Raison, C.L.; Miller, A.H. Early activation of p38 mitogen activated protein kinase is associated with interferon-alpha-induced depression and fatigue. Brain. Behav. Immun 2011, 25, 1094–1098. [Google Scholar]
- Fransson, P. Fatigue in prostate cancer patients treated with external beam radiotherapy: A prospective 5-year long-term patient-reported evaluation. J. Cancer res. Ther 2010, 6, 516–520. [Google Scholar]
- Piper, B.F.; Dibble, S.L.; Dodd, M.J.; Weiss, M.C.; Slaughter, R.E.; Paul, S.M. The revised Piper Fatigue Scale: psychometric evaluation in women with breast cancer. Oncol. Nurs. Forum 1998, 25, 677–684. [Google Scholar]
- Byar, K.L.; Berger, A.M.; Bakken, S.L.; Cetak, M.A. Impact of adjuvant breast cancer chemotherapy on fatigue, other symptoms, and quality of life. Oncol. Nurs. Forum 2006, 33, E18–E26. [Google Scholar]
- Hays, R.D.; Bjorner, J.B.; Revicki, D.A.; Spritzer, K.L.; Cella, D. Development of physical and mental health summary scores from the patient-reported outcomes measurement information system (PROMIS) global items. Qual. Life Res 2009, 18, 873–880. [Google Scholar]
- Dittner, A.J.; Wessely, S.C.; Brown, R.G. The assessment of fatigue: A practical guide for clinicians and researchers. J. Psychosom. Res 2004, 56, 157–170. [Google Scholar]
- Lydiatt, W.M.; Denman, D.; McNeilly, D.P.; Puumula, S.E.; Burke, W.J. A randomized, placebo-controlled trial of citalopram for the prevention of major depression during treatment for head and neck cancer. Arch. Otolaryngol. Head Neck Surg 2008, 134, 528–535. [Google Scholar]
- Wang, X.M.; Wu, T.X.; Hamza, M.; Ramsay, E.S.; Wahl, S.M.; Dionne, R.A. Rofecoxib modulates multiple gene expression pathways in a clinical model of acute inflammatory pain. Pain 2007, 128, 136–147. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Participants (n = 40) | p value | |
|---|---|---|---|
| Patients (n = 20) Mean (±SD) or N (%) | Controls (n = 20) Mean (±SD) or N (%) | ||
| Mean Age, years | 65.6 ± 7.5 | 62.8 ± 6.1 | 0.36 |
| T-stage | |||
| T1c | 7 (35%) | 15 (75%) | |
| T2a | 7 (35%) | 5 (25%) | |
| T2b- T2c | 3 (15%) | ||
| T3a- T3b | 3 (15%) | ||
| Gleason Score | |||
| 6–7 | 10 (50%) | 20 (100%) | |
| 8–10 | 10 (50%) | ||
| Karnofsky performance scale | 89.5 (±2.2) | 95.2 (±1.2) | 0.91 |
| Testosterone (ng/dL) | 211.7 (±167.5) | ||
| Thyroid Stimulating Hormone (μIU/mL) | 2.01 (±1.1) | ||
| PSA (ng/mL) | 15.8 (±13.2) | 2.68 (±1.9) | 0.02 |
| Albumin (g/dL) | 3.9 (±0.3) | ||
| Hemoglobin (mg/dL) | 13.7 (±0.9) | ||
| Hematocrit (%) | 40.3 (±3.8) | 42.1 (±1.7) | 0.22 |
| Depression (HAM-D) | 1.2 (±2.1) | 0.5 (±0.6) | 0.11 |
| Fatigue score (rPFS) | 1.51 (±1.4) | 1.46 (±1.7) | 0.93 |
| Total EBRT dosage (Gray) | |||
| 75.6 | 18 (90%) | ||
| 68.4 | 2 (10%) | ||
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hsiao, C.-P.; Araneta, M.; Wang, X.M.; Saligan, L.N. The Association of IFI27 Expression and Fatigue Intensification during Localized Radiation Therapy: Implication of a Para-Inflammatory Bystander Response. Int. J. Mol. Sci. 2013, 14, 16943-16957. https://doi.org/10.3390/ijms140816943
Hsiao C-P, Araneta M, Wang XM, Saligan LN. The Association of IFI27 Expression and Fatigue Intensification during Localized Radiation Therapy: Implication of a Para-Inflammatory Bystander Response. International Journal of Molecular Sciences. 2013; 14(8):16943-16957. https://doi.org/10.3390/ijms140816943
Chicago/Turabian StyleHsiao, Chao-Pin, Maria Araneta, Xiao Min Wang, and Leorey N. Saligan. 2013. "The Association of IFI27 Expression and Fatigue Intensification during Localized Radiation Therapy: Implication of a Para-Inflammatory Bystander Response" International Journal of Molecular Sciences 14, no. 8: 16943-16957. https://doi.org/10.3390/ijms140816943
APA StyleHsiao, C.-P., Araneta, M., Wang, X. M., & Saligan, L. N. (2013). The Association of IFI27 Expression and Fatigue Intensification during Localized Radiation Therapy: Implication of a Para-Inflammatory Bystander Response. International Journal of Molecular Sciences, 14(8), 16943-16957. https://doi.org/10.3390/ijms140816943