Effects of Reduced Prolamin on Seed Storage Protein Composition and the Nutritional Quality of Rice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
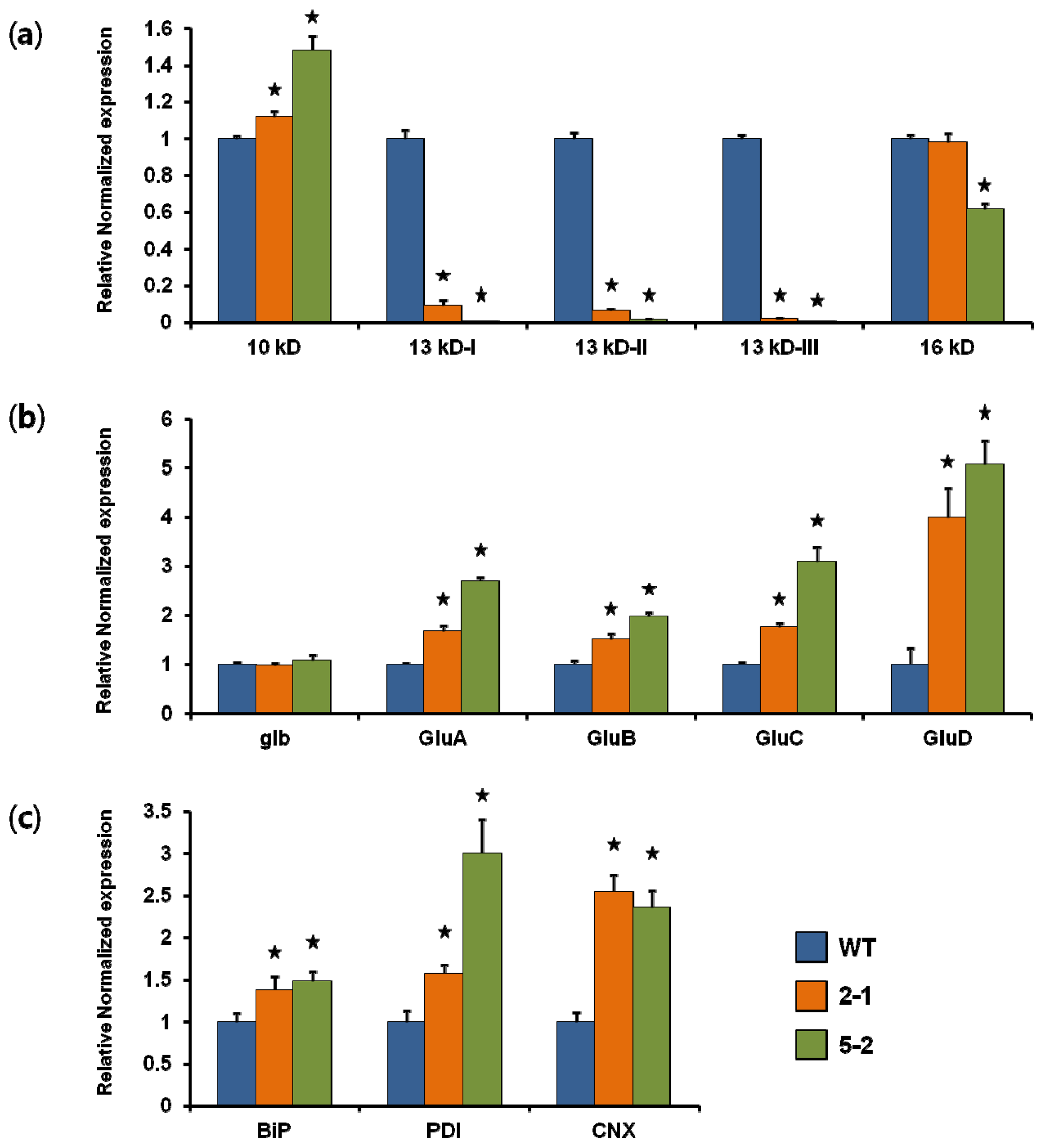
2.1. Generation of Transgenic Rice and Analysis of Transcript Levels
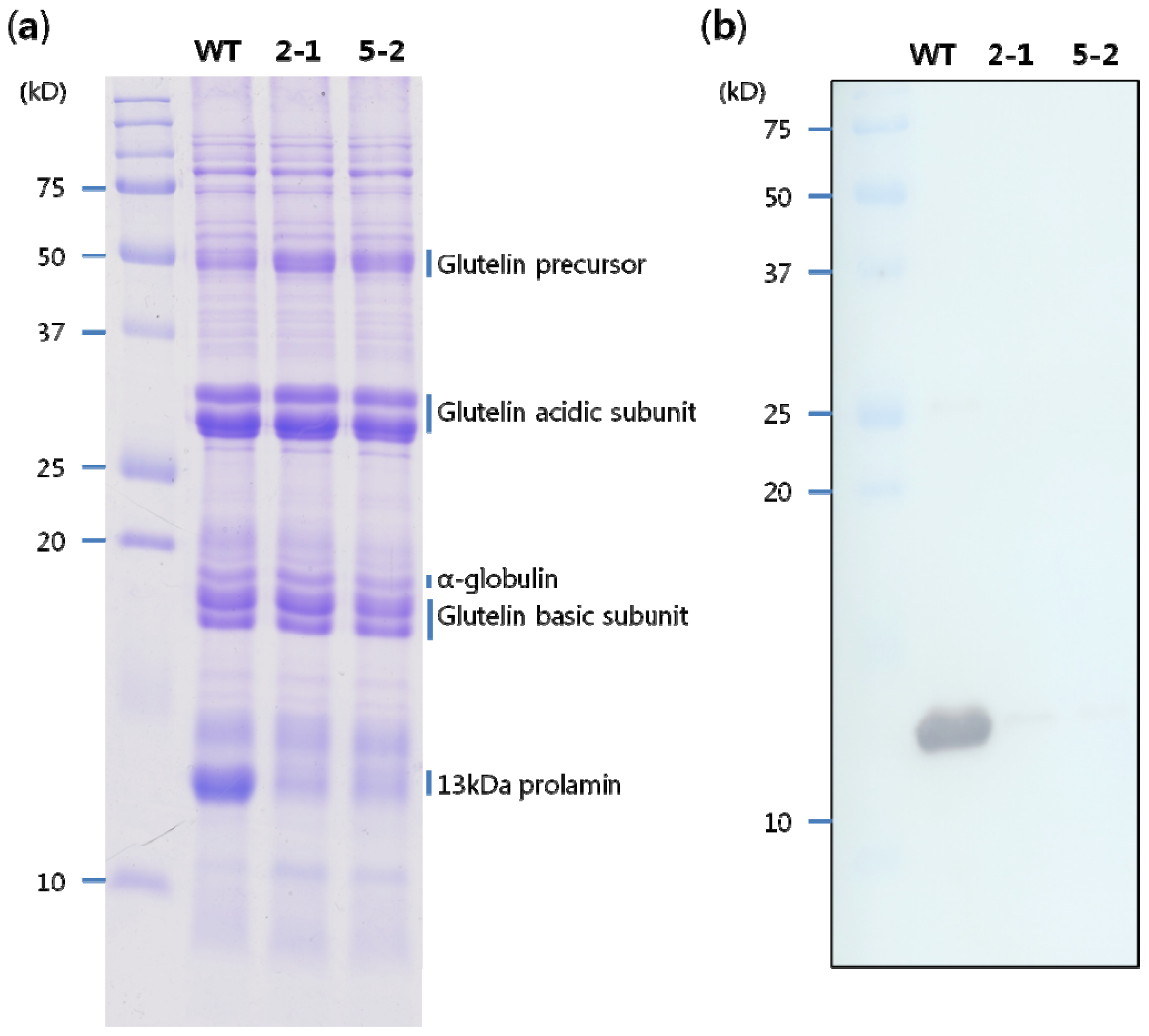
2.2. Storage Protein Profiles of Transgenic Rice Seeds
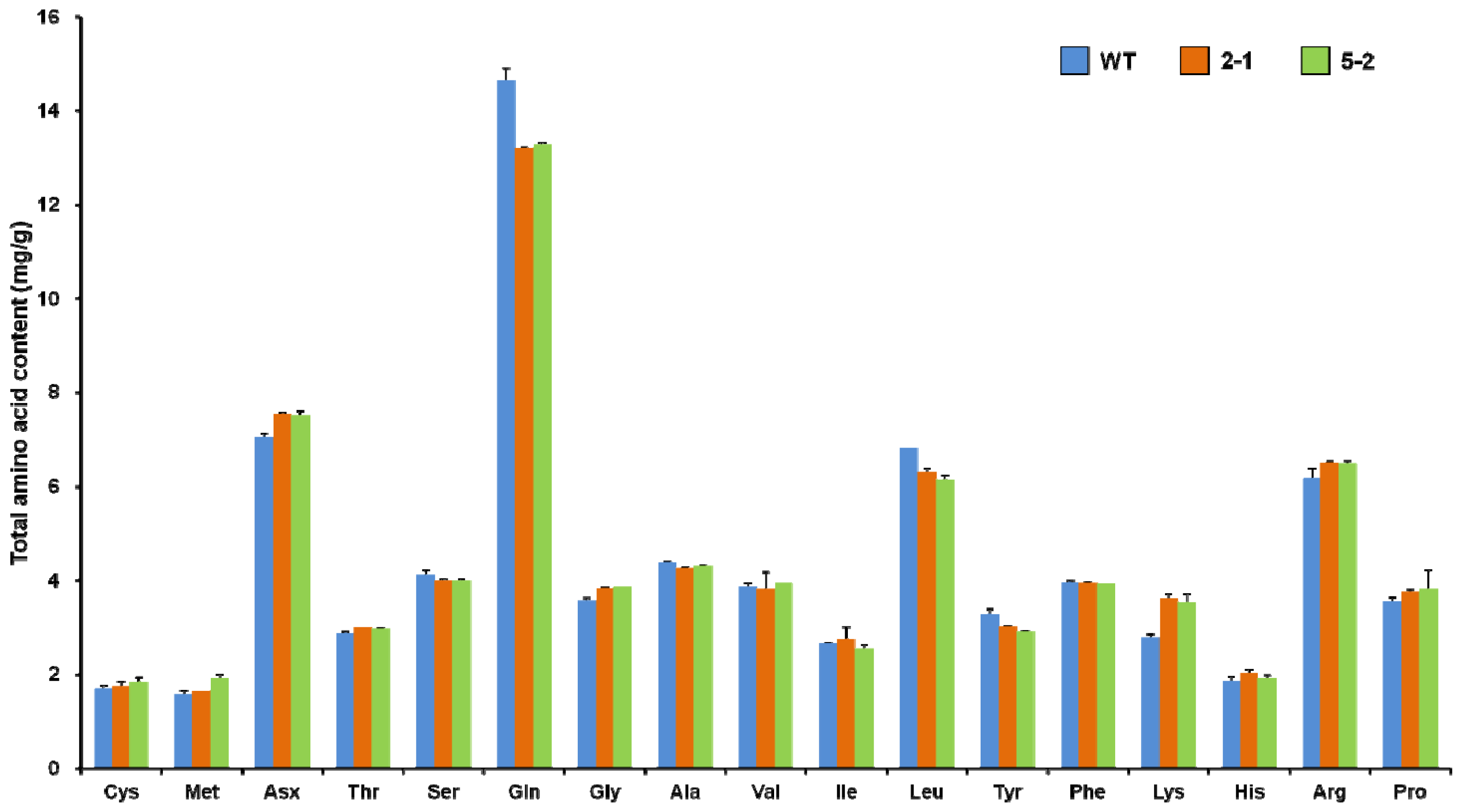
2.3. Total Amino Acid Profiles of Transgenic Rice Seeds
2.4. PB-I Formation in Transgenic Rice Seeds
3. Experimental Section
3.1. Plasmid Constructions
3.2. Rice Transformation
3.3. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
3.4. Seed Protein Extraction and Immunoblotting
3.5. Transmission Electron Microscopy (TEM)
3.6. Determination of Amino Acid Content
4. Conclusions
Supplementary Materials
ijms-14-17073-s001.pdfAcknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar]
- Bechtel, D.B.; Juliano, B.O. Formation of protein bodies in the starchy endosperm of rice (Oryza sativa L.): A re-investigation. Ann. Bot 1980, 45, 503–509. [Google Scholar]
- Krishnan, H.B.; Franceschi, V.R.; Okita, T.W. Immunochemical studies on the role of the golgi complex in protein-body formation in rice seeds. Planta 1986, 169, 471–480. [Google Scholar]
- Oparka, K.J.; Harris, N. Rice protein-body formation: All types are initiated by dilation of the endoplasmic reticulum. Planta 1982, 154, 184–188. [Google Scholar]
- Yamagata, H.; Tanaka, K. The site of synthesis and accumulation of rice storage proteins. Plant Cell Physiol 1986, 27, 135–145. [Google Scholar]
- Kawakatsu, T.; Yamamoto, M.P.; Hirose, S.; Yano, M.; Takaiwa, F. Characterization of a new rice glutelin gene GluD-1 expressed in the starchy endosperm. J. Exp. Bot 2008, 59, 4233–4245. [Google Scholar]
- Ogawa, M.; Kumamaru, T.; Satoh, H.; Iwata, N.; Omura, T.; Kasai, Z.; Tanaka, K. Purification of protein body-I of rice seed and its polypeptide composition. Plant Cell Physiol 1987, 28, 1517–1527. [Google Scholar]
- Li, X.; Okita, T.W. Accumulation of prolamines and glutelins during rice seed development: A quantitative-evaluation. Plant Cell Physiol 1993, 34, 385–390. [Google Scholar]
- Xu, J.H.; Messing, J. Amplification of prolamin storage protein genes in different subfamilies of the Poaceae. Theor. Appl. Genet 2009, 119, 1397–1412. [Google Scholar]
- Muench, D.G.; Ogawa, M.; Okita, T.W. The prolamins of rice. In Seed Proteins, 2nd ed.; Shewry, P.R., Casey, R., Eds.; Springer: Netherlands: Dordrecht, The Netherlands, 1999; pp. 93–108. [Google Scholar]
- Shorrosh, B.S.; Wen, L.; Zen, K.C.; Huang, J.K.; Pan, J.S.; Hermodson, M.A.; Tanaka, K.; Muthukrishnan, S.; Reeck, G.R. A novel cereal storage protein: Molecular genetics of the 19 kDa globulin of rice. Plant Mol. Biol 1992, 18, 151–154. [Google Scholar]
- Krishnan, H.B.; White, J.A.; Pueppke, S.G. Characterization and localization of rice (Oryza sativa L.) seed globulins. Plant Sci 1992, 81, 1–11. [Google Scholar]
- Tanaka, K.; Sugimoto, T.; Ogawa, M.; Kasai, Z. Isolation and characterization of two types of protein bodies in rice endosperm. Agric. Biol. Chem 1980, 44, 1633–1639. [Google Scholar]
- Li, X.; Wu, Y.; Zhang, D.Z.; Gillikin, J.W.; Boston, R.S.; Franceschi, V.R.; Okita, T.W. Rice prolamine protein body biogenesis: A BiP-mediated process. Science 1993, 262, 1054–1056. [Google Scholar]
- Maruta, Y.; Ueki, J.; Saito, H.; Nitta, N.; Imaseki, H. Transgenic rice with reduced glutelin content by transformation with glutelin a antisense gene. Mol. Breed 2001, 8, 273–284. [Google Scholar]
- Kusaba, M.; Miyahara, K.; Iida, S.; Fukuoka, H.; Takano, T.; Sassa, H.; Nishimura, M.; Nishio, T. Low glutelin content1: A dominant mutation that suppresses the glutelin multigene family via RNA silencing in rice. Plant Cell 2003, 15, 1455–1467. [Google Scholar]
- Kawakatsu, T.; Hirose, S.; Yasuda, H.; Takaiwa, F. Reducing rice seed storage protein accumulation leads to changes in nutrient quality and storage organelle formation. Plant Physiol 2010, 154, 1842–1854. [Google Scholar]
- Nagamine, A.; Matsusaka, H.; Ushijima, T.; Kawagoe, Y.; Ogawa, M.; Okita, T.W.; Kumamaru, T. A role for the cysteine-rich 10 kDa prolamin in protein body I formation in rice. Plant Cell Physiol 2011, 52, 1003–1016. [Google Scholar]
- Yoon, U.H.; Lee, J.H.; Hahn, J.H.; Kim, Y.K.; Lee, G.S.; Ji, H.S.; Kim, C.K.; Mun, J.H.; Kim, Y.M.; Kim, T.H. Structural and expression analysis of prolamin genes in Oryza sativa L. Plant Biotechnol. Rep 2012, 6, 251–262. [Google Scholar]
- Kim, Y.M.; Lee, J.Y.; Lee, T.; Lee, Y.H.; Kim, S.H.; Kang, S.H.; Yoon, U.H.; Ha, S.H.; Lim, S.H. The suppression of the glutelin storage protein gene in transgenic rice seeds results in a higher yield of recombinant protein. Plant Biotechnol. Rep 2012, 6, 347–353. [Google Scholar]
- Kim, W.T.; Okita, T.W. Structure, expression, and heterogeneity of the rice seed prolamines. Plant Physiol 1988, 88, 649–655. [Google Scholar]
- Masumura, T.; Hibino, T.; Kidzu, K.; Mitsukawa, N.; Tanaka, K.; Fujii, S. Cloning and characterization of a cDNA encoding a rice13 kDa prolamin. Mol. Gen. Genet 1990, 221, 1–7. [Google Scholar]
- Mitsukawa, N.; Konishi, R.; Uchiki, M.; Masumura, T.; Tanaka, K. Molecular cloning and characterization of a cysteine-rich 16.6-kDa prolamin in rice seeds. Biosci. Biotechnol. Biochem 1999, 63, 1851–1858. [Google Scholar]
- Muench, D.G.; Wu, Y.; Zhang, Y.; Li, X.; Boston, R.S.; Okita, T.W. Molecular cloning, expression and subcellular localization of a BiP homolog from rice endosperm tissue. Plant Cell Physiol 1997, 38, 404–412. [Google Scholar]
- Takemoto, Y.; Coughlan, S.J.; Okita, T.W.; Satoh, H.; Ogawa, M.; Kumamaru, T. The rice mutant esp2 greatly accumulates the glutelin precursor and deletes the protein disulfide isomerase. Plant Physiol 2002, 128, 1212–1222. [Google Scholar]
- Kawagoe, Y.; Suzuki, K.; Tasaki, M.; Yasuda, H.; Akagi, K.; Katoh, E.; Nishizawa, N.K.; Ogawa, M.; Takaiwa, F. The critical role of disulfide bond formation in protein sorting in the endosperm of rice. Plant Cell 2005, 17, 1141–1153. [Google Scholar]
- Vitale, A.; Boston, R.S. Endoplasmic reticulum quality control and the unfolded protein response: Insights from plants. Traffic 2008, 9, 1581–1588. [Google Scholar]
- Onda, Y.; Kumamaru, T.; Kawagoe, Y. ER membrane-localized oxidoreductase Ero1 is required for disulfide bond formation in the rice endosperm. Proc. Natl. Acad. Sci. USA 2009, 106, 14156–14161. [Google Scholar]
- Kubota, M.; Satto, Y.; Masumura, T.; Kumagai, T.; Watanabe, R.; Fujimura, S.; Kadowaki, M. Improvement in the vivo digestibility of rice protein by alkali extraction is due to structural changes in prolamin/protein body-I particle. Biosci. Biotechnol. Biochem 2010, 74, 614–619. [Google Scholar]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot 2002, 53, 947–958. [Google Scholar]
- Saito, Y.; Shigemitsu, T.; Yamasaki, R.; Sasou, A.; Goto, F.; Kishida, K.; Kuroda, M.; Tanaka, K.; Morita, S.; Satoh, S.; et al. Formation mechanism of the internal structure of type I protein bodies in rice endosperm: Relationship between the localization of prolamin species and the expression of individual genes. Plant J 2012, 70, 1043–1055. [Google Scholar]
- Karimi, M.; Inze, D.; Depicker, A. GATEWAY vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci 2002, 7, 193–195. [Google Scholar]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 1994, 6, 271–282. [Google Scholar]
- ImageJ, version 1.47; software for image processing and analysis in Java; National Institutes of Health: Bethesda, MD, USA, 2013.




© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, H.-J.; Lee, J.-Y.; Yoon, U.-H.; Lim, S.-H.; Kim, Y.-M. Effects of Reduced Prolamin on Seed Storage Protein Composition and the Nutritional Quality of Rice. Int. J. Mol. Sci. 2013, 14, 17073-17084. https://doi.org/10.3390/ijms140817073
Kim H-J, Lee J-Y, Yoon U-H, Lim S-H, Kim Y-M. Effects of Reduced Prolamin on Seed Storage Protein Composition and the Nutritional Quality of Rice. International Journal of Molecular Sciences. 2013; 14(8):17073-17084. https://doi.org/10.3390/ijms140817073
Chicago/Turabian StyleKim, Hyun-Jung, Jong-Yeol Lee, Ung-Han Yoon, Sun-Hyung Lim, and Young-Mi Kim. 2013. "Effects of Reduced Prolamin on Seed Storage Protein Composition and the Nutritional Quality of Rice" International Journal of Molecular Sciences 14, no. 8: 17073-17084. https://doi.org/10.3390/ijms140817073
APA StyleKim, H.-J., Lee, J.-Y., Yoon, U.-H., Lim, S.-H., & Kim, Y.-M. (2013). Effects of Reduced Prolamin on Seed Storage Protein Composition and the Nutritional Quality of Rice. International Journal of Molecular Sciences, 14(8), 17073-17084. https://doi.org/10.3390/ijms140817073