Icariin Protects Rat Cardiac H9c2 Cells from Apoptosis by Inhibiting Endoplasmic Reticulum Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
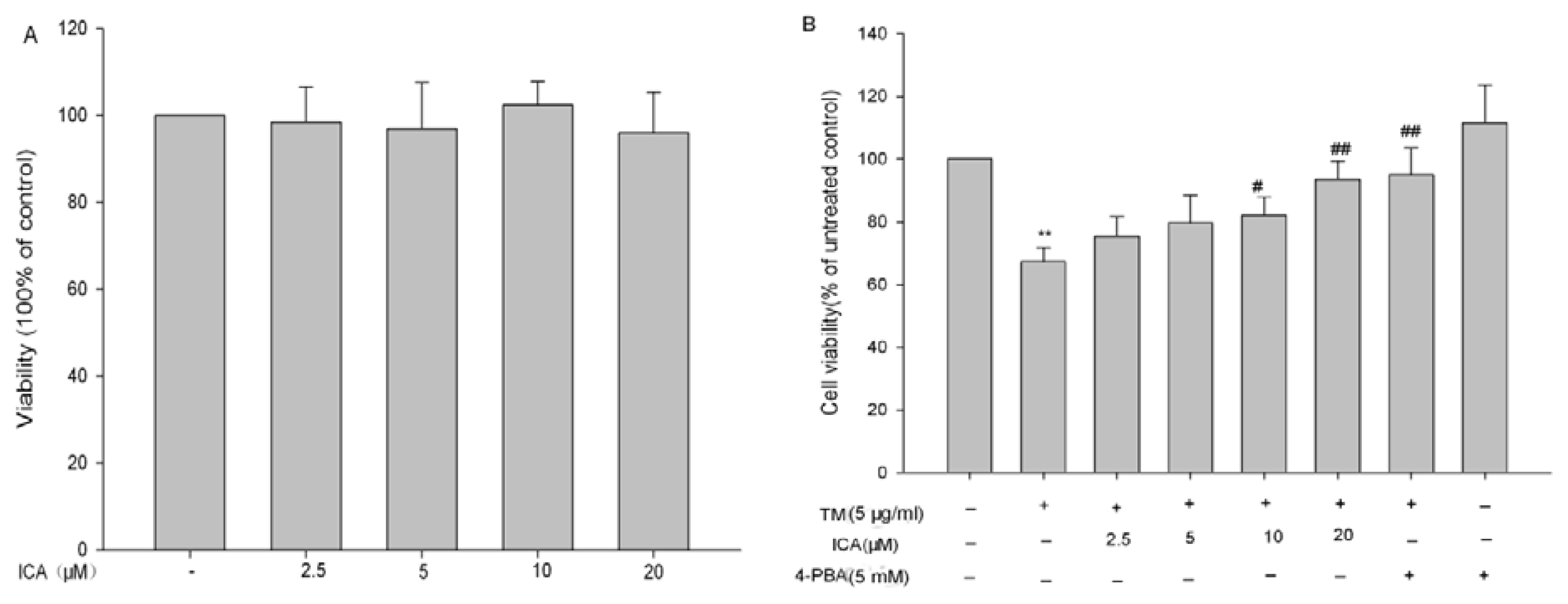
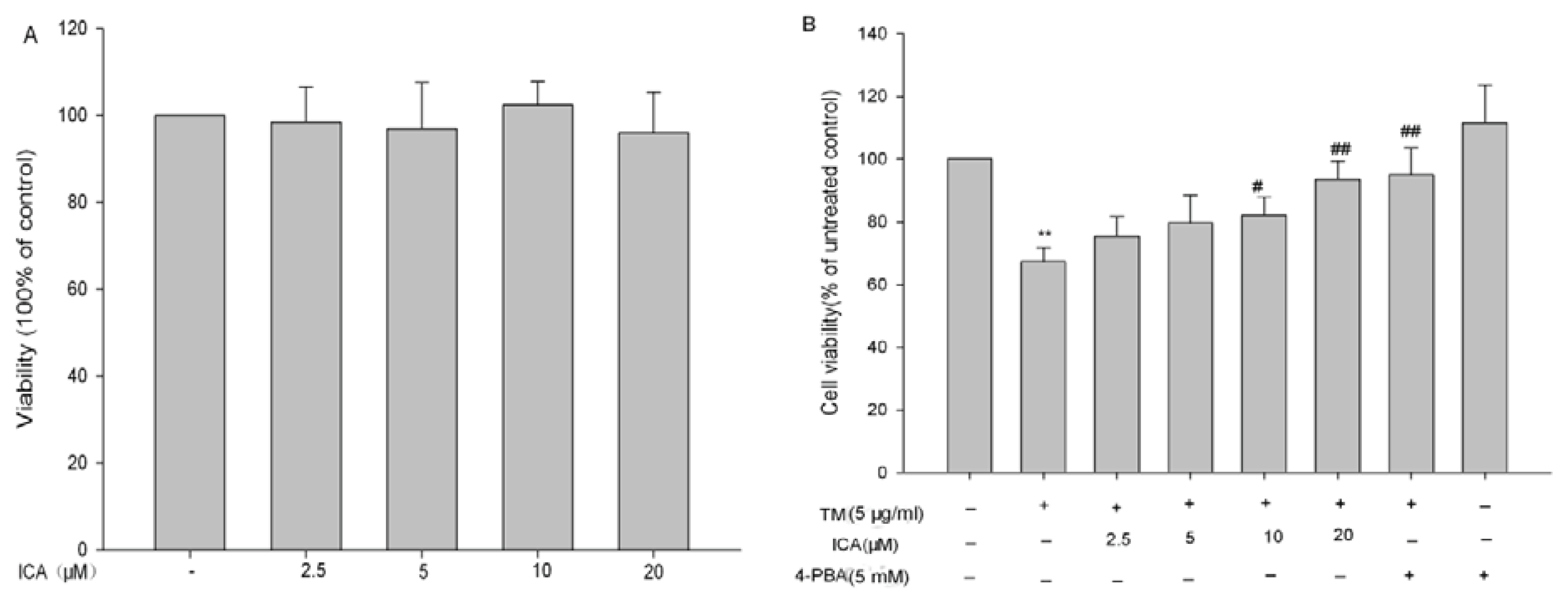
2.1. The Effects of Icariin on Cell Viability in H9c2 Cells
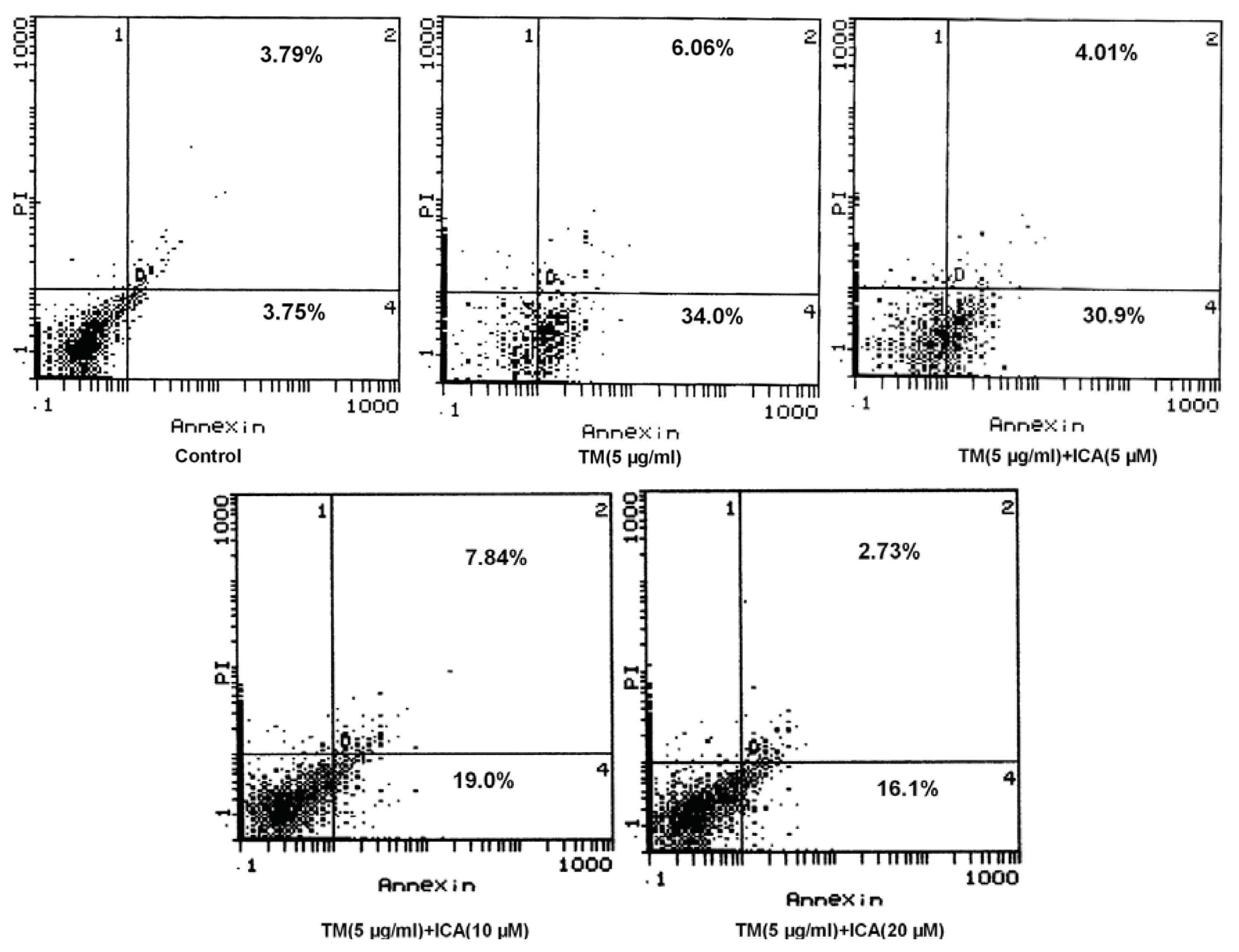
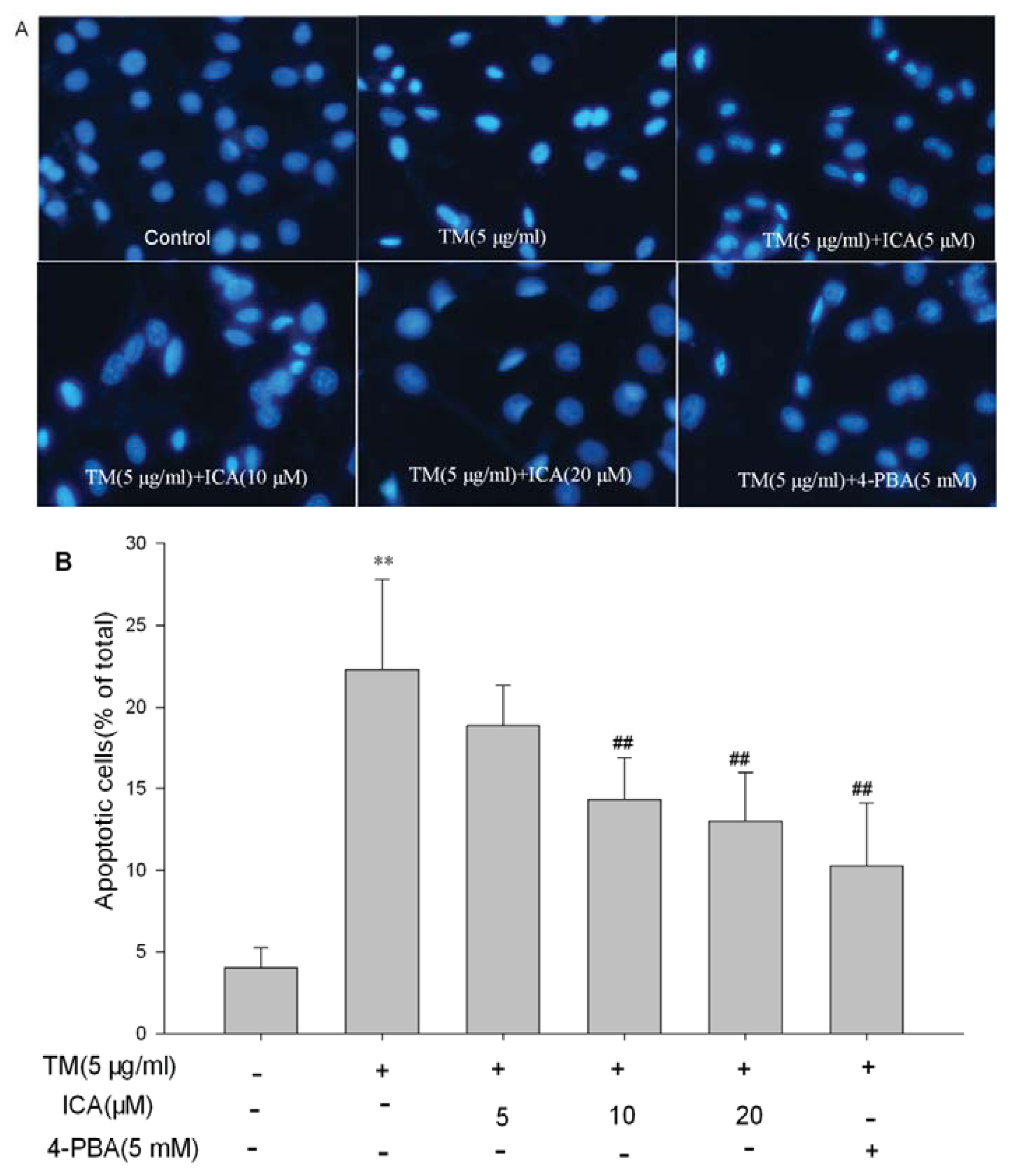
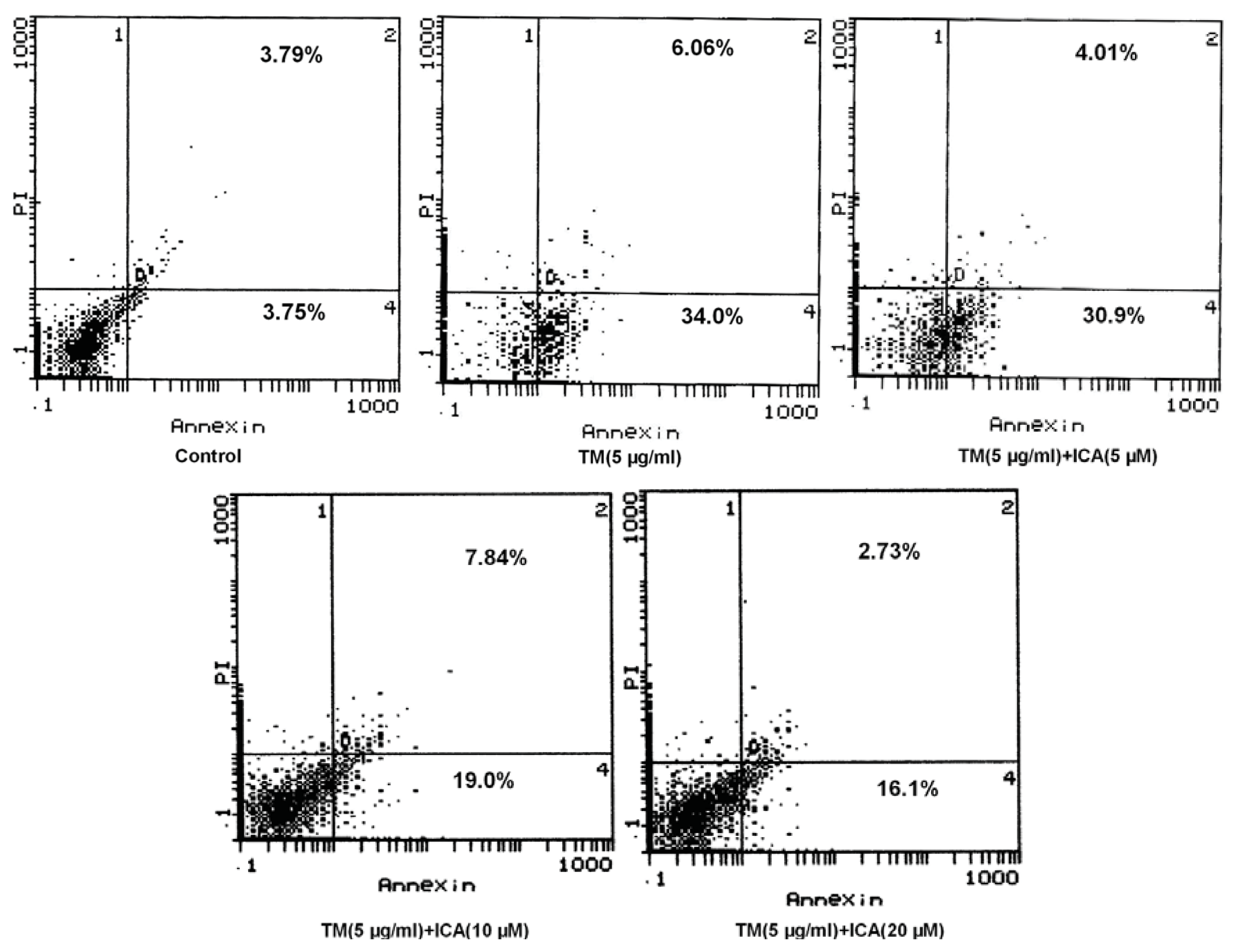
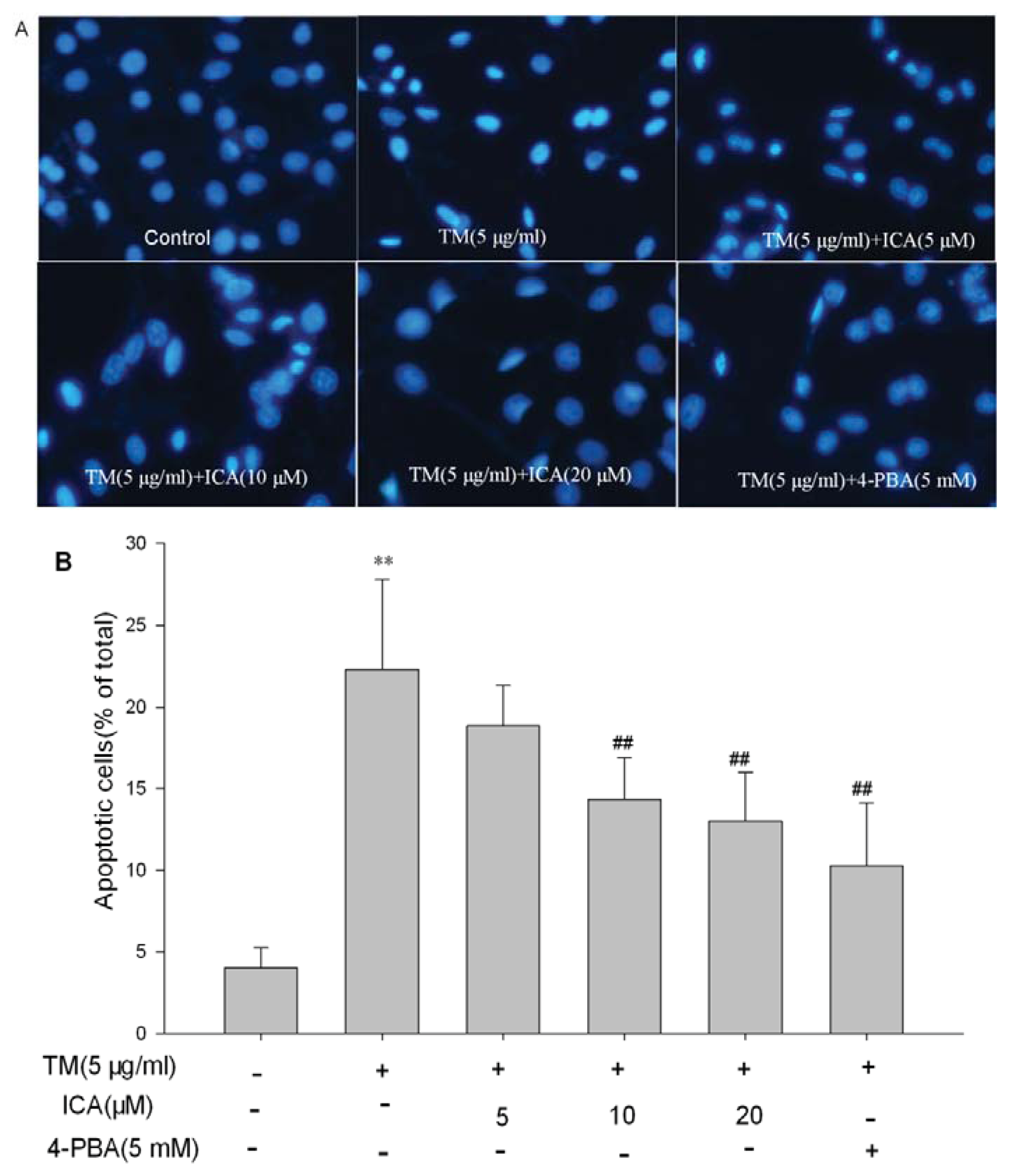
2.2. Icariin Reduces Apoptotic Cells Induced by Tunicamycin in H9c2 Cells
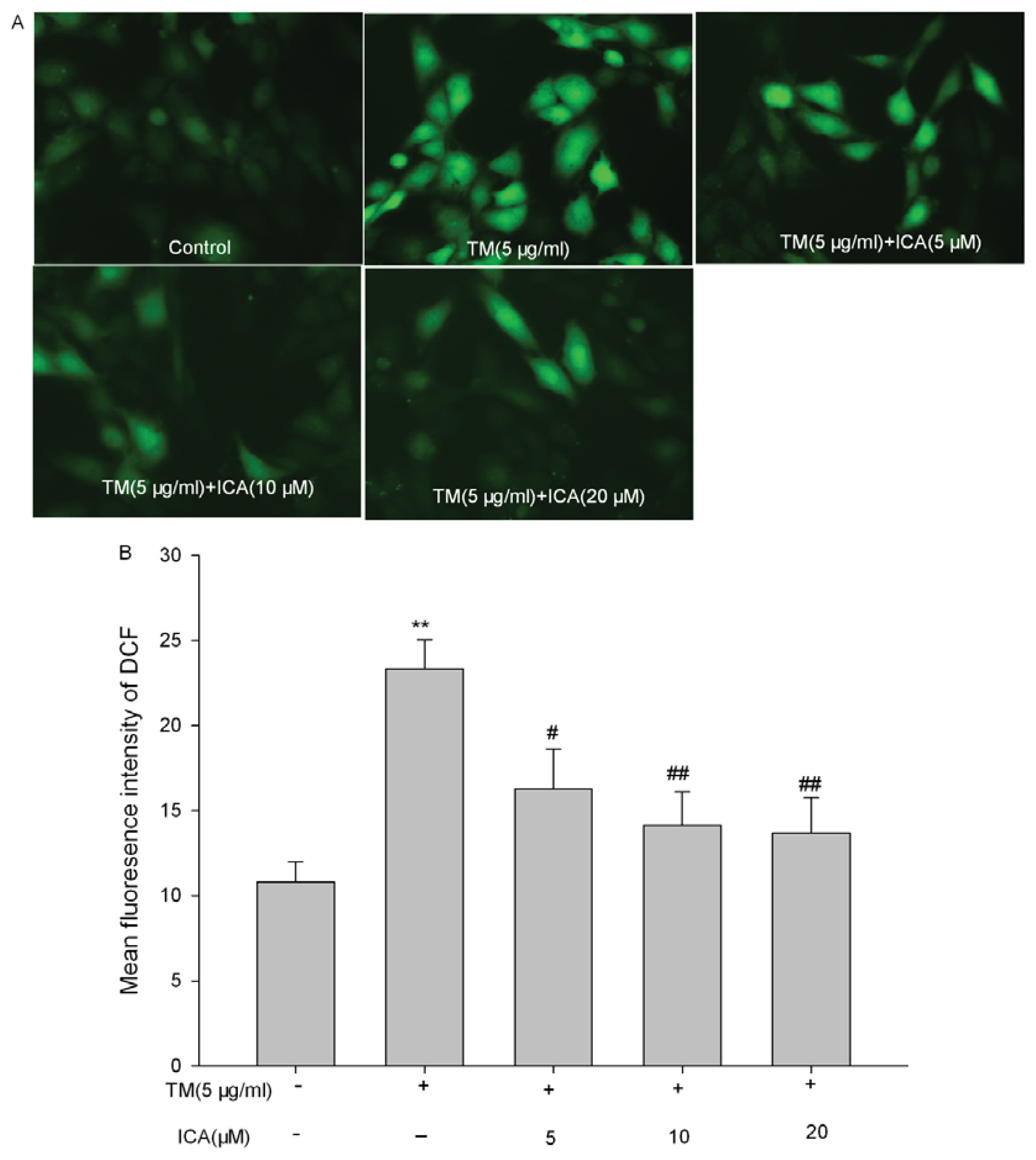
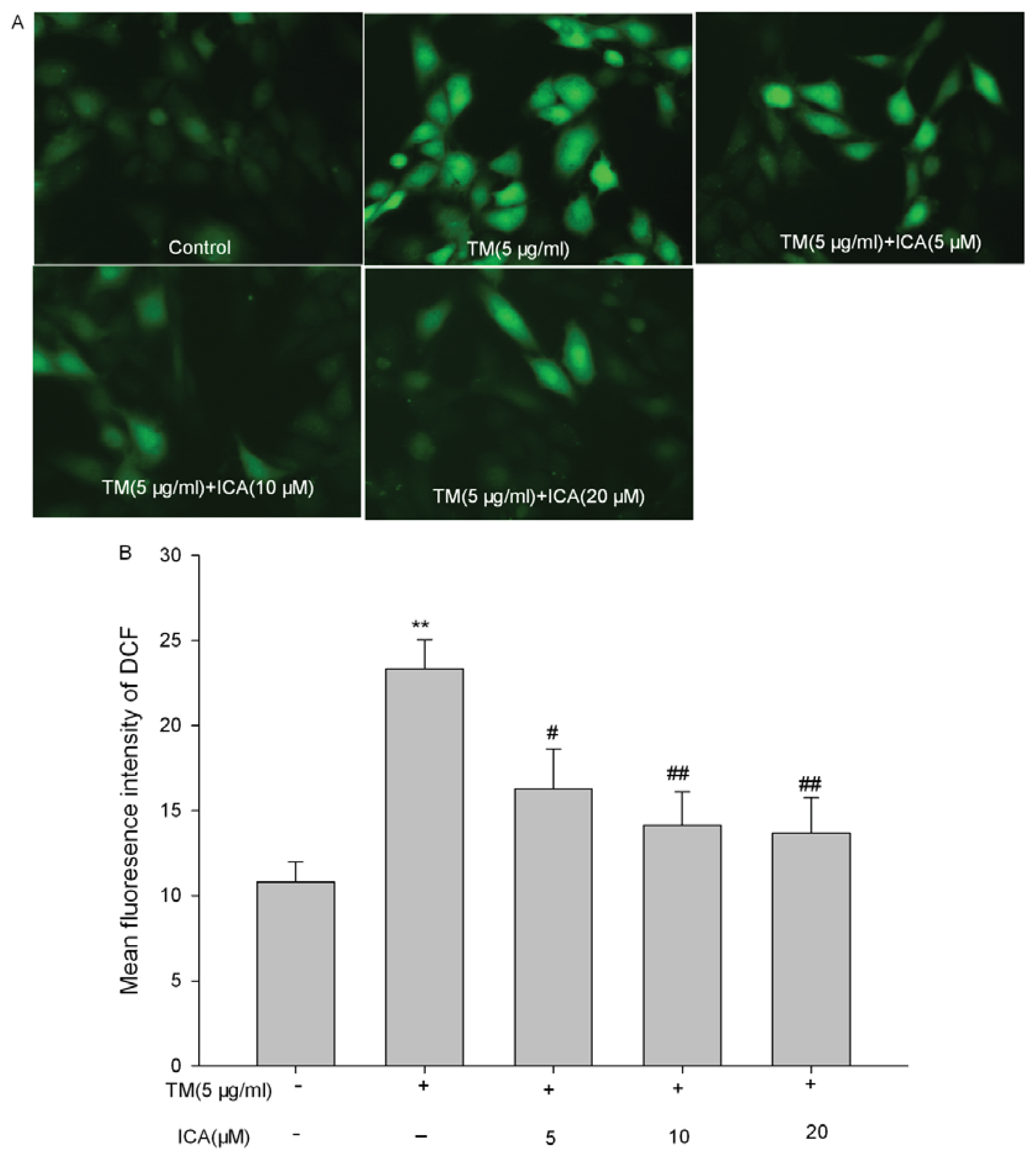
2.3. Effects of Icariin on the Intracellular ROS Level in Tunicamycin-Treated Cells
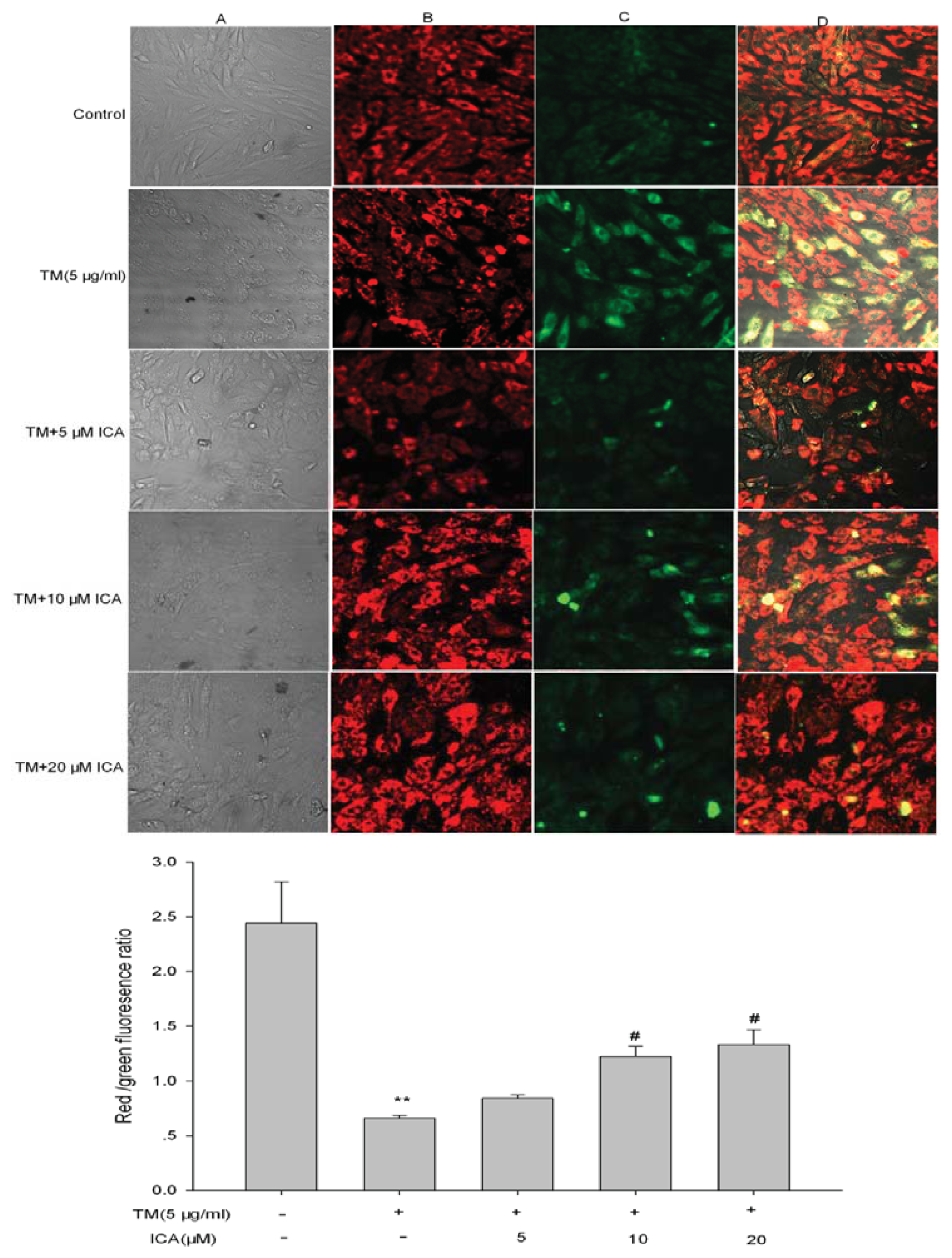
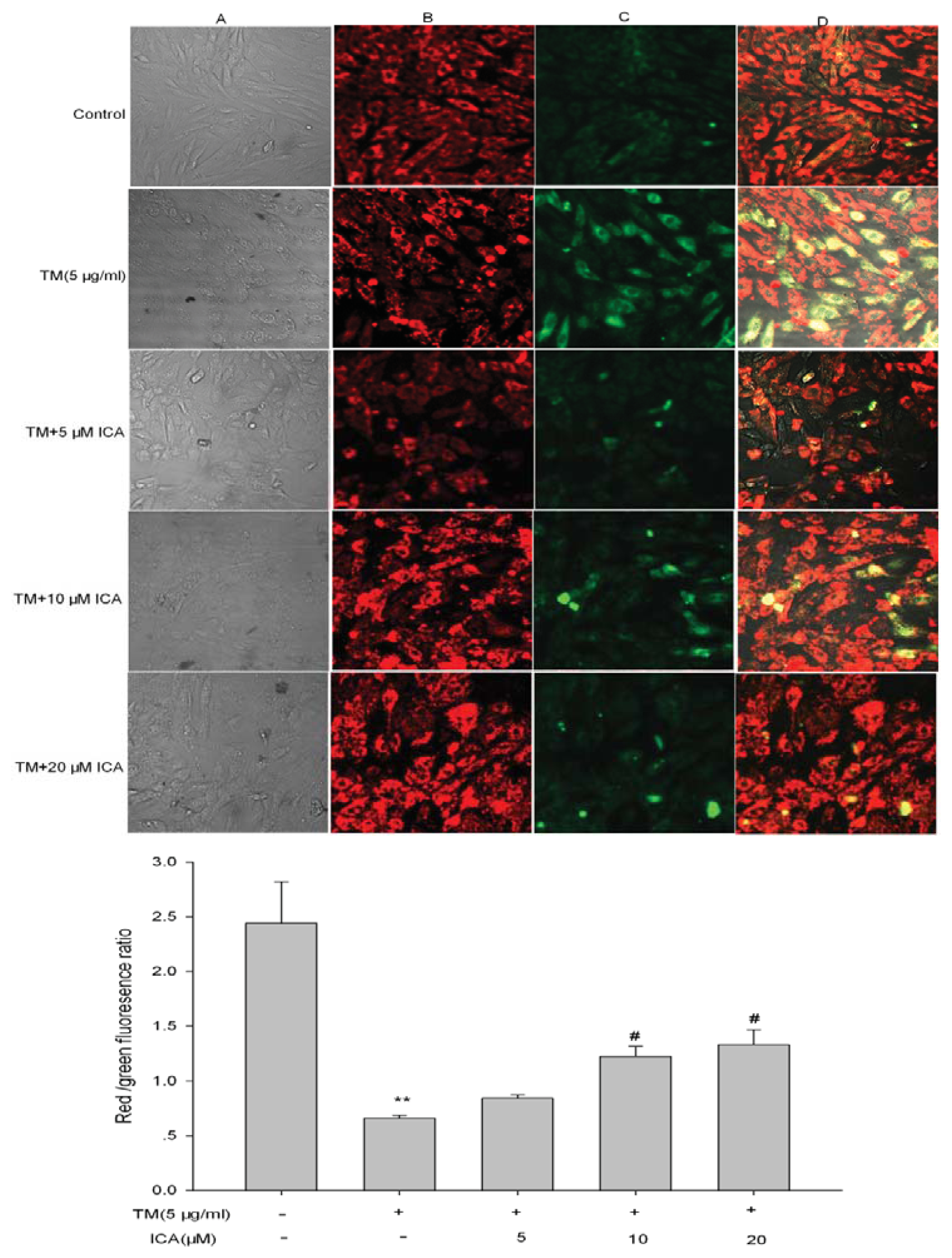
2.4. The Effect of Icariin on ΔΨm in H9c2 Cells
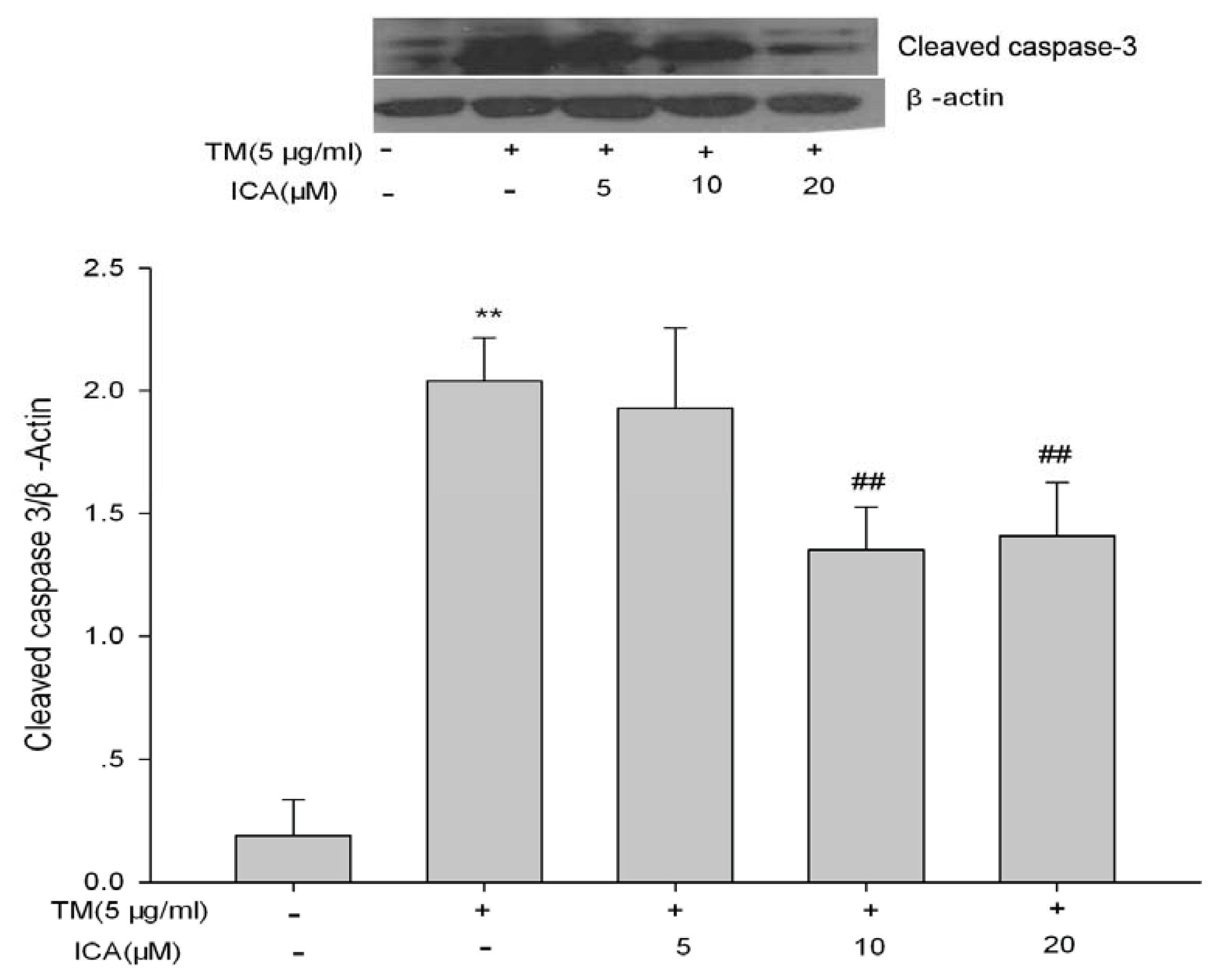
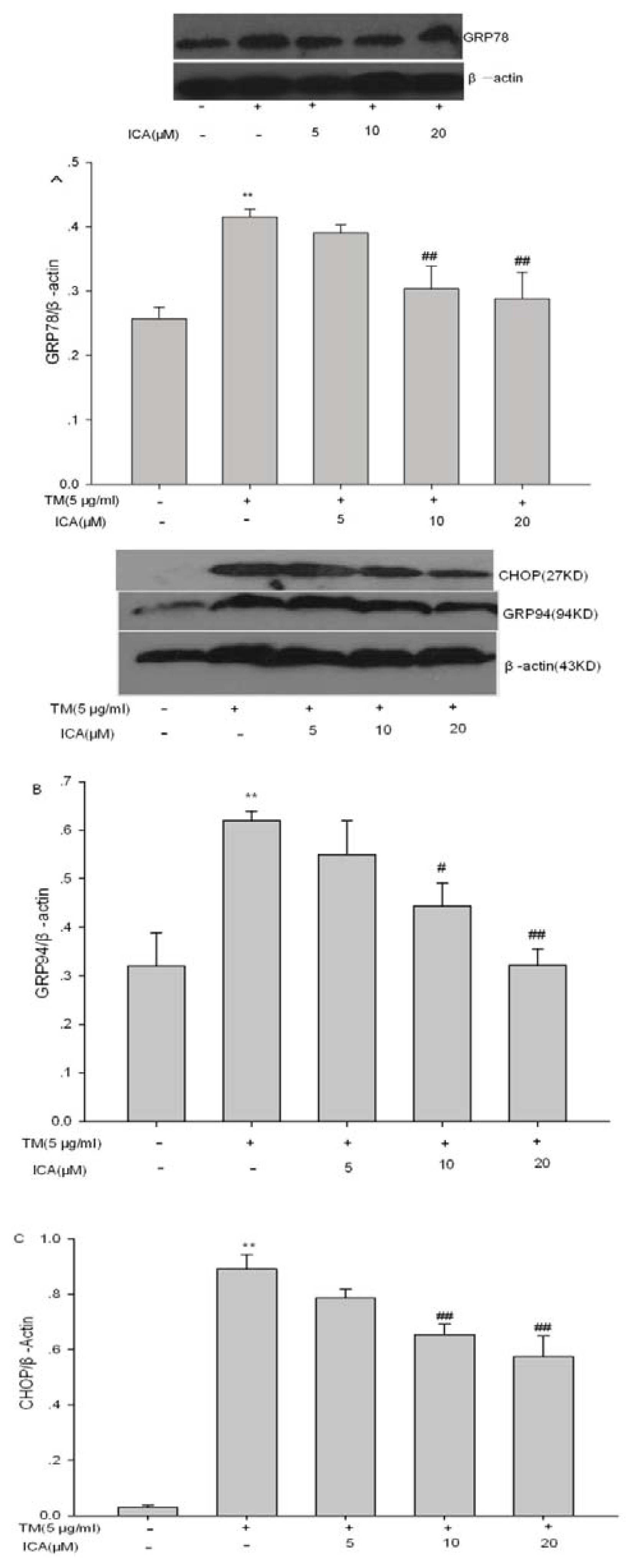
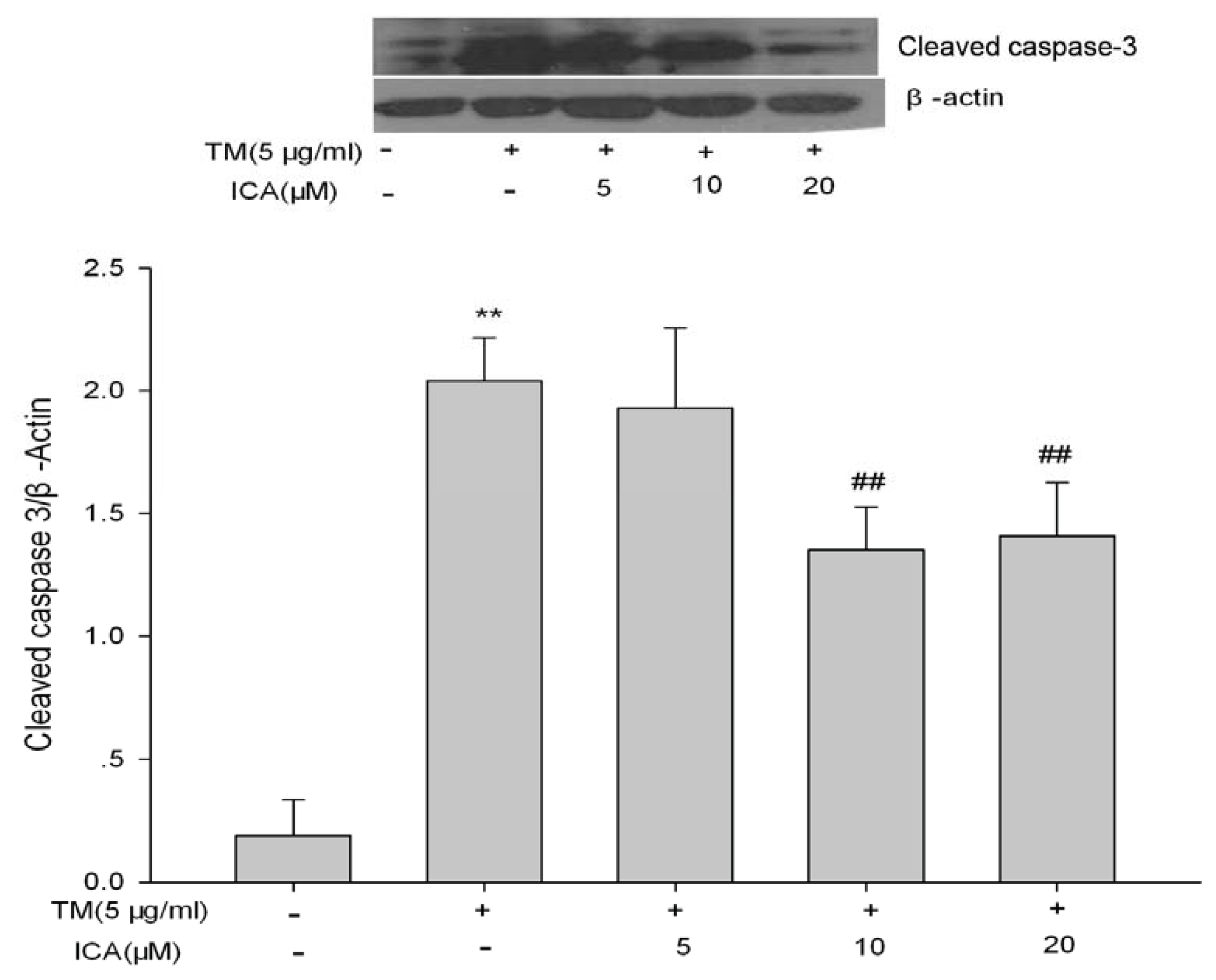
2.5. The Effects of Icariin on ERS Marker Proteins
3. Discussion
4. Experimental Section
4.1. Antibodies and Reagents
4.2. Cell Cultures
4.3. MTT Assay for Cell Viability
4.4. Hoechst 33258 Staining
4.5. Assessment of Apoptosis by Flow Cytometry
4.6. Assay of Intracellular Reactive Oxygen Species (ROS)
4.7. Measurement of Mitochondrial Membrane Potential (ΔΨm) Using JC-1
4.8. Western Blot Analysis of GRP78, GRP94, CHOP and Cleaved-Caspase 3
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Isodono, K.; Takahashi, T.; Imoto, H.; Nakanishi, N.; Ogata, T.; Asada, S.; Adachi, A.; Ueyama, T.; Oh, H.; Matsubara, H. PARM-1 is an endoplasmic reticulum molecule involved in endoplasmic reticulum stress-induced apoptosis in rat cardiac myocytes. PLoS One 2010, 5, e9746. [Google Scholar]
- Fu, H.Y.; Okada, K.; Liao, Y.; Tsukamoto, O.; Isomura, T.; Asai, M.; Sawada, T.; Okuda, K.; Asano, Y.; Sanada, S.; et al. Ablation of C/EBP homologous protein attenuates endoplasmic reticulum-mediated apoptosis and cardiac dysfunction induced by pressure overload. Circulation 2010, 122, 361–369. [Google Scholar]
- Wang, X.; Robbins, J. Heart failure and protein quality control. Circ. Res 2006, 99, 1315–1328. [Google Scholar]
- Minamino, T.; Komuro, I.; Kitakaze, M. Endoplasmic reticulum stress as a therapeutic target in cardiovascular disease. Circ. Res 2010, 107, 1071–1082. [Google Scholar]
- Kostin, S.; Pool, L.; Elsasser, A.; Hein, S.; Drexler, H.C.; Arnon, E.; Hayakawa, Y.; Zimmermann, R.; Bauer, E.; Klovekorn, W.P.; et al. Myocytes die by multiple mechanisms in failing human hearts. Circ. Res 2003, 92, 715–724. [Google Scholar]
- Anelli, T.; Sitia, R. Protein quality control in the early secretory pathway. EMBO J 2008, 27, 315–327. [Google Scholar]
- Kim, I.; Shu, C.W.; Xu, W.; Shiau, C.W.; Grant, D.; Vasile, S.; Cosford, N.D.; Reed, J.C. Chemical biology investigation of cell death pathways activated by endoplasmic reticulum stress reveals cytoprotective modulators of ASK1. J. Biol. Chem 2009, 284, 1593–1603. [Google Scholar]
- Han, S.W.; Li, C.; Ahn, K.O.; Lim, S.W.; Song, H.G.; Jang, Y.S.; Cho, Y.M.; Jang, Y.M.; Ghee, J.Y.; Kim, J.Y.; et al. Prolonged endoplasmic reticulum stress induces apoptotic cell death in an experimental model of chronic cyclosporine nephropathy. Am. J. Nephrol 2008, 28, 707–714. [Google Scholar]
- Shore, G.C.; Papa, F.R.; Oakes, S.A. Signaling cell death from the endoplasmic reticulum stress response. Curr. Opin. Cell Biol 2011, 23, 143–149. [Google Scholar]
- Okada, K.; Minamino, T.; Tsukamoto, Y.; Liao, Y.; Tsukamoto, O.; Takashima, S.; Hirata, A.; Fujita, M.; Nagamachi, Y.; Nakatani, T.; et al. Prolonged endoplasmic reticulum stress in hypertrophic and failing heart after aortic constriction: Possible contribution of endoplasmic reticulum stress to cardiac myocyte apoptosis. Circulation 2004, 110, 705–712. [Google Scholar]
- Leem, J.; Koh, E.H. Interaction between mitochondria and the endoplasmic reticulum: Implications for the pathogenesis of type 2 diabetes mellitus. Exp. Diabetes Res 2012, 2012, 242984, :1–242984:8.. [Google Scholar]
- Chen, K.M.; Ge, B.F.; Ma, H.P.; Liu, X.Y.; Bai, M.H.; Wang, Y. Icariin, a flavonoid from the herb Epimedium enhances the osteogenic differentiation of rat primary bone marrow stromal cells. Pharmazie 2005, 60, 939–942. [Google Scholar]
- Wang, L.; Zhang, L.; Chen, Z.B.; Wu, J.Y.; Zhang, X.; Xu, Y. Icariin enhances neuronal survival after oxygen and glucose deprivation by increasing SIRT1. Eur. J. Pharmacol 2009, 609, 40–44. [Google Scholar]
- Li, F.; Gong, Q.H.; Wu, Q.; Lu, Y.F.; Shi, J.S. Icariin isolated from Epimedium brevicornum Maxim attenuates learning and memory deficits induced by d-galactose in rats. Pharmacol. Biochem. Behav 2010, 96, 301–305. [Google Scholar]
- Pan, Y.; Kong, L.D.; Li, Y.C.; Xia, X.; Kung, H.F.; Jiang, F.X. Icariin from Epimedium brevicornum attenuates chronic mild stress-induced behavioral and neuroendocrinological alterations in male Wistar rats. Pharmacol. Biochem. Behav 2007, 87, 130–140. [Google Scholar]
- Pan, Y.; Wang, F.M.; Qiang, L.Q.; Zhang, D.M.; Kong, L.D. Icariin attenuates chronic mild stress-induced dysregulation of the LHPA stress circuit in rats. Psychoneuroendocrinology 2010, 35, 272–283. [Google Scholar]
- Urano, T.; Tohda, C. Icariin improves memory impairment in Alzheimer’s disease model mice (5xFAD) and attenuates amyloid beta-induced neurite atrophy. Phytother. Res 2010, 24, 1658–1663. [Google Scholar]
- Zeng, K.W.; Ko, H.; Yang, H.O.; Wang, X.M. Icariin attenuates beta-amyloid-induced neurotoxicity by inhibition of tau protein hyperphosphorylation in PC12 cells. Neuropharmacology 2010, 59, 542–550. [Google Scholar]
- Sun, X.; Jin, X.; Zhang, X.; Liu, C.; Lei, L.; Jin, L.; Liu, H. Icariin induces mouse embryonic stem cell differentiation into beating functional cardiomyocytes. Mol. Cell. Biochem 2011, 349, 117–123. [Google Scholar]
- Li, L.; Wu, Q.; Zhou, Q.X.; Shi, J.S. Protective effect of icariin against mitochondrial damage induced by oxygen free radical in rat cerebral cells. Chin. J. Pharmacol. Toxicol 2005, 19, 333–337. [Google Scholar]
- Song, Y.H.; Li, B.S.; Chen, X.M.; Cai, H. Ethanol extract from Epimedium brevicornum attenuates left ventricular dysfunction and cardiac remodeling through down-regulating matrix metalloproteinase-2 and-9 activity and myocardial apoptosis in rats with congestive heart failure. Int. J. Mol. Med 2008, 21, 117–124. [Google Scholar]
- Xu, J.; Wang, G.; Wang, Y.; Liu, Q.; Xu, W.; Tan, Y.; Cai, L. Diabetes- and angiotensin II-induced cardiac endoplasmic reticulum stress and cell death: Metallothionein protection. J. Cell. Mol. Med 2009, 13, 1499–1512. [Google Scholar]
- Dou, G.; Sreekumar, P.G.; Spee, C.; He, S.; Ryan, S.J.; Kannan, R.; Hinton, D.R. Deficiency of alphaB crystallin augments ER stress-induced apoptosis by enhancing mitochondrial dysfunction. Free Radic. Biol. Med 2012, 53, 1111–1122. [Google Scholar]
- Ono, Y.; Shimazawa, M.; Ishisaka, M.; Oyagi, A.; Tsuruma, K.; Hara, H. Imipramine protects mouse hippocampus against tunicamycin-induced cell death. Eur. J. Pharmacol 2012, 696, 83–88. [Google Scholar]
- Zhao, H.; Liao, Y.; Minamino, T.; Asano, Y.; Asakura, M.; Kim, J.; Asanuma, H.; Takashima, S.; Hori, M.; Kitakaze, M. Inhibition of cardiac remodeling by pravastatin is associated with amelioration of endoplasmic reticulum stress. Hypertens. Res 2008, 31, 1977–1987. [Google Scholar]
- Marciniak, S.J.; Yun, C.Y.; Oyadomari, S.; Novoa, I.; Zhang, Y.; Jungreis, R.; Nagata, K.; Harding, H.P.; Ron, D. CHOP induces death by promoting protein synthesis and oxidation in the stressed endoplasmic reticulum. Genes Dev 2004, 18, 3066–3077. [Google Scholar]
- Song, Y.H.; Cai, H.; Gu, N.; Qian, C.F.; Cao, S.P.; Zhao, Z.M. Icariin attenuates cardiac remodelling through down-regulating myocardial apoptosis and matrix metalloproteinase activity in rats with congestive heart failure. J. Pharm. Pharmacol 2011, 63, 541–549. [Google Scholar]
- Choi, S.E.; Lee, Y.J.; Jang, H.J.; Lee, K.W.; Kim, Y.S.; Jun, H.S.; Kang, S.S.; Chun, J.; Kang, Y. A chemical chaperone 4-PBA ameliorates palmitate-induced inhibition of glucose-stimulated insulin secretion (GSIS). Arch. Biochem. Biophys 2008, 475, 109–114. [Google Scholar]
- Malo, A.; Kruger, B.; Goke, B.; Kubisch, C.H. 4-phenylbutyric acid reduces endoplasmic reticulum stress, trypsin activation, and acinar cell apoptosis while increasing secretion in rat pancreatic acini. Pancreas 2012, 42, 92–101. [Google Scholar]
- Kim, H.R.; Lee, G.H.; Cho, E.Y.; Chae, S.W.; Ahn, T.; Chae, H.J. Bax inhibitor 1 regulates ER-stress-induced ROS accumulation through the regulation of cytochrome P450 2E1. J. Cell Sci 2009, 122, 1126–1133. [Google Scholar]
- Li, L.; Tsai, H.J.; Wang, X.M. Icariin inhibits the increased inward calcium currents induced by amyloid-beta(25–35) peptide in CA1 pyramidal neurons of neonatal rat hippocampal slice. Am. J. Chin. Med 2010, 38, 113–125. [Google Scholar]
- Li, L.; Zhou, Q.X.; Shi, J.S. Protective effects of icariin on neurons injured by cerebral ischemia/reperfusion. Chin. Med. J. (Engl. ) 2005, 118, 1637–1643. [Google Scholar]
- Wang, Y.K.; Huang, Z.Q. Protective effects of icariin on human umbilical vein endothelial cell injury induced by H2O2 in vitro. Pharmacol. Res 2005, 52, 174–182. [Google Scholar]
- Minamino, T.; Kitakaze, M. ER stress in cardiovascular disease. J. Mol. Cell. Cardiol 2010, 48, 1105–1110. [Google Scholar]
- Ayala, P.; Montenegro, J.; Vivar, R.; Letelier, A.; Urroz, P.A.; Copaja, M.; Pivet, D.; Humeres, C.; Troncoso, R.; Vicencio, J.M.; et al. Attenuation of endoplasmic reticulum stress using the chemical chaperone 4-phenylbutyric acid prevents cardiac fibrosis induced by isoproterenol. Exp. Mol. Pathol 2012, 92, 97–104. [Google Scholar]
- Yang, C.; Wang, Y.; Liu, H.; Li, N.; Sun, Y.; Zhonghui, L.; Ping, Y. Ghrelin rotects H9c2 ardiomyocytes from ngiotensin II-induced apoptosis through the endoplasmic reticulum stress pathway. J. Cardiovasc. Pharmacol 2012, 59, 465–471. [Google Scholar]
- Wang, X.Y.; Yang, C.T.; Zheng, D.D.; Mo, L.Q.; Lan, A.P.; Yang, Z.L.; Hu, F.; Chen, P.X.; Liao, X.X.; Feng, J.Q. Hydrogen sulfide protects H9c2 cells against doxorubicin-induced cardiotoxicity through inhibition of endoplasmic reticulum stress. Mol. Cell. Biochem 2012, 363, 419–426. [Google Scholar]
- Alexandratou, E.; Yova, D.; Loukas, S. A confocal microscopy study of the very early cellular response to oxidative stress induced by zinc phthalocyanine sensitization. Free Radic. Biol. Med 2005, 39, 1119–1127. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Q.; Li, H.; Wang, S.; Liu, M.; Feng, Y.; Wang, X. Icariin Protects Rat Cardiac H9c2 Cells from Apoptosis by Inhibiting Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2013, 14, 17845-17860. https://doi.org/10.3390/ijms140917845
Zhang Q, Li H, Wang S, Liu M, Feng Y, Wang X. Icariin Protects Rat Cardiac H9c2 Cells from Apoptosis by Inhibiting Endoplasmic Reticulum Stress. International Journal of Molecular Sciences. 2013; 14(9):17845-17860. https://doi.org/10.3390/ijms140917845
Chicago/Turabian StyleZhang, Qiufang, Hongliang Li, Shanshan Wang, Ming Liu, Yibin Feng, and Xuanbin Wang. 2013. "Icariin Protects Rat Cardiac H9c2 Cells from Apoptosis by Inhibiting Endoplasmic Reticulum Stress" International Journal of Molecular Sciences 14, no. 9: 17845-17860. https://doi.org/10.3390/ijms140917845
APA StyleZhang, Q., Li, H., Wang, S., Liu, M., Feng, Y., & Wang, X. (2013). Icariin Protects Rat Cardiac H9c2 Cells from Apoptosis by Inhibiting Endoplasmic Reticulum Stress. International Journal of Molecular Sciences, 14(9), 17845-17860. https://doi.org/10.3390/ijms140917845