1. Introduction
XPC is a 940 amino acid protein which harbors domains that can bind to damaged DNA and repair factors. In complex with RAD23B and CETN2 [
1], XPC recognizes DNA damage based on bulky disfigurations of DNA and recruits TFIIH to these sites, initiating the global genome nucleotide excision repair (GG-NER) pathway. XPC appears to not only recognize various structurally unrelated lesions, but also bind to undamaged DNA with substantial affinity. GG-NER is responsible for identifying and removing bulky adducts from the DNA typically caused by UV radiation. XPC, along with DDB2, has been established as a DNA damage recognition protein for GG-NER.
XPC is one of the seven complementation groups of xeroderma pigmentosum (XP). First described in 1933 with its seven complementation groups and their frequencies characterized in 1991 [
2], the molecular mechanism of xeroderma pigmentosum has become better understood though its genotype-phenotype relationship remains complex. Manifesting in children younger than one year old, XP symptoms include photosensitivity, neurological defects, and a predisposition to skin cancers. Patients in
XPC, the complementation group corresponding to mutations in the gene coding for the XPC protein, can exhibit XP or XP with neurological abnormalities. These patients have been observed with diminished DNA repair capacity, abnormal skin lesions, freckling, atrophy, telangiectasia, hypopigmentation, actinic keratosis, and multiple skin cancers including squamous cell carcinomas, basal cell carcinomas, and melanomas [
3], and unusual cases have been reported with neurological symptoms such as those appearing in systemic lupus erythematosus [
4], hyperactivity potentially linked to low levels of glycine, and autistic features [
5] and ophthalmological symptoms such as the clouding of the cornea, prominent vascular growth on the conjunctiva, and loss of lashes.
XPC is covalently bonded to different modifiers, including ubiquitin and its relative SUMO, during the course of its role in GG-NER [
6]. XPC ubiquitination is carried out by the DDB2-associated DDB1-CUL4-ROC1 complex, though the complete role of this modification has yet to be determined. Typically, ubiquitination is indicative of degradation by the ubiquitin-proteasome system; interestingly, the ubiquitination of XPC does not lead to its degradation, indicating an alternative role for XPC ubiquitination. Sugasawa
et al. indicated that ubiquitination of XPC by the DDB1-CUL4-ROC1 complex increased the affinity of XPC to damaged DNA and is potentially involved in the handoff of 6-4PP repair from DDB2 to XPC. The sumoylation of XPC has been proposed to protect XPC from degradation after UV irradiation. Recently, it has been indicated that XPC is ubiquitinated after sumoylation by RNF111, which serves to promote NER [
7]. How XPC is deubiquitinated and removed from sites of damage remains unexplored.
XPC functionally interacts with RAD23B, CETN2, TFIIH, and XPA in the context of NER. XPC functions in GG-NER within the XPC-RAD23B-CETN2 complex; the interaction of XPC and RAD23B has been shown to increase the affinity of XPC for damaged DNA [
8] while the interaction between XPC and CETN2 has been shown to stabilize XPC and promote NER [
1,
9]. The interactions of XPC with CETN2, RAD23B, and XPA have been biochemically characterized [
10,
11]. Interestingly, XPC can functionally interact with both RAD23B and RAD23A, a homolog of RAD23B [
12,
13]. XPC interacts with TFIIH to recruit the transcription factor to damaged DNA for the completion of NER [
14,
15]. XPC-RAD23B can also interact with XPA-RPA [
16] and HMG1 [
17] to recognize psoralen interstrand crosslinks (ICLs). Some of these interactors are chromatin remodeling factors. For example, XPC has been shown to interact with hSNF5, a component of the SWI/SNF ATP-dependent chromatin remodeling complex, in response to UV radiation [
18] and potentially interacts weakly with p150, a subunit of chromatin assembly factor 1 (CAF-1), though this interaction has yet to be confirmed [
19]. In base excision repair (BER), XPC interacts with thymine DNA glycosylase (TDG), an initiator of BER which responds to G/T mismatches formed from the deamination of 5-methylcytosines. XPC-RAD23B was shown to form a complex with TDG-bound DNA and stimulate TDG activity [
20]. XPC has also been shown to play roles outside damage repair. The XPC-RAD23B-CETN2 complex, shown to interact directly with Oct4 and Sox2, is requisite for stem cell self-renewal and efficient somatic cell reprogramming [
21]. Additionally, XPC has been identified in large screenings as interacting with other proteins in as of yet unknown capacities. These proteins include CHRAC1, MECP2, TOP2B, USP11, WRAP53, ZCCHC6 [
22], LSM3 [
23], SMAD1, ZNF512B [
24], and BANF1 [
25].
Though the majority of the XP symptoms can be explained by XPC’s role in the GG-NER pathway as a sensor of DNA damage, the causes of some symptoms, particularly those with neurological or ophthalmological effects, are unknown. Discovering the proteins that interact with XPC within the cell and, therefore, the cellular functions of XPC in addition to its role in GG-NER, could provide the understanding necessary to comprehend the full effects of xeroderma pigmentosum. In this study, we used a high-throughput Yeast Two Hybrid screening to elucidate the interactome of XPC. We identified 49 proteins that interact with XPC with roles in DNA repair and replication, proteolysis and post-translational modifications, transcription regulation, signal transduction, and metabolism. The diversity of these roles indicates that XPC is involved in many more cellular processes than previously thought and provides a gateway for further understanding of the effects of xeroderma pigmentosum.
2. Results and Discussion
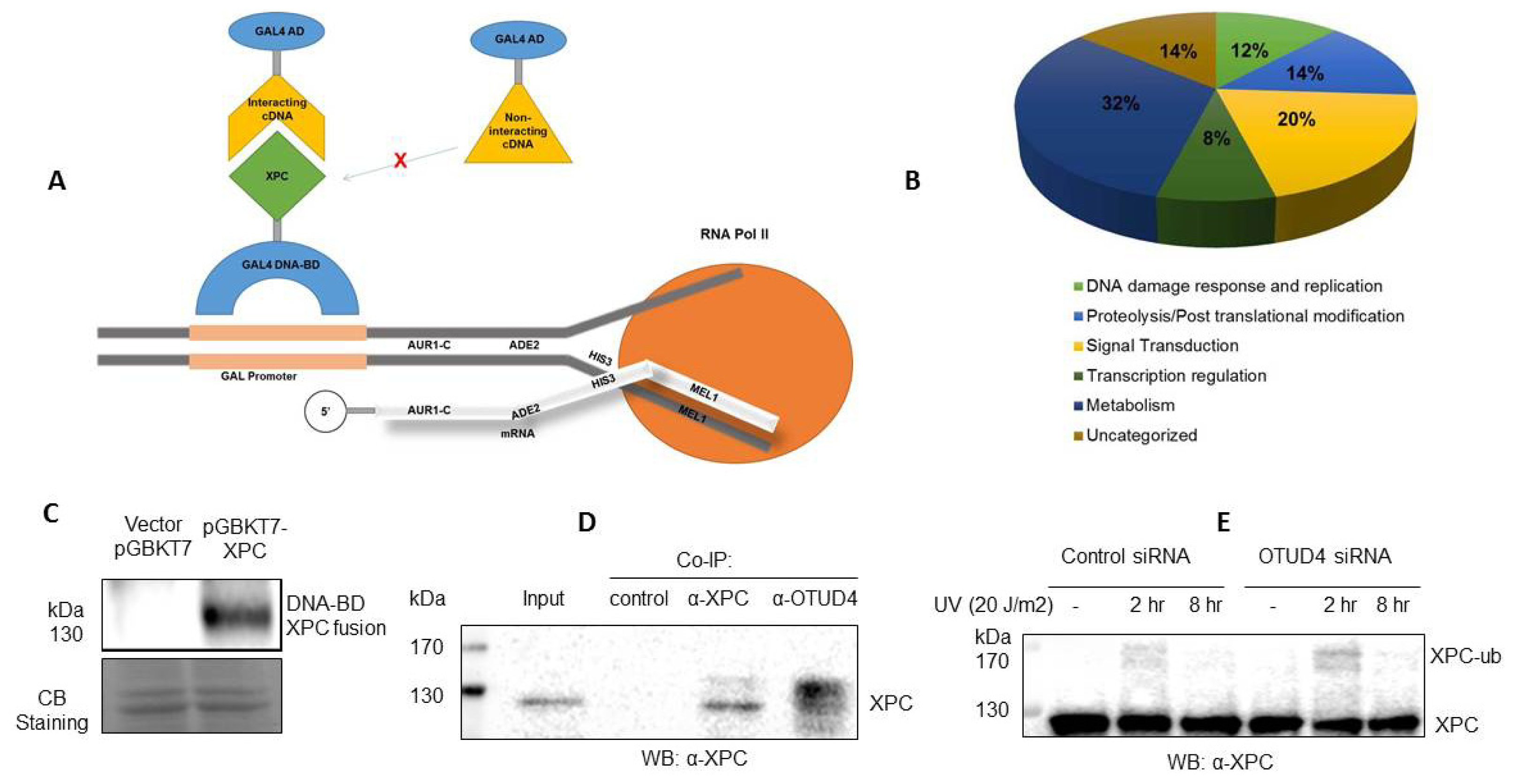
In this study, using an improved yeast two-hybrid system (
Figure 1A), we have identified 49 novel protein interactions with XPC. In order to further investigate the role of XPC within the cell, we have organized the functions into several categories: DNA repair and replication, proteolysis and post-translational modifications, transcription regulation, signal transduction, and metabolism (
Figure 1B). While XPC has been known to play a major role in DNA damage and be modified by ubiquitin and ubiquitin-like factors, the other pathways indicated in this screening could represent novel functions of XPC and explain symptoms of xeroderma pigmentosum with as of yet unknown etiology.
The yeast two-hybrid system we used is the Matchmaker Gold Yeast Two-Hybrid System, an advanced, high performing system for investigating protein–protein interactions. As the proximity of the DNA-BD and AD domains from the bait-prey interaction results in the transcription of four independent reporter genes (AUR1-C, ADE2, HIS3, and MEL1) of a new yeast strain (Y2H Gold), the incidence of false positives is very low. The rate of false positives was also reduced by the use of the SMART-based Normalized Yeast Two-Hybrid cDNA Library which removes highly abundant transcripts to allow for the screening of low abundance and rare cDNAs. However, the use of the Universal Human version of this library, while allowing for a broad gene representation, decreased the chances of finding a single, specific interaction.
We first confirmed that a specific XPC antibody recognizes the DNA-BD-XPC fusion protein used in our Yeast Two-Hybrid system (
Figure 1C). In addition, we have validated the OTUD4–XPC interaction in Human cells. We then focused on OTUD4, a novel deubiquitinase that we identified as an interactor with XPC. We found that XPC and OTUD4 interact directly in a co-immunoprecipitation experiment (
Figure 1D). The co-IP indicated a potential affinity of OTUD4 for modified XPC, as demonstrated by antibody binding specifically to bands higher than the 130 kDa XPC band. Significantly, a knockdown of
OTUD4 in the human HCT116 cells results in an increase of ubiquitination of XPC or, perhaps, a decrease of deubiquitination of XPC in response to UV radiation (
Figure 1E). The significant increase in XPC ubiquitination in response to a small knockdown of
OTUD4 (~30% knockdown of OTUD4) indicates a functional link between XPC and OTUD4. Our hypothesis is that the deubiquitinase OTUD4 is involved in XPC recycling by removing ubiquitin from ubiquitinated XPC. Experiments are ongoing to test this hypothesis.
The yeast two-hybrid screening identified 6 XPC-interacting proteins with functions of DNA repair and replication (
Table 1). Interestingly, one of these proteins was XPC itself, indicating a potential dimerization of XPC in the course of NER. DNA damage-inducible transcript 3 (DDIT3) is upregulated in response to a variety of stressors and promotes apoptosis.
BRCA1, a known tumor suppressor gene involved with the repair of double-stranded breaks (DSBs), induces the expression of DDIT3 in response to DNA damage induced by UV and doxorubicin in PC3 cells [
26]. DDIT3 has since been implicated as a transcriptional regulator involved in the apoptotic response to DNA damage and endoplasmic reticulum (ER) stress [
27,
28]. For example, DDIT3 inhibits the expression of CCAAT/enhancer binding proteins (C/EBPs) as a dominant negative and activates the expression of AP-1 through interactions with Jun/Fos AP-1 complex proteins [
29]. DDIT3 regulates apoptosis through regulation of PUMA and BLIM. DDIT3 expression is now used as a marker of ER stress. RPS3A is a ribosomal protein of the 40S subunit of the ribosome. In addition to its functions in ribosomal activity, RPS3A has been shown to upregulate NF-κB activity through chaperoning function [
30,
31] and play a role in preventing apoptosis in association with Bcl-2 and PARP [
32–
35]. Interestingly, RPS3A has been shown to interact with DDIT3. Considering the similar putative roles of RPS3A and DDIT3 in apoptosis in response to stress and interactions with Bcl-2, their interaction with each other, and their interactions with XPC, there is a possibility of a DDIT3-RPS3A-XPC regulated apoptotic pathway in response to DNA damage and other cell stressors. Support for a XPC-regulated UV damage inducible apoptotic pathway lies in the identification of XPC enhancing apoptosis in response to DNA damage [
36]. PHC1 was identified in the yeast two-hybrid screening as interacting with XPC. PHC1 has previously been implicated in DNA damage repair; cells from patients with primary microcephaly (PM), a disease featuring a mutation in PHC1, showed impaired DNA damage repair in response to both ionized radiation and H
2O
2. Compared with control cells, IR of cells from PM patients resulted in a decrease in PHC1 association with chromatin, correlating with a decrease in ubiquitinated H2A and indicating a potential role of PHC1 in chromatin remodeling in response to DNA damage. The overexpression of wild-type PHC1 was sufficient to restore DNA damage repair including the ubiquitination of H2A to cells from PM patients [
37]. Chromatin remodeling factors, including INO80 [
38], Snf5 and BRG1 [
39,
40], components of the SWI/SNF chromatin remodeling complex, have been previously shown to aid in the recruitment of XPC to sites of DNA damage. PHC1 could function similarly in its putative role as a chromatin remodeling factor and recruit XPC to sites of DNA damage. Alternatively, XPC interacts with other chromatin remodeling factors to regulate downstream factors; XPC and hSNF5 interact to recruit and phosphorylate ATM [
18] and it has been proposed that the XPC-BRG1 interaction functions to regulate chromatin relaxation and recruit XPG and PCNA [
41]. The XPC-PHC1 interaction could function similarly in the recruitment of downstream NER factors.
The yeast two-hybrid screening identified 7 XPC-interacting proteins with functions of post-translational modification/proteolysis (
Table 2). OTUD4, a putative deubiquitinase, was identified. Many cellular proteins are stabilized posttranslationally by deubiquitination, which is carried out by a class of enzymes called deubiquitinases (DUBs). DUBs remove the polyubiquitin chains from their substrates and thereby increase their cellular pool. XPC is ubiquitinated after UV, but ubiquitinated XPC does not appear to be directed for degradation [
6]. We speculate that deubiquitination of XPC by OTUD4 could be responsible for the removal of XPC from damage sites, possibly leading to XPC recycling. Indeed, our data presented in
Figure 1 support that there is a functional link between OTUD4 and XPC in human cells. We also identified PSMA4 as interacting with XPC. Although current data indicate that XPC is not sent to the proteasome for degradation following ubiquitination, it has been postulated that XPC interacts directly with the proteasome to signal its degradation. Further, it has been shown that the 19S subunit of the proteasome positively modulates NER while inhibition of the 20S subunit, of which PSMA4 is a part, reduces the recruitment of XPC to damaged sites. This interaction could be responsible for signaling XPC degradation or for recruiting XPC to damaged sites. Ubiquitin-like modifier activating enzyme 3 (UBA3) was identified in the yeast two-hybrid screening. UBA3 binds with NEDD8 activating enzyme E1 subunit 1 (NAE1) to form a heterodimer that serves as the sole E1 enzyme of neddylation and is implicated in cell cycle progression. Since NEDD8 accumulates in response to UV radiation, we speculate that XPC, in its interaction with UBA3, initiates a neddylation cascade in response to its recognition of DNA damage. It has previously been shown that neddylation of histone H4 occurs in response to DNA damage by ionized radiation [
43]; therefore, H4 could also be the target of neddylation in NER.
The yeast two-hybrid screening identified 10 XPC-interacting proteins with functions in signal transduction (
Table 3). The MAPK signaling pathway involves a family of serine threonine kinases activated in response to different cell stressors. Several proteins within the MAPK signaling pathway have been identified as playing a role in GG-NER. For example, p300, a co-transcriptional factor, is phosphorylated by p38 and AKT kinases of the MAPK family and then acetylates histones to allow DNA damage recognition factors such as XPC to access damaged DNA [
50]. Regulation of XPC itself has also been attributed to the MAPK family: p38 MAPK is required for the recruitment of XPC and TFIIH to damaged DNA sites [
51]. The identification of mitogen-activated protein kinase kinase kinase 5 (MAP3K5), a kinase within the MAPK family whose roles include activation of the p38 pathway, as interacting with XPC further supports the idea of the recruitment of XPC to DNA damage sites through MAPK signaling. Further, PTEN, a regulator of AKT signaling within the p38 MAPK pathway, has been shown to positively regulate the transcription of XPC [
52]. Protein tyrosine phosphatase type IVA, member 2 (PTP4A2), also known as PRL2, was recently shown to down regulate PTEN, promoting AKT signaling. It can be extrapolated that this downregulation of PTEN decreases the transcription of XPC and, therefore, the recognition of lesions within GG-NER. Identified by our Y2H screening as interacting with XPC, PTP4A2 could potentially be a negative regulator of XPC and the DNA damage response. Another signaling pathway associated with the regulation of GG-NER is the PKA pathway. PKA, involved in mitotic regulation and chromatin remodeling, has been implicated in DNA damage repair through its phosphorylation of p19INK4d which promotes the DNA damage response after a cell is subject to UV radiation, β-amyloid peptides, and cisplatin; its phosphorylation of Cdc20, a factor involved in DNA damage checkpoints; and its potential phosphorylation of Pol δ, a DNA polymerase involved in DNA repair and replication. PKA comprises two regulatory subunits and two catalytic subunits, all of which have been identified as existing in different isoforms. The yeast two-hybrid screening identified two isoforms of the catalytic subunit of PKA—PRKACB and PRKACA—as potential interactors of XPC. PRKACA, in particular, has been suggested to play a role in the DNA damage response pathway through its translocation-causing phosphorylation of S100A11, a stress response protein associated with p21. PRKACB or PRKACA could stimulate GG-NER through chromatin remodeling or translocation of XPC into the nucleus.
In addition to XPC-interacting proteins with functions of DNA repair and replication, proteolysis and post-translational modifications, and signal transduction, the yeast two-hybrid screening identified 4 XPC-interacting proteins with functions of transcription regulation (
Table 4), 15 XPC-interacting proteins with functions of metabolism (
Table 5), and 7 XPC-interacting proteins with uncategorized functions (
Table 6).



{kind=link}