Optimization of a DNA Nicking Assay to Evaluate Oenocarpus bataua and Camellia sinensis Antioxidant Capacity

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. In Vitro Chemical Antioxidant Capacity of Oenocarpus bataua and Camellia sinensis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | TPC | DPPH | ORAC | FRAP |
|---|---|---|---|---|
| Biological relevance | Reduction of phosphomolybdique and phosphotungstique complexes | DPPH radical scavenging activity | ROO scavenging activity (radical chain breaking) | Fe3+ reduction (Fe2+ production) |
| Extract | (µg GAEq/mg DE) | (µmol TEq/g DE) | (µmol TEq/g DE) | (mmol Fe(II)Eq/g DE) |
| Cs (W) | 364.6 ± 31.8 | 2741 ± 191 | 4941 ± 167 | 7.0 ± 0.3 |
| Ob (W) | 107.6 ± 6.2 | 424 ± 3 | 2189 ± 163 | 1.8 ± 0.1 |
| Cs (M) | 275.5 ± 22.5 | 2927 ± 193 | 6628 ± 86 | 7.5 ± 0.4 |
| Ob (M) | 306.5 ± 26 | 2054 ± 100 | 3708 ± 359 | 4.8 ± 0.1 |
| Cs (A) | 371.3 ± 11.4 | 3486 ± 191 | 6375 ± 107 | 7.9 ± 0.6 |
| Ob (A) | 183.9 ± 7.2 | 1325 ± 99 | 2132 ± 104 | 2.7 ± 0.2 |
2.2. Water pUC18 DNA Nicking Assay

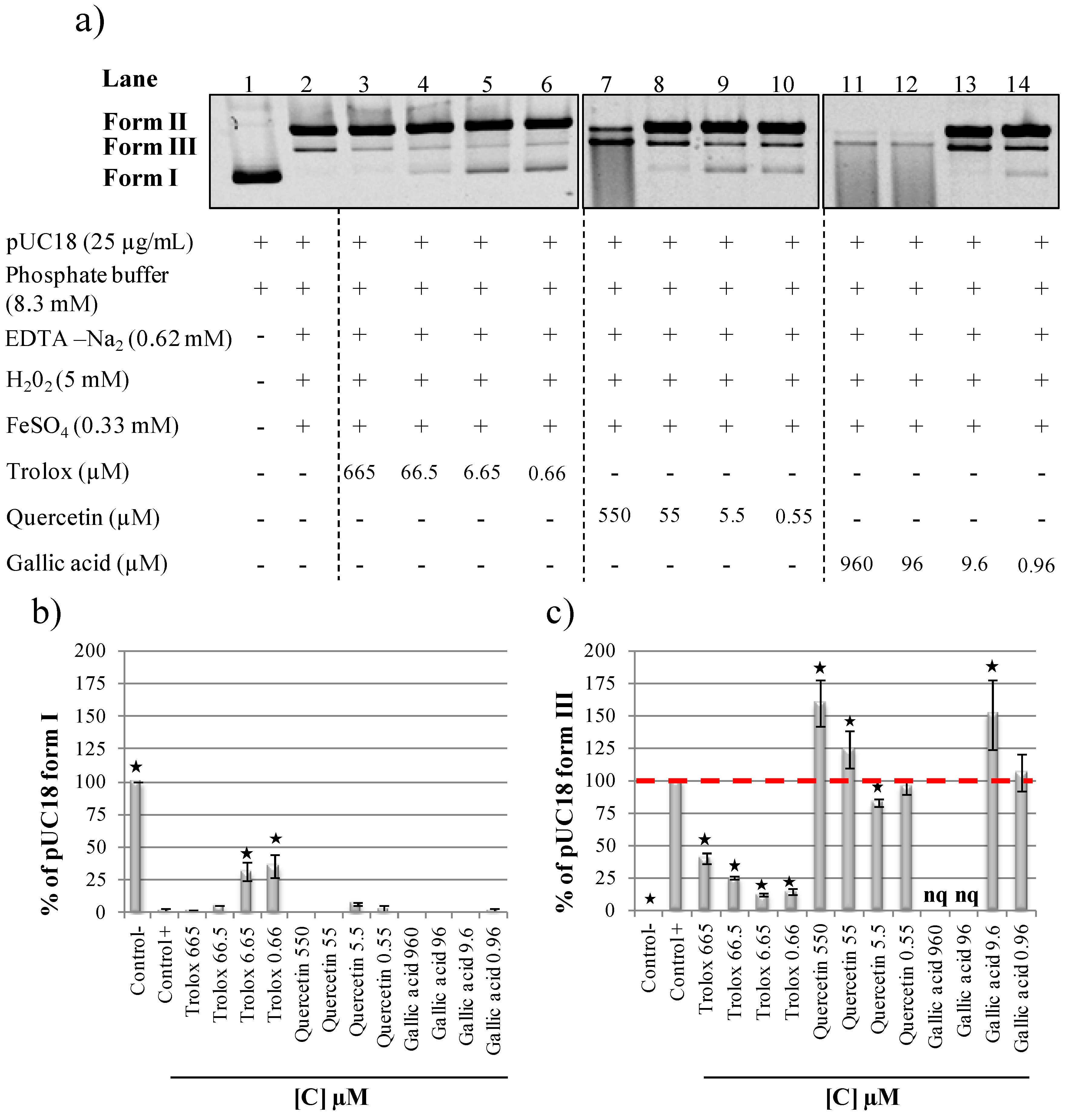
2.3. Evaluation of Antioxidant and Prooxidant Capacity of Trolox, Quercetin and Gallic Acid

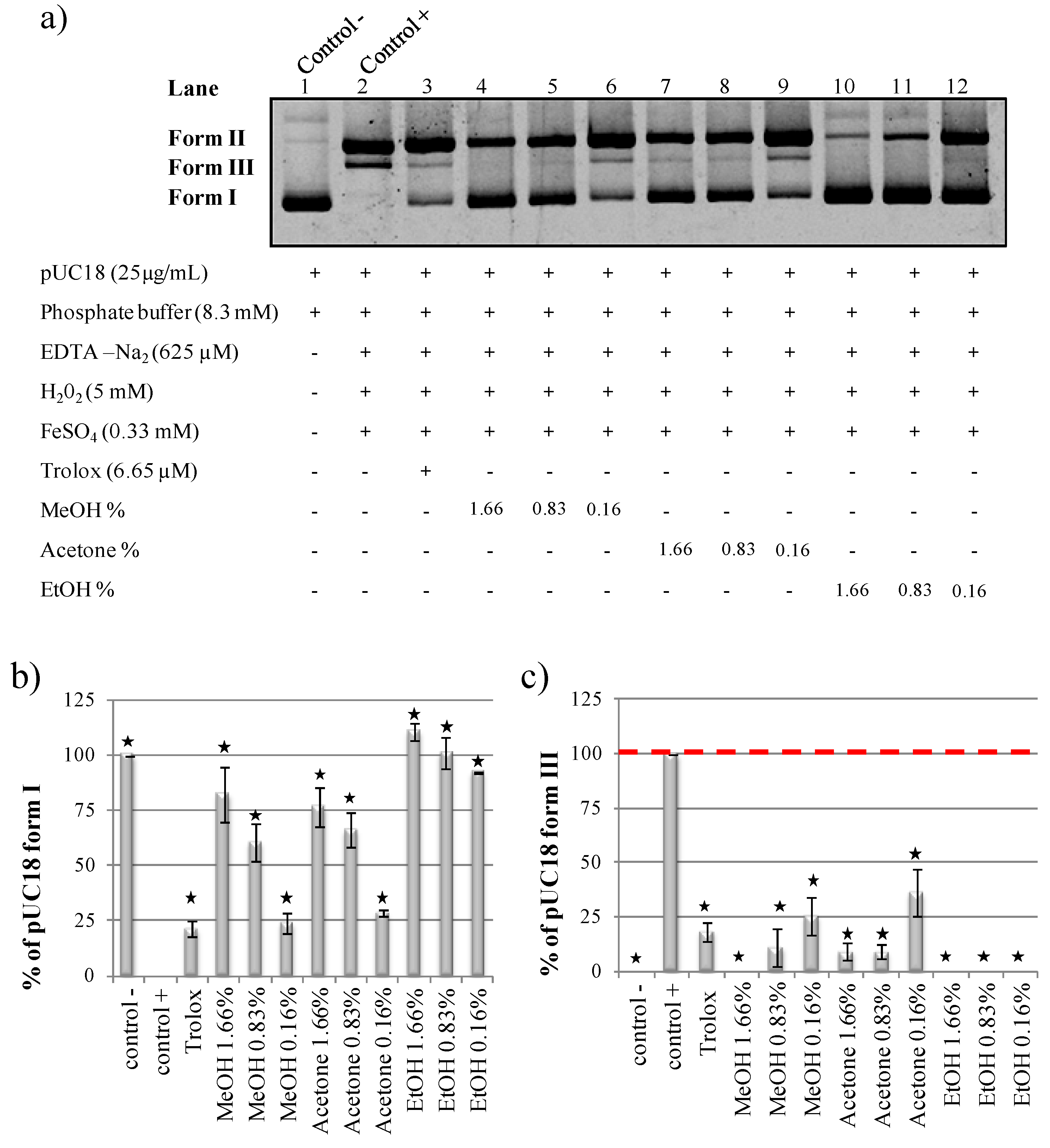
2.4. Organic Solvents Effect on pUC18 DNA Nicking Assay

2.5. Methanol and Acetone pUC18 Strand Breaks Assay

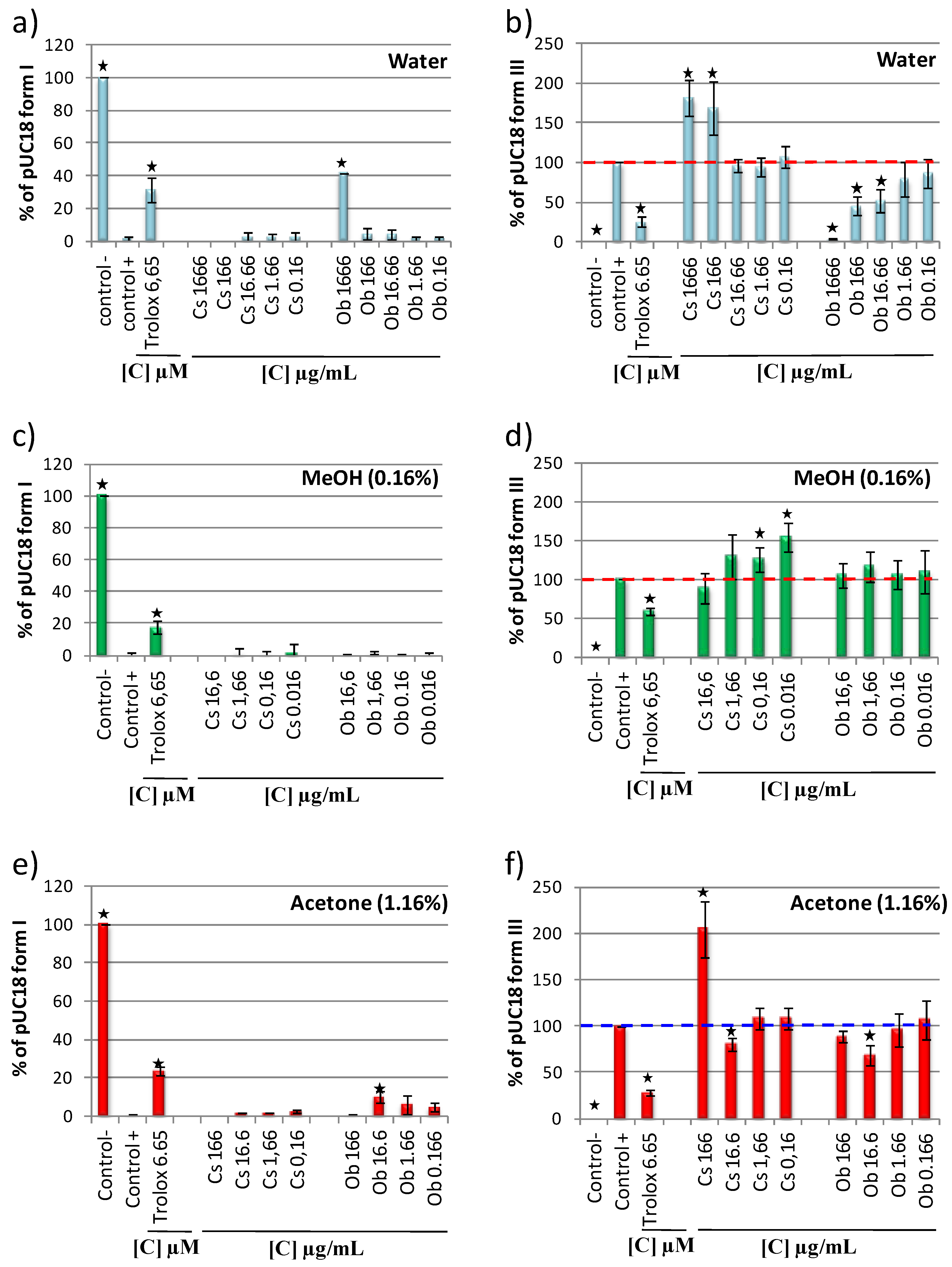
2.6. DNA Antioxidant and Prooxidant Capacity of Camellia sinensis and Oenocarpus bataua

3. Experimental Section
3.1. Plant Material
3.2. Extracts Preparation
3.3. Chemicals
3.4. Optimization of DNA •OH Nicking Assay Conditions of Aqueous and Organic Extracts
3.5. DNA Nicking Assays of Pure Compounds and Plant Extracts
3.6. Quantification of pUC18 Form I and Form III Percentage
3.7. Phytochemical Analysis
Total Phenolic Content (TPC)
3.8. Evaluation of in Vitro Antioxidant Activity by Chemical Assays
3.8.1. DPPH Assay
3.8.2. FRAP Assay
3.8.3. ORAC Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Mitochondrial oxidative stress: implications for cell death. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 143–83. [Google Scholar] [CrossRef] [PubMed]
- Huang, X. Iron overload and its association with cancer risk in humans: Evidence for iron as a carcinogenic metal. Mutat. Res. Mol. Mech. Mutagen. 2003, 533, 153–171. [Google Scholar] [CrossRef]
- Hadjiliadis, N. Cytotoxic, Mutagenic, and Carcinogenic Potential of Heavy Metals Related to Human Environment; Kluwer Academic Press: Dordrecht, The Netherlands, 1997; Volume 26. [Google Scholar]
- Drew, B.; Leeuwenburgh, C. Aging and the role of reactive nitrogen species. Ann. N. Y. Acad. Sci. 2002, 959, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, B. Oxidative stress and the pathogenesis of Alzheimer’s disease. Oxid. Med. Cell. Longev. 2013, 2013, 316523. [Google Scholar] [PubMed]
- Liebert, M.A.; Markesbery, W.R.; Lovell, M.A. DNA Oxidation in Alzheimer’s disease. Antioxid. Redox Signal. 2006, 8, 2039–2045. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant capacity of tea and common vegetables. J. Agric. Food Chem. 1996, 44, 3426–3431. [Google Scholar] [CrossRef]
- Richelle, M.; Tavazzi, I.; Offord, E. Comparison of the antioxidant activity of commonly consumed polyphenolic beverages (coffee, cocoa, and tea) prepared per cup serving. J. Agric. Food Chem. 2001, 49, 3438–3442. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Deemer, E.K.; Prior, R.L.; Huang, D. Novel fluorometric assay for hydroxyl radical prevention capacity using fluorescein as the probe. J. Agric. Food Chem. 2002, 50, 2772–2777. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.D.; Dizdaroglu, M.; Cooke, M.S. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res. 2004, 567, 1–61. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.; Meyerstein, D.; Czapski, G. The Fenton reagents. Free Radic. Biol. Med. 1993, 15, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Kitts, D.D.; Wijewickreme, A.N.; Hu, C. Antioxidant properties of a North American ginseng extract. Mol. Cell. Biochem. 2000, 203, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Flemmig, J.; Arnhold, J. Ferrous ion-induced strand breaks in the DNA plasmid pBR322 are not mediated by hydrogen peroxide. Eur. Biophys. J. 2007, 36, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, M.D.; Bobier, R.T.; Hiatt, T.; Cheng, I.F. Variability of the Fenton reaction characteristics of the EDTA, DTPA, and citrate complexes of iron. Biometals 2003, 16, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Rezaire, A.; Robinson, J.C.; Bereau, D.; Verbaere, A.; Sommerer, N.; Khan, M.K.; Durand, P.; Prost, E.; Fils-Lycaon, B. Amazonian palm Oenocarpus bataua (“patawa”): Chemical and biological antioxidant activity—Phytochemical composition. Food Chem. 2014, 149, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Aviram, M.; Zhang, Y.; Henning, S.M.; Feng, L.; Dreher, M.; Heber, D. Comparison of antioxidant potency of commonly consumed polyphenol-rich beverages in the United States. J. Agric. Food Chem. 2008, 56, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Repine, J.E.; Pfenninger, O.W.; Talmage, D.W.; Berger, E.M.; Pettijohn, D.E. Dimethyl sulfoxide prevents DNA nicking mediated by ionizing radiation or iron/hydrogen peroxide-generated hydroxyl radical. Proc. Natl. Acad. Sci. USA 1981, 78, 1001–1003. [Google Scholar] [CrossRef] [PubMed]
- Girard-Lalancette, K.; Pichette, A.; Legault, J. Sensitive cell-based assay using DCFH oxidation for the determination of pro- and antioxidant properties of compounds and mixtures: Analysis of fruit and vegetable juices. Food Chem. 2009, 115, 720–726. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology. 2002, 177, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.; Hynes, M.J. The kinetics and mechanisms of the complex formation and antioxidant behavior of the polyphenols EGCg and ECG with iron(III). J. Inorg. Biochem. 2007, 101, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, Y.; Yoshinaga, K.; Yoshioka, H.; Yoshioka, H. Kinetic analysis of the effect of (−)-epigallocatechin gallate on the DNA scission induced by Fe(II). Biosci. Biotechnol. Biochem. 2002, 66, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ahmedna, M.; Goktepe, I. Effects of processing methods and extraction solvents on concentration and antioxidant activity of peanut skin phenolics. Food Chem. 2005, 90, 199–206. [Google Scholar] [CrossRef]
- Turkmen, N.; Sari, F.; Velioglu, Y.S. Effects of extraction solvents on concentration and antioxidant activity of black and black mate tea polyphenols determined by ferrous tartrate and Folin–Ciocalteu methods. Food Chem. 2006, 99, 835–841. [Google Scholar] [CrossRef]
- Monod, A.; Chebbi, A.; Durand-jolibois, H.; Carlier, P. Oxidation of methanol by hydroxyl radicals in aqueous solution under simulated cloud droplet conditions. Atmos. Environ. 2000, 34, 5283–5294. [Google Scholar] [CrossRef]
- Luo, Y.; Han, Z.; Chin, S.M.; Linn, S. Three chemically distinct types of oxidants formed by iron-mediated Fenton reactions in the presence of DNA. Proc. Natl. Acad. Sci. USA 1994, 91, 12438–12442. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Wang, S.; Stewart, G.; Hwang, H.; Fu, P.P. Effect of organic solvents and biologically relevant ions on the light-induced DNA cleavage by pyrene and its amino and hydroxy derivatives. Int. J. Mol. Sci. 2002, 3, 937–947. [Google Scholar] [CrossRef]
- Perva-Uzunalić, A.; Škerget, M.; Knez, Ž.; Weinreich, B.; Otto, F.; Grüner, S. Extraction of active ingredients from green tea (Camellia sinensis): Extraction efficiency of major catechins and caffeine. Food Chem. 2006, 96, 597–605. [Google Scholar] [CrossRef]
- Arnous, A.; Makris, D.P.; Kefalas, P. Correlation of pigment and flavanol content with antioxidant properties in selected aged regional wines from Greece. J. Food Compos. Anal. 2002, 15, 655–665. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leba, L.-J.; Brunschwig, C.; Saout, M.; Martial, K.; Vulcain, E.; Bereau, D.; Robinson, J.-C. Optimization of a DNA Nicking Assay to Evaluate Oenocarpus bataua and Camellia sinensis Antioxidant Capacity. Int. J. Mol. Sci. 2014, 15, 18023-18039. https://doi.org/10.3390/ijms151018023
Leba L-J, Brunschwig C, Saout M, Martial K, Vulcain E, Bereau D, Robinson J-C. Optimization of a DNA Nicking Assay to Evaluate Oenocarpus bataua and Camellia sinensis Antioxidant Capacity. International Journal of Molecular Sciences. 2014; 15(10):18023-18039. https://doi.org/10.3390/ijms151018023
Chicago/Turabian StyleLeba, Louis-Jérôme, Christel Brunschwig, Mona Saout, Karine Martial, Emmanuelle Vulcain, Didier Bereau, and Jean-Charles Robinson. 2014. "Optimization of a DNA Nicking Assay to Evaluate Oenocarpus bataua and Camellia sinensis Antioxidant Capacity" International Journal of Molecular Sciences 15, no. 10: 18023-18039. https://doi.org/10.3390/ijms151018023
APA StyleLeba, L.-J., Brunschwig, C., Saout, M., Martial, K., Vulcain, E., Bereau, D., & Robinson, J.-C. (2014). Optimization of a DNA Nicking Assay to Evaluate Oenocarpus bataua and Camellia sinensis Antioxidant Capacity. International Journal of Molecular Sciences, 15(10), 18023-18039. https://doi.org/10.3390/ijms151018023