Signal Transduction of Platelet-Induced Liver Regeneration and Decrease of Liver Fibrosis

{kind=link}
{kind=link}
Abstract
:1. Introduction: Cirrhosis
2. Platelets
3. Platelets and Liver Regeneration
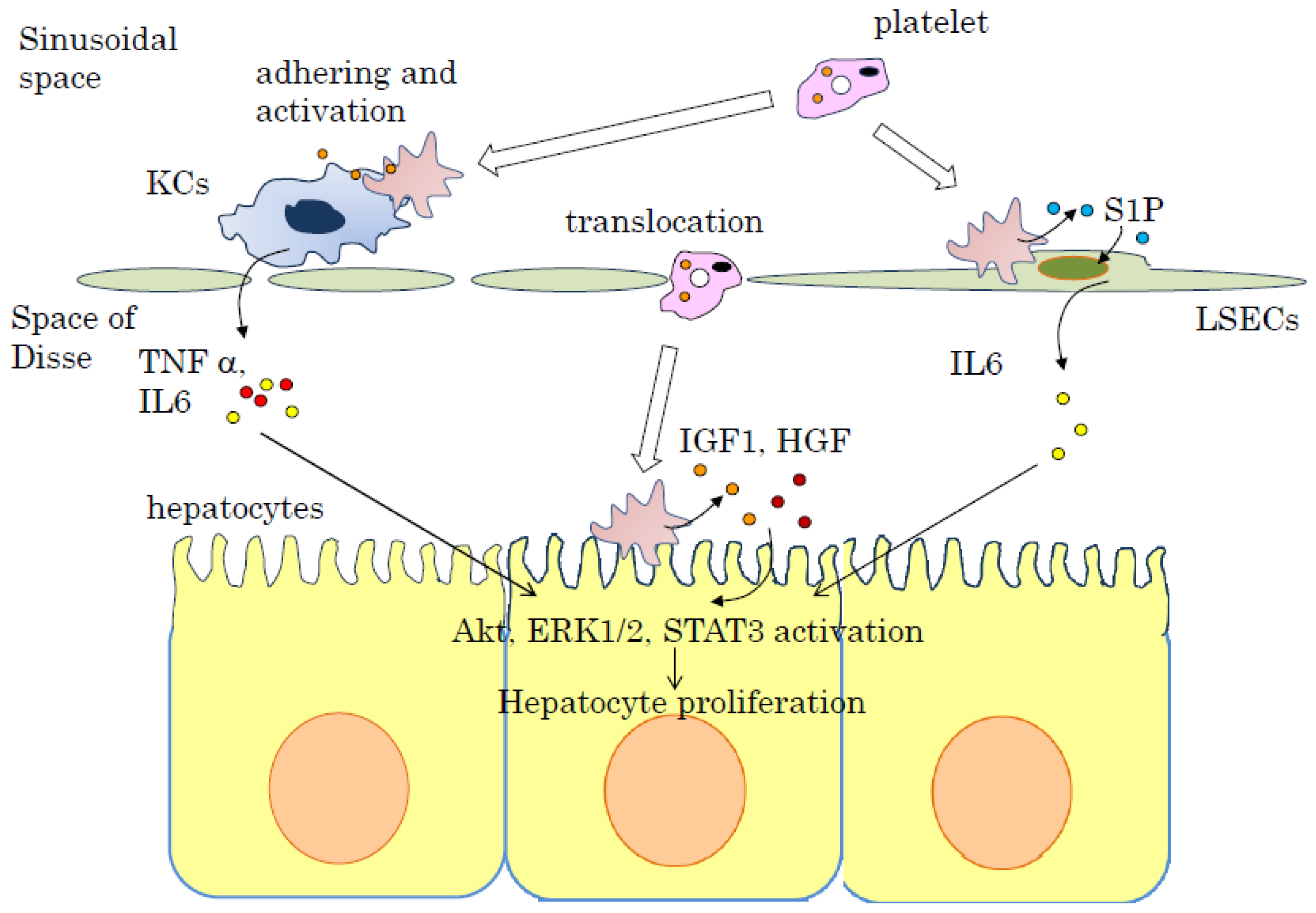
4. Signal Transduction of Liver Regeneration by Platelets
5. Effect of Platelets and Thrombopoietin Receptor Agonist in Liver Cirrhosis
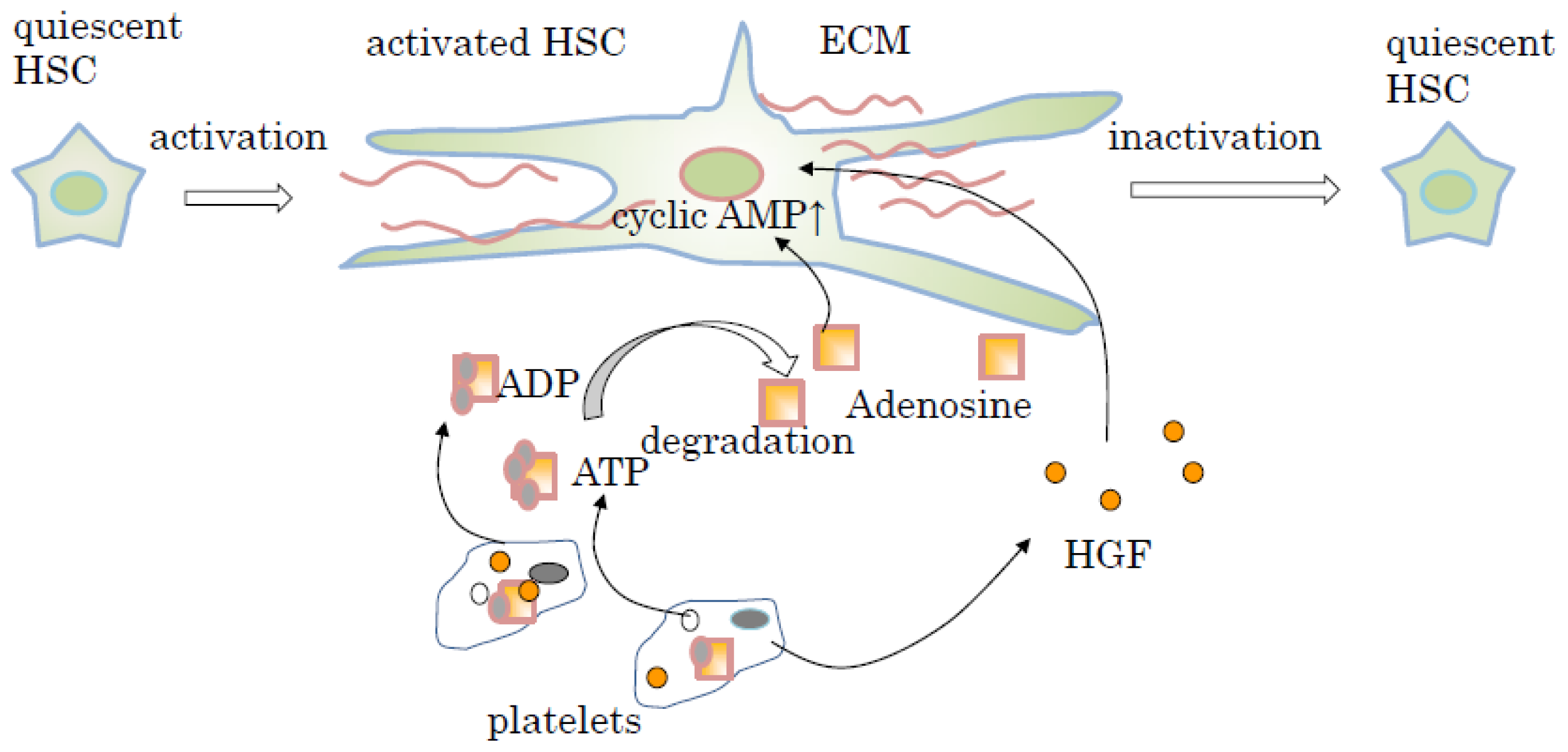
6. Signal Transduction of Liver Fibrolysis Induced by Platelets and Thrombopoietin (TPO)
7. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsSoichiro Murata wrote this manuscript. Takehito Maruyama, Takeshi Nowatari and Kazuhiro Takahashi performed some of the experiments and participated in a discussion. Nobuhiro Ohkohchi supervised this manuscript.
References
- Popper, H. Pathologic aspects of cirrhosis. A review. Am. J. Pathol 1977, 87, 228–264. [Google Scholar]
- Poordad, F. Review article: Thrombocytopenia in chronic liver disease. Aliment. Pharmacol. Ther 2007, 26, S5–S11. [Google Scholar]
- Afdhal, N.; McHutchison, J.; Brown, R.; Jacobson, I.; Manns, M.; Poordad, F.; Weksler, B.; Esteban, R. Thrombocytopenia associated with chronic liver disease. J. Hepatol 2008, 48, 1000–1007. [Google Scholar]
- Witters, P.; Freson, K.; Verslype, C.; Peerlinck, K.; Hoylaerts, M.; Nevens, F.; van Geet, C.; Cassiman, D. Review article: Blood platelet number and function in chronic liver disease and cirrhosis. Aliment. Pharmacol. Ther 2008, 27, 1017–1029. [Google Scholar]
- Bashour, F.N.; Teran, J.C.; Mullen, K.D. Prevalence of peripheral blood cytopenias (hypersplenism) in patients with nonalcoholic chronic liver disease. Am. J. Gastroenterol 2000, 95, 2936–2939. [Google Scholar]
- Merion, R.M.; Schaubel, D.E.; Dykstra, D.M.; Freeman, R.B.; Port, F.K.; Wolfe, R.A. The survival benefit of liver transplantation. Am. J. Transplant 2005, 5, 307–313. [Google Scholar]
- Neuberger, J.; James, O. Guidelines for selection of patients for liver transplantation in the era of donor-organ shortage. Lancet 1999, 354, 1636–1639. [Google Scholar]
- Guarrera, J.V.; Emond, J.C. Advances in segmental liver transplantation: Can we solve the donor shortage? Transplant. Proc 2001, 33, 3451–3455. [Google Scholar]
- Taylor, M.C.; Grieg, P.D.; Detsky, A.S.; McLeod, R.S.; Abdoh, A.; Krahn, M.D. Factors associated with the high cost of liver transplantation in adults. Can. J. Surg 2002, 45, 425–434. [Google Scholar]
- Mueller, A.R.; Platz, K.P.; Kremer, B. Early postoperative complications following liver transplantation. Best Pract. Res. Clin. Gastroenterol 2004, 18, 881–900. [Google Scholar]
- Porrett, P.M.; Hsu, J.; Shaked, A. Late surgical complications following liver transplantation. Liver Transplant 2009, 15, S12–S18. [Google Scholar]
- Chang, Y.; Bluteau, D.; Debili, N.; Vainchenker, W. From hematopoietic stem cells to platelets. J. Thromb. Haemost 2007, 5, S318–S327. [Google Scholar]
- Suzuki, H.; Yamazaki, H.; Tanoue, K. Immunocytochemical aspects of platelet adhesive proteins and membrane glycoproteins during activation. Prog. Histochem. Cytochem 1996, 30, 1–106. [Google Scholar]
- Blair, P.; Flaumenhaft, R. Platelet α-granules: Basic biology and clinical correlates. Blood Rev 2009, 23, 177–189. [Google Scholar]
- McNicol, A.; Israels, S.J. Platelet dense granules: Structure, function and implications for haemostasis. Thromb. Res 1999, 95, 1–18. [Google Scholar]
- Polasek, J. Platelet secretory granules or secretory lysosomes? Platelets 2005, 16, 500–501. [Google Scholar]
- Holmsen, H. Physiological functions of platelets. Ann. Med 1989, 21, 23–30. [Google Scholar]
- Broos, K.; Feys, H.B.; de Meyer, S.F.; Vanhoorelbeke, K.; Deckmyn, H. Platelets at work in primary hemostasis. Blood Rev 2011, 25, 155–167. [Google Scholar]
- Suzuki, H.; Nakamura, S.; Itoh, Y.; Tanaka, T.; Yamazaki, H.; Tanoue, K. Immunocytochemical evidence for the translocation of alpha granule membrane glycoprotein IIb/IIIa (integrin alpha IIb beta 3) of human platelets to the surface membrane during the release reaction. Histochemistry 1992, 97, 381–388. [Google Scholar]
- Mazzucco, L.; Borzini, P.; Gope, R. Platelet-derived factors involved in tissue repair-from signal to function. Transfus. Med. Rev 2010, 24, 218–234. [Google Scholar]
- Ranzato, E.; Balbo, V.; Boccafoschi, F.; Mazzucco, L.; Burlando, B. Scratch wound closure of C2C12 mouse myoblasts is enhanced by human platelet lysate. Cell Biol. Int 2009, 33, 911–917. [Google Scholar]
- Rozman, P.; Bolta, Z. Use of platelet growth factors in treating wounds and soft-tissue injuries. Acta Dermatovenerol. Alp. Panon. Adriat 2007, 16, 156–165. [Google Scholar]
- Yamaguchi, R.; Terashima, H.; Yoneyama, S.; Tadano, S.; Ohkohchi, N. Effects of platelet-rich plasma on intestinal anastomotic healing in rats: PRP concentration is a key factor. J. Surg. Res 2012, 173, 258–266. [Google Scholar]
- Radice, F.; Yánez, R.; Gutiérrez, V.; Rosales, J.; Pinedo, M.; Coda, S. Comparison of magnetic resonance imaging findings in anterior cruciate ligament grafts with and without autologous platelet derived growth factors. Arthroscopy 2010, 26, 50–57. [Google Scholar]
- Dugrillon, A.; Eichler, H.; Kern, S.; Klüter, H. Autologous concentrated platelet-rich plasma (cPRP) for local application in bone regeneration. Int. J. Oral Maxillofac. Surg 2002, 31, 615–619. [Google Scholar]
- Hartmann, E.K.; Heintel, T.; Morrison, R.H.; Weckbach, A. Influence of platelet-rich plasma on the anterior fusion in spinal injuries: A qualitative and quantitative analysis using computer tomography. Arch. Orthop. Trauma Surg 2010, 130, 909–914. [Google Scholar]
- De Vos, R.J.; Weir, A.; van Schie, H.T.; Bierma-Zeinstra, S.M.; Verhaar, J.A.; Weinans, H.; Tol, J.L. Platelet-rich plasma injection for chronic Achilles tendinopathy: A randomized controlled trial. JAMA 2010, 303, 144–149. [Google Scholar]
- Rodeo, S.A.; Delos, D.; Weber, A.; Ju, X.; Cunningham, M.E.; Fortier, L.; Maher, S. What’s new in orthopaedic research. J. Bone Jt. Surg. Am 2010, 92, 2491–2501. [Google Scholar]
- Nocito, A.; Georgiev, P.; Dahm, F.; Jochum, W.; Bader, M.; Graf, R.; Clavien, P.A. Platelets and platelet-derived serotonin promote tissue repair after normothermic hepatic ischemia in mice. Hepatology 2007, 45, 369–376. [Google Scholar]
- McNicol, A.; Israels, S.J. Beyond hemostasis: The role of platelets in inflammation, malignancy and infection. Cardiovasc. Hematol. Disord. Drug Targets 2008, 8, 99–117. [Google Scholar]
- Mehta, P. Potential role of platelets in the pathogenesis of tumor metastasis. Blood 1984, 63, 55–63. [Google Scholar]
- Nash, G.F.; Turner, L.F.; Scully, M.F.; Kakkar, A.K. Platelets and cancer. Lancet Oncol 2002, 3, 425–430. [Google Scholar]
- Elzey, B.D.; Sprague, D.L.; Ratliff, T.L. The emerging role of platelets in adaptive immunity. Cell. Immunol 2005, 238, 1–9. [Google Scholar]
- Sowa, J.M.; Crist, S.A.; Ratliff, T.L.; Elzey, B.D. Platelet influence on T- and B-cell responses. Arch. Immunol. Ther. Exp 2009, 57, 235–241. [Google Scholar]
- Klinger, M.H.; Jelkmann, W. Role of blood platelets in infection and inflammation. J. Interf. Cytokine Res 2002, 22, 913–922. [Google Scholar]
- Sprague, D.L.; Elzey, B.D.; Crist, S.A.; Waldschmidt, T.J.; Jensen, R.J.; Ratliff, T.L. Platelet-mediated modulation of adaptive immunity: Unique delivery of CD154 signal by platelet-derived membrane vesicles. Blood 2008, 111, 5028–5036. [Google Scholar]
- Khandoga, A.; Hanschen, M.; Kessler, J.S.; Krombach, F. CD4+ T cells contribute to postischemic liver injury in mice by interacting with sinusoidal endothelium and platelets. Hepatology 2006, 43, 306–315. [Google Scholar]
- Khandoga, A.; Biberthaler, P.; Messmer, K.; Krombach, F. Platelet-endothelial cell interactions during hepatic ischemia-reperfusion in vivo: A systematic analysis. Microvasc. Res 2003, 65, 71–77. [Google Scholar]
- Pak, S.; Kondo, T.; Nakano, Y.; Murata, S.; Fukunaga, K.; Oda, T.; Sasaki, R.; Ohkohchi, N. Platelet adhesion in the sinusoid caused hepatic injury by neutrophils after hepatic ischemia reperfusion. Platelets 2010, 21, 282–288. [Google Scholar]
- Nakano, Y.; Kondo, T.; Matsuo, R.; Hashimoto, I.; Kawasaki, T.; Kohno, K.; Myronovych, A.; Tadano, S.; Hisakura, K.; Ikeda, O.; et al. Platelet dynamics in the early phase of postischemic liver in vivo. J. Surg. Res. 2008, 149, 192–198. [Google Scholar]
- Zaldivar, M.M.; Pauels, K.; von Hundelshausen, P.; Berres, M.L.; Schmitz, P.; Bornemann, J.; Kowalska, M.A.; Gassler, N.; Streetz, K.L.; Weiskirchen, R.; et al. CXC chemokine ligand 4 (Cxcl4) is a platelet-derived mediator of experimental liver fibrosis. Hepatology 2010, 51, 1345–1353. [Google Scholar]
- Laschke, M.W.; Dold, S.; Menger, M.D.; Jeppsson, B.; Thoelacius, H. Platelet-dependent accumulation of leukocytes in sinusoids mediates hepatocellular damage in bile duct ligation-induced cholestasis. Br. J. Pharmacol 2008, 153, 148–156. [Google Scholar]
- Lang, P.A.; Contalado, C.; Gergiev, P.; El-Bardy, A.M.; Recher, M.; Kurrer, M.; Cervantes-Barragan, L.; Ludewig, B.; Calzascia, T.; Bolinger, B.; et al. Aggravation of viral hepatitis by platelet-derived serotonin. Nat. Med 2008, 14, 756–761. [Google Scholar]
- Murata, S.; Ohkohchi, N.; Matsuo, R.; Ikeda, O.; Myronovych, A.; Hoshi, R. Platelets promote liver regeneration in early period after hepatectomy in mice. World J. Surg 2007, 31, 808–816. [Google Scholar]
- Lesurtel, M.; Graf, R.; Aleil, B.; Walther, D.J.; Tian, Y.; Jochum, W.; Gachet, C.; Bader, M.; Clavien, P.A. Platelet-derived serotonin mediates liver regeneration. Science 2006, 312, 104–107. [Google Scholar]
- Wolber, E.M.; Jelkmann, W. Thrombopoietin: The novel hepatic hormone. News Physiol. Sci 2002, 17, 6–10. [Google Scholar]
- Cheng, G. Eltrombopag, a thrombopoietin-receptor agonist in the treatment of adult chronic immune thrombocytopenia: A review of the efficacy and safety profile. Ther. Adv. Hematol 2012, 3, 155–164. [Google Scholar]
- Murata, S.; Matsuo, R.; Ikeda, O.; Myronovych, A.; Watanabe, M.; Hisakura, K.; Nakano, Y.; Hashimoto, I.; Ohkohchi, N. Platelets promote liver regeneration under conditions of kupffer cell depletion after hepatectomy in mice. World J. Surg 2008, 32, 1088–1096. [Google Scholar]
- Myronovych, A.; Murata, S.; Chiba, M.; Matsuo, R.; Ikeda, O.; Watanabe, M.; Hisakura, K.; Nakano, Y.; Kohno, K.; Kawasaki, T.; et al. Role of platelets on liver regeneration after 90% hepatectomy in mice. J. Hepatol 2008, 49, 363–372. [Google Scholar]
- Matsuo, R.; Nakano, Y.; Ohkohchi, N. Platelet administration via the portal vein promotes liver regeneration in rats after 70% hepatectomy. Ann. Surg 2011, 253, 759–763. [Google Scholar]
- Murata, S.; Hashimoto, I.; Nakano, Y.; Myronovych, A.; Watanabe, M.; Ohkohchi, N. Single administration of thrombopoietin prevents progression of liver fibrosis and promotes liver regeneration after partial hepatectomy in cirrhotic rats. Ann. Surg 2008, 248, 821–828. [Google Scholar]
- Hisakura, K.; Murata, S.; Fukunaga, K.; Myronovych, A.; Tadano, S.; Kawasaki, T.; Kohno, K.; Kobayashi, E.; Saito, T.; Yasue, H.; et al. Platelets prevent acute liver damage after extended hepatectomy in pigs. J. Hepatobiliary Pancreat. Sci 2010, 17, 855–864. [Google Scholar]
- Hisakura, K.; Murata, S.; Takahashi, K.; Matsuo, R.; Pak, S.; Ikeda, N.; Kawasaki, T.; Kohno, K.; Myronovych, A.; Nakano, Y.; et al. Platelets prevent acute hepatitis induced by anti-Fas antibody. J. Gastroenterol. Hepatol 2011, 26, 348–355. [Google Scholar]
- Braet, F.; Wisse, E. Structural and functional aspects of liver sinusoidal endothelial cell fenestrae: A review. Comp. Hepatol 2002, 23, 1:1–1:17. [Google Scholar]
- Wack, K.E.; Ross, M.A.; Zegarra, V.; Sysko, L.R.; Watkins, S.C.; Stolz, D.B. Sinusoidal ultrastructure evaluated during the revascularization of regenerating rat liver. Hepatology 2001, 33, 363–378. [Google Scholar]
- Nakamura, M.; Shibasaki, M.; Nitta, Y.; Endo, Y. Translocation of platelets into Disse space and their entry into hepatocytes in response to lipopolysaccharides, interleukin-1 and tumour necrosis factor: The role of Kupffer cells. J. Hepatol 1998, 28, 991–999. [Google Scholar]
- Endo, Y.; Nakamura, M. The effect of lipopolysaccharide, interleukin-1 and tumour necrosis factor on the hepatic accumulation of 5-hydroxytryptamine and platelets in the mouse. Br. J. Pharmacol 1992, 105, 613–619. [Google Scholar]
- Ohtaki, Y.; Shimauchi, H.; Yokochi, T.; Endo, Y. In vivo platelet response to lipopolysaccharide in mice: Proposed method for evaluating new antiplatelet drugs. Thromb. Res 2003, 108, 303–309. [Google Scholar]
- Nakamura, T.; Nishizawa, T.; Hagiya, M.; Seki, T.; Shimonishi, M.; Sugimura, A.; Tashiro, K.; Shimizu, S. Molecular cloning and expression of human hepatocyte growth factor. Nature 1989, 342, 440–443. [Google Scholar]
- Matsuo, R.; Ohkohchi, N.; Murata, S.; Ikeda, O.; Nakano, Y.; Watanabe, M.; Hisakura, K.; Myronovych, A.; Kubota, T.; Narimatsu, H.; et al. Platelets strongly induce hepatocyte proliferation with IGF-1 and HGF in vitro. J. Surg. Res. 2008, 145, 279–286. [Google Scholar]
- Ozaki, M.; Haga, S.; Zhang, H.Q.; Irani, K.; Suzuki, S. Inhibition of hypoxia/reoxygenation-induced oxidative stress in HGF-stimulated antiapoptotic signaling: Role of PI3-K and Akt kinase upon rac1. Cell Death Differ 2003, 10, 508–515. [Google Scholar]
- Conery, A.R.; Cao, Y.; Thompson, E.A.; Townsend, C.M., Jr.; Ko, T.C.; Luo, K. Akt interacts directly with Smad3 to regulate the sensitivity to TGF-β induced apoptosis. Nat. Cell Biol 2004, 6, 366–372. [Google Scholar]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocr. Rev 2001, 22, 153–183. [Google Scholar]
- Gauldie, J.; Richards, C.; Baumann, H. IL-6 and the acute phase reaction. Res. Immunol 1992, 143, 755–759. [Google Scholar]
- Montalvo-Jave, E.E.; Escalante-Tattersfield, T.; Ortega-Salgado, J.A.; Piña, E.; Geller, D.A. Factors in the pathophysiology of the liver ischemia-reperfusion injury. J. Surg. Res 2008, 147, 153–159. [Google Scholar]
- Croner, R.S.; Hoerer, E.; Kulu, Y.; Hackert, T.; Gebhard, M.M.; Herfarth, C.; Klar, E. Hepatic platelet and leukocyte adherence during endotoxemia. Crit. Care 2006, 10, R15:1–R15:6. [Google Scholar]
- Kawasaki, T.; Murata, S.; Takahashi, K.; Nozaki, R.; Ohshiro, Y.; Ikeda, N.; Pak, S.; Myronovych, A.; Hisakura, K.; Fukunaga, K.; et al. Activation of human liver sinusoidal endothelial cell by human platelets induces hepatocyte proliferation. J. Hepatol 2010, 53, 648–654. [Google Scholar]
- Yatomi, Y.; Ohmori, T.; Rile, G.; Kazama, F.; Okamoto, H.; Sano, T.; Satoh, K.; Kume, S.; Tigyi, G.; Igarashi, Y.; et al. Sphingosine-1-phosphate as a major bioactive lysophospholipid that is released from platelets and interacts with endothelial cells. Blood 2000, 96, 3431–3438. [Google Scholar]
- Takuwa, Y.; Okamoto, Y.; Yoshioka, K.; Takuwa, N. Sphingosine-1-phosphate signaling and biological activities in the cardiovascular system. Biochim. Biophys. Acta 2008, 1781, 483–488. [Google Scholar]
- Zheng, D.M.; Kitamura, T.; Ikejima, K.; Enomoto, N.; Yamashina, S.; Suzuki, S.; Takei, Y.; Sato, N. Sphingosine-1-phosphate protects rat liver sinusoidal endothelial cells from ethanol-induced apoptosis: Role of intracellular calcium and nitric oxide. Hepatology 2006, 44, 1278–1287. [Google Scholar]
- Fernández-Pisonero, I.; Dueňas, A.I.; Barreiro, O.; Montero, O.; Sánchez-Madrid, F.; Garcia-Rodriguez, C. Lipopolysaccharide and sphingosine-1-phosphate cooperate to induce inflammatory molecules and leukocyte adhesion in endothelial cells. J. Immunol 2012, 189, 5402–5410. [Google Scholar]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int 2006, 26, 1175–1186. [Google Scholar]
- Meijer, C.; Wiezer, M.J.; Diehl, A.M.; Schouten, H.J.; Schouten, H.J.; Meijer, S.; van Rooijen, N.; van Lambalgen, A.A.; Dijkstra, C.D.; van Leeuwen, P.A. Kupffer cell depletion by CI2MDP-liposomes alters hepatic cytokine expression and delays liver regeneration after partial hepatectomy. Liver 2000, 20, 66–77. [Google Scholar]
- Akerman, P.; Cote, P.; Yang, S.Q.; McClain, C.; Nelson, S.; Bagby, G.J.; Diehl, A.M. Antibodies to tumor necrosis factor-α inhibit liver regeneration after partial hepatectomy. Am. J. Physiol 1992, 263, G579–G585. [Google Scholar]
- Yamada, Y.; Webber, E.M.; Kirillova, I.; Peschon, J.J.; Fausto, N. Analysis of liver regeneration in mice lacking type 1 or type 2 tumor necrosis factor receptor: Requirement for type 1 but not type 2 receptor. Hepatology 1998, 28, 959–970. [Google Scholar]
- Yamada, Y.; Kirillova, I.; Peschon, J.J.; Fausto, N. Initiation of liver growth by tumor necrosis factor: Deficient liver regeneration in mice lacking type I tumor necrosis factor receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 1441–1446. [Google Scholar]
- Malik, R.; Selden, C.; Hodgson, H. The role of non-parenchymal cells in liver growth. Semin. Cell Dev. Biol 2002, 13, 425–431. [Google Scholar]
- Abshagen, K.; Eipel, C.; Kalff, J.C.; Menger, M.D.; Vollmar, B. Loss of NF-κB activation in Kupffer cell-depleted mice impairs liver regeneration after partial hepatectomy. Am. J. Physiol. Gastrointest. Liver Physiol 2007, 292, G1570–G1577. [Google Scholar]
- Takahashi, K.; Kozuma, Y.; Suzuki, H.; Tamura, T.; Maruyama, T.; Fukunaga, K.; Murata, S.; Ohkohchi, N. Human platelets promote liver regeneration with Kupffer cells in SCID mice. J. Surg. Res 2013, 180, 62–72. [Google Scholar]
- Cooper, K.L.; Fitzgerald, P.; Dillingham, K.; Helme, K.; Akehurst, R. Romiplostim and eltrombopag for immune thrombocytopenia: Methods for indirect comparison. Int. J. Technol. Assess. Health Care 2012, 28, 249–258. [Google Scholar]
- McHutchison, J.G.; Dusheiko, G.; Shiffman, M.L.; Rodriguez-Torres, M.; Sigal, S.; Bourliere, M.; Berg, T.; Gordon, S.C.; Campbell, F.M.; Theodore, D.; et al. Eltrombopag for thrombocytopenia in patients with cirrhosis associated with hepatitis C. N. Engl. J. Med 2007, 357, 2227–2236. [Google Scholar]
- Afdhal, N.H.; Giannini, E.G.; Tayyab, G.; Mohsin, A.; Lee, J.W.; Andriulli, A.; Jeffers, L.; McHutchison, J.; Chen, P.J.; Han, K.H.; et al. Eltrombopag before procedures in patients with cirrhosis and thrombocytopenia. N. Engl. J. Med 2012, 367, 716–724. [Google Scholar]
- Kawaguchi, T.; Komori, A.; Seike, M.; Fujiyama, S.; Watanabe, H.; Tanaka, M.; Sakisaka, S.; Nakamuta, M.; Sasaki, Y.; Oketani, M.; et al. Efficacy and safety of eltrombopag in Japanese patients with chronic liver disease and thrombocytopenia: A randomized, open-label, phase II study. J. Gastroenterol 2012, 47, 1342–1351. [Google Scholar]
- Watanabe, M.; Murata, S.; Hashimoto, I.; Nakano, Y.; Ikeda, O.; Aoyagi, Y.; Matsuo, R.; Fukunaga, K.; Yasue, H.; Ohkohchi, N. Platelets contribute to the reduction of liver fibrosis in mice. J. Gastroenterol. Hepatol 2009, 24, 78–89. [Google Scholar]
- Arthur, M.J.; Fibrogenesis, I.I. Metalloproteinases and their inhibitors in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol 2000, 279, G245–G249. [Google Scholar]
- Benyon, R.C.; Arthur, M.J. Extracellular matrix degradation and the role of hepatic stellate cells. Semin. Liver Dis 2001, 21, 373–384. [Google Scholar]
- Winwood, P.J.; Schuppan, D.; Iredale, J.P.; Kawser, C.A.; Docherty, A.J.; Arthur, M.J.P. Kupffer cell-derived 95-kd type IV collagenase/gelatinase B: Characterization and expression in cultured cells. Hepatology 1995, 22, 304–315. [Google Scholar]
- Hemmann, S.; Graf, J.; Roderfeld, M.; Roeb, E. Expression of MMPs and TIMPs in liver fibrosis—A systematic review with special emphasis on anti-fibrotic strategies. J. Hepatol 2007, 46, 955–975. [Google Scholar]
- Kodama, T.; Takehara, T.; Hikita, H.; Shimizu, S.; Li, W.; Miyagi, T.; Hosui, A.; Tatsumi, T.; Ishida, H.; Tadokoro, S.; et al. Thrombocytopenia exacerbates cholestasis-induced liver fibrosis in mice. Gastroenterology 2010, 138, 2487–2498. [Google Scholar]
- Takahashi, K.; Murata, S.; Fukunaga, K.; Ohkohchi, N. Human platelets inhibit liver fibrosis in severe combined immunodeficiency mice. World J. Gastroenterol 2013, 19, 5250–5260. [Google Scholar]
- Maruyama, T.; Murata, S.; Takahashi, K.; Tamura, T.; Nozaki, R.; Ikeda, N.; Fukunaga, K.; Oda, T.; Sasaki, R.; Ohkohchi, N. Platelet transfusion improves liver function in patients with chronic liver disease and cirrhosis. Tohoku J. Exp. Med 2013, 229, 213–220. [Google Scholar]
- Gressner, A.M.; Weiskirchen, R. Modern pathogenetic concepts of liver fibrosis suggest stellate cells and TGF-β as major players and therapeutic targets. J. Cell. Biol. Med 2006, 10, 76–99. [Google Scholar]
- Friedman, S.L. The cellular basis of hepatic fibrosis—Mechanisms and treatment strategies. N. Engl. J. Med 1993, 328, 1828–1835. [Google Scholar]
- Friedman, S.L.; Maher, J.J.; Bissell, D.M. Mechanisms and therapy of hepatic fibrosis: Report of the AASLD single topic basic research conference. Hepatology 2000, 32, 1403–1408. [Google Scholar]
- Friedman, S.L. Mechanisms of hepatic fibrogenesis. Gastroenterology 2008, 134, 1655–1669. [Google Scholar]
- Ikeda, N.; Murata, S.; Maruyama, T.; Tamura, T.; Nozaki, R.; Kawasaki, T.; Fukunaga, K.; Oda, T.; Sasaki, R.; Homma, M.; et al. Platelet-derived adenosine 5′-triphosphate suppresses activation of human hepatic stellate cell: In vitro study. Hepatol. Res 2012, 42, 91–102. [Google Scholar]


© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Murata, S.; Maruyama, T.; Nowatari, T.; Takahashi, K.; Ohkohchi, N. Signal Transduction of Platelet-Induced Liver Regeneration and Decrease of Liver Fibrosis. Int. J. Mol. Sci. 2014, 15, 5412-5425. https://doi.org/10.3390/ijms15045412
Murata S, Maruyama T, Nowatari T, Takahashi K, Ohkohchi N. Signal Transduction of Platelet-Induced Liver Regeneration and Decrease of Liver Fibrosis. International Journal of Molecular Sciences. 2014; 15(4):5412-5425. https://doi.org/10.3390/ijms15045412
Chicago/Turabian StyleMurata, Soichiro, Takehito Maruyama, Takeshi Nowatari, Kazuhiro Takahashi, and Nobuhiro Ohkohchi. 2014. "Signal Transduction of Platelet-Induced Liver Regeneration and Decrease of Liver Fibrosis" International Journal of Molecular Sciences 15, no. 4: 5412-5425. https://doi.org/10.3390/ijms15045412
APA StyleMurata, S., Maruyama, T., Nowatari, T., Takahashi, K., & Ohkohchi, N. (2014). Signal Transduction of Platelet-Induced Liver Regeneration and Decrease of Liver Fibrosis. International Journal of Molecular Sciences, 15(4), 5412-5425. https://doi.org/10.3390/ijms15045412