Neuroprotective Effect of Melatonin against Kainic Acid-Induced Oxidative Injury in Hippocampal Slice Culture of Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
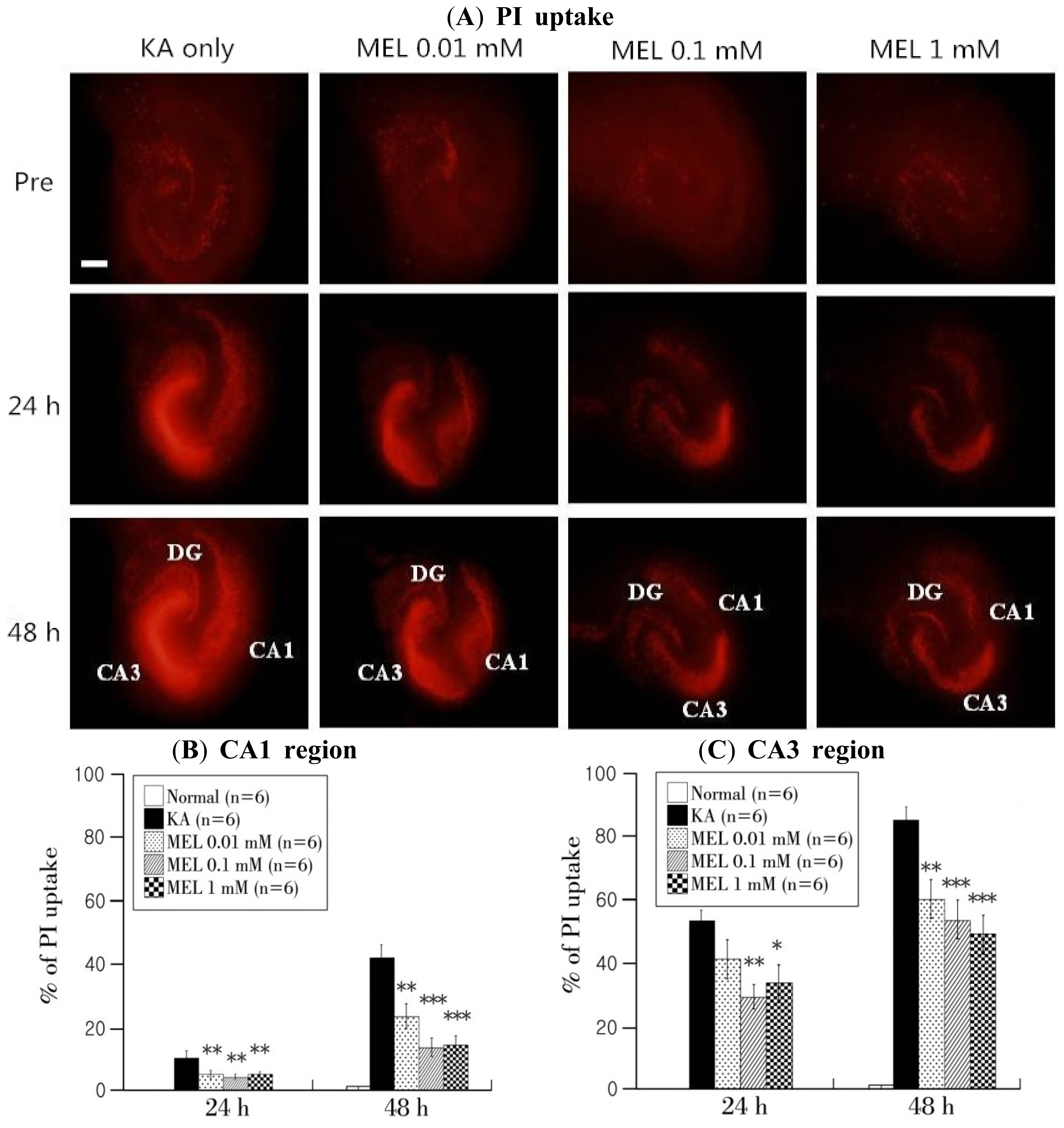
2.1. Effects of Melatonin on KA-Induced Neuronal Toxicity in OHSCs
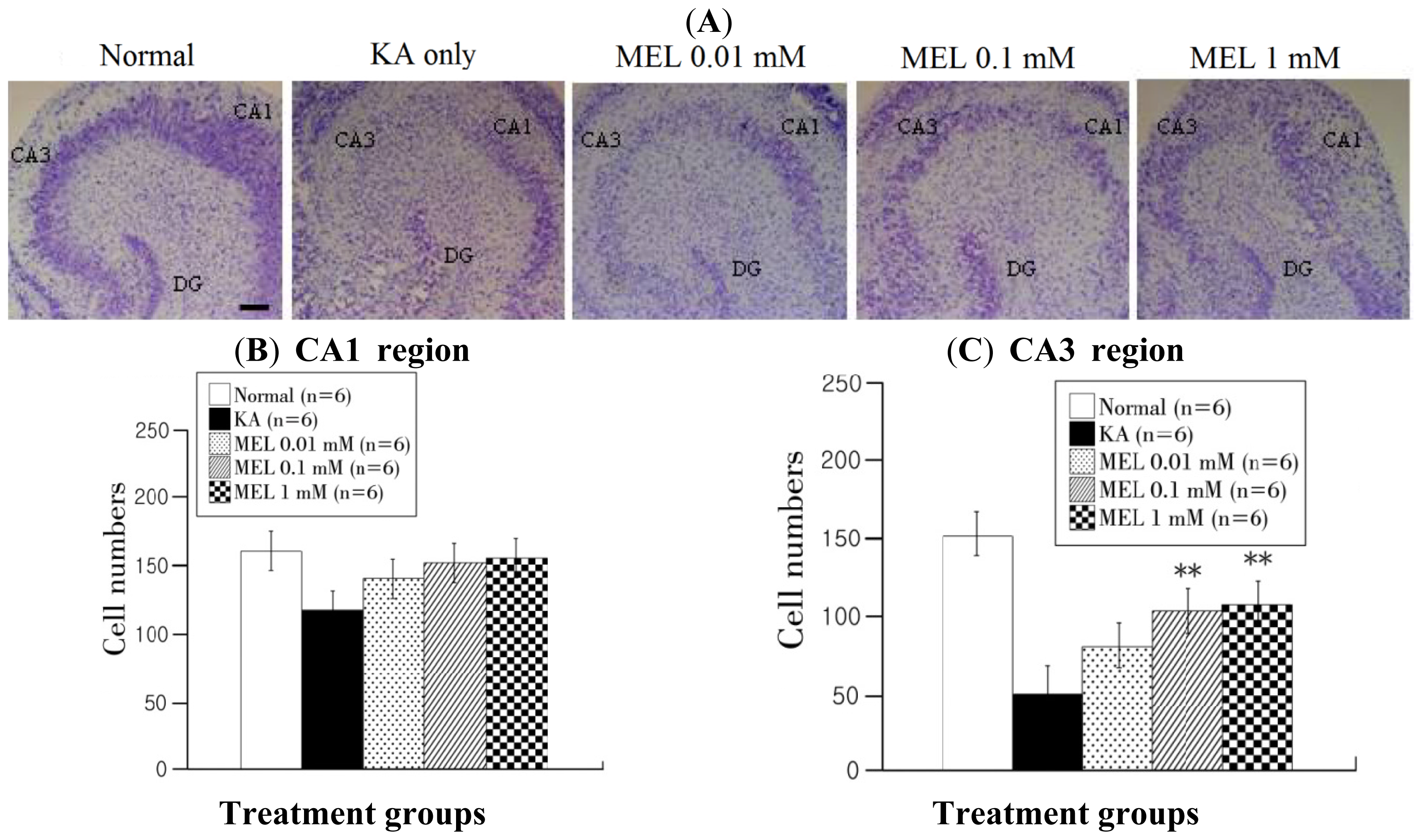
2.2. Cresyl Violet Staining
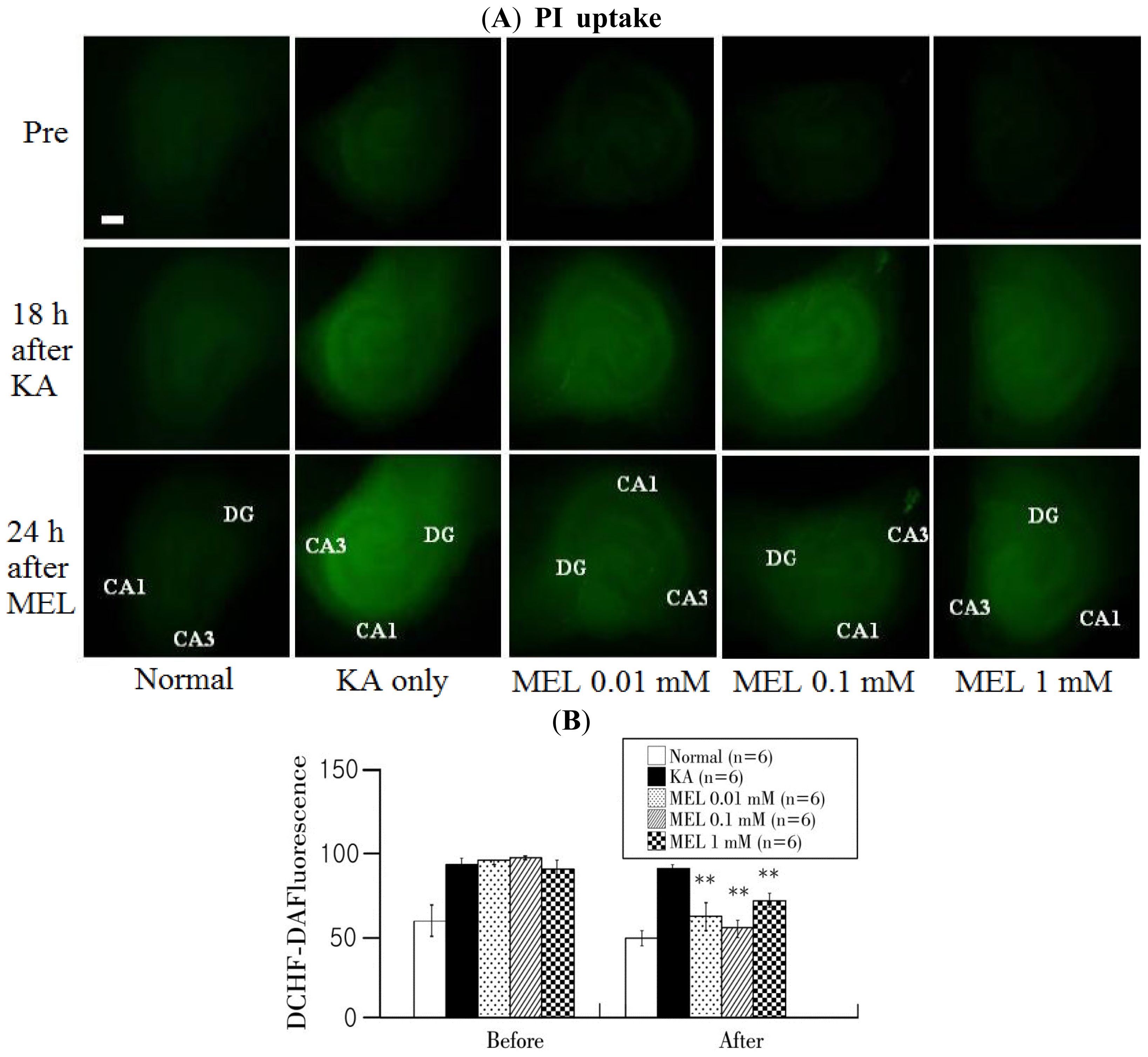
2.3. Formation of ROS in KA-Induced Oxidative Stress
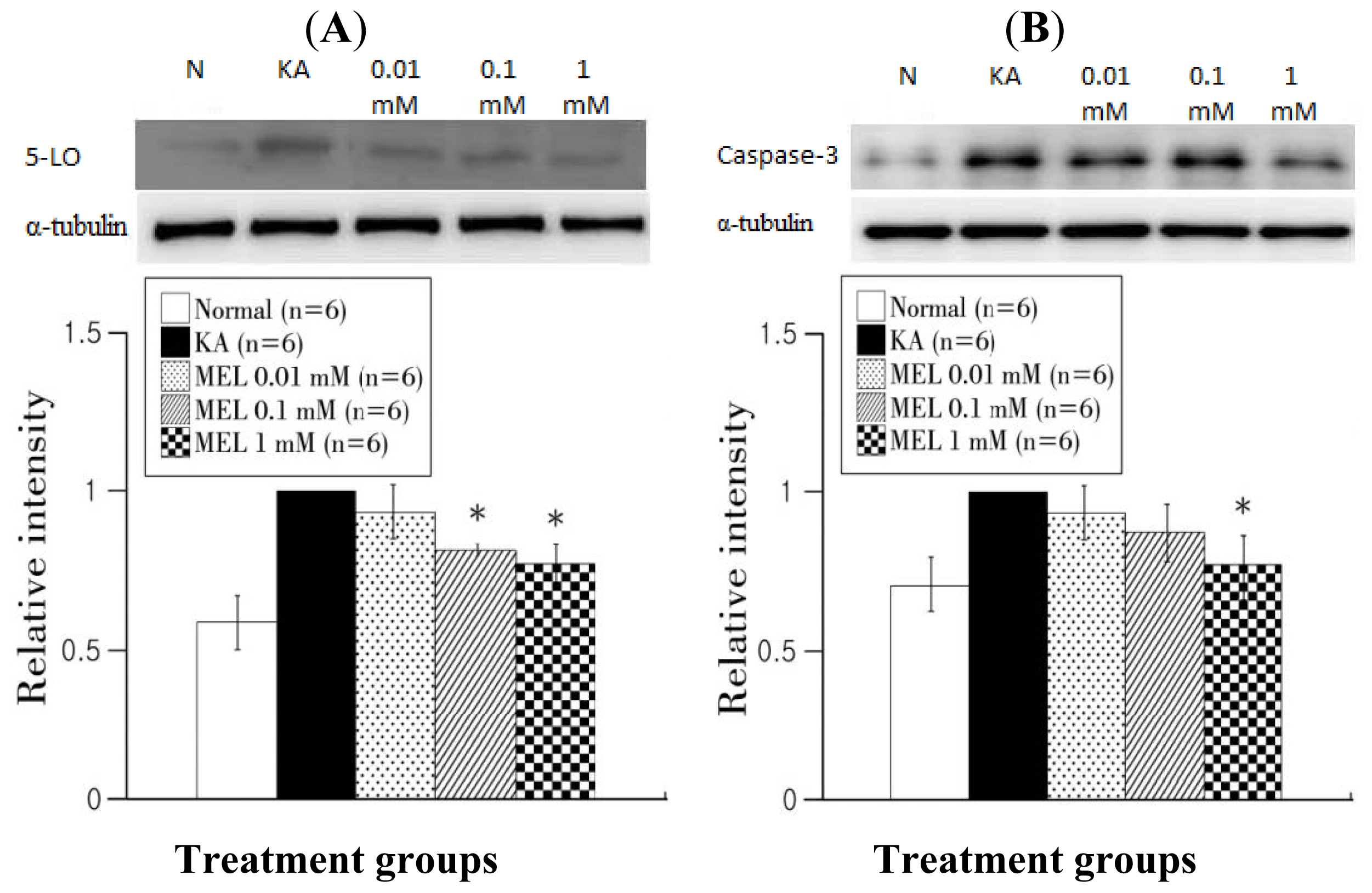
2.4. Western Blotting
2.5. Discussion
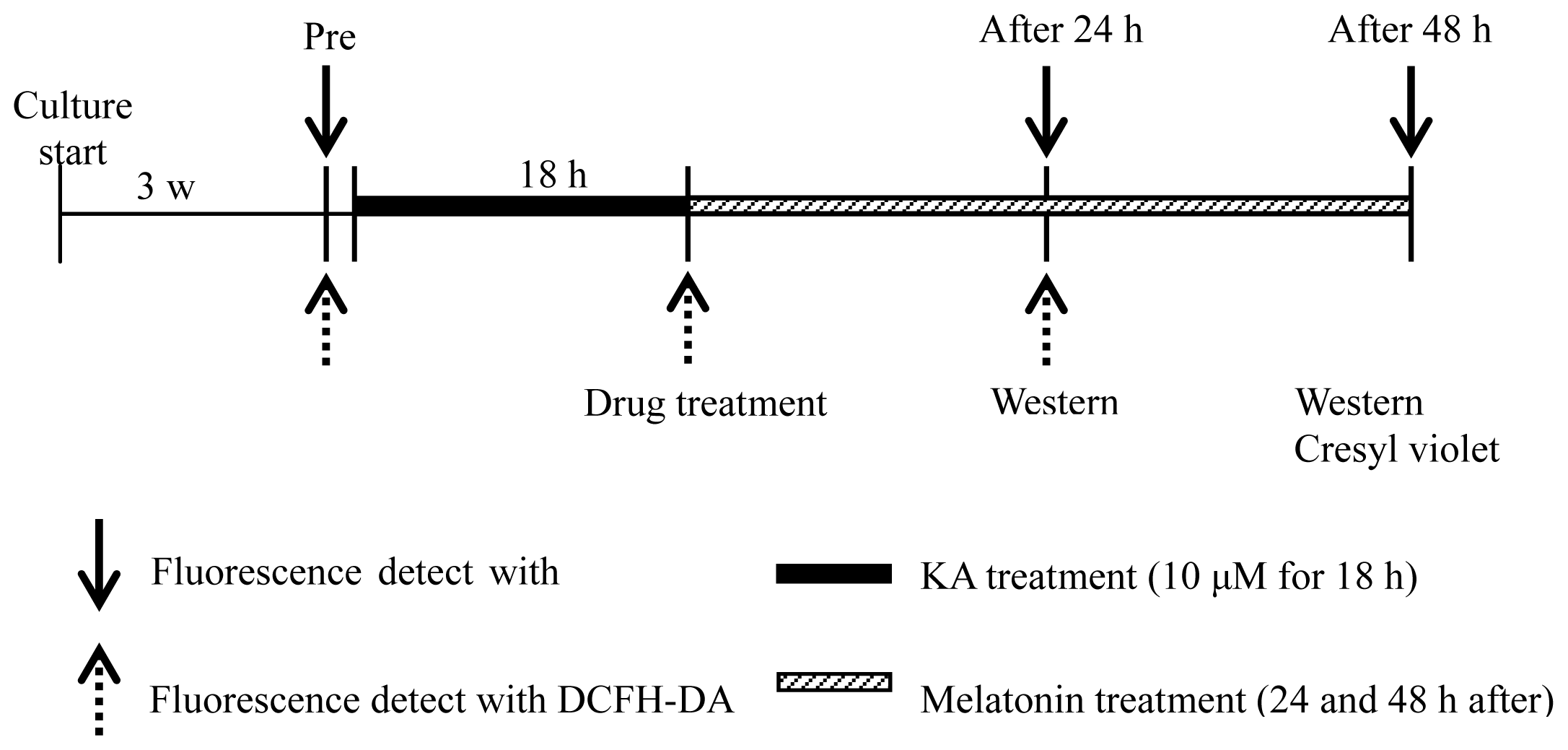
3. Experimental Section
3.1. Organotypic Hippocampal Slice Culture (OHSC)
3.2. Drug Preparation and Treatment
3.3. Measurement of Neuronal Injury
3.4. Evaluation of Intracellular ROS Formation
3.5. Cresyl Violet Staining
3.6. Western Blot Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsAll authors contributed to the conception and design. H.A.K. conducted the experiments. Manuscript was drafted by H.A.K. and edited by K.H.L. and B.H.L.
References
- Floyd, R.A.; Carney, J.M. Free radical damage to protein and DNA: Mechanisms involved and relevant observations on brain undergoing oxidative stress. Ann. Neurol 1992, 32, 22–27. [Google Scholar]
- Floyd, R.A. Antioxidants, oxidative stress, and degenerative neurological disorders. Proc. Soc. Exp. Biol. Med 1999, 222, 236–245. [Google Scholar]
- Liang, L.P.; Ho, Y.S.; Patel, M. Mitochondrial superoxide production in kainate-induced hippocampal damage. Neuroscience 2000, 101, 563–570. [Google Scholar]
- Alvira, D.; Tajes, M.; Verdaguer, E. Inhibition of the cdk5/p25 fragment formation may explain the antiapoptotic effects of melatonin in an experimental model of Parkinsons disease. J. Pineal Res 2006, 40, 251–258. [Google Scholar]
- Mayo, J.C.; Sainz, R.M.; Tan, D.X.; Antolín, I.; Rodríguez, C.; Reiter, R.J. Melatonin and Parkinson’s disease. Endocrine 2005, 27, 169–178. [Google Scholar]
- Srinivasan, V.; Pandi-Perumal, S.R.; Maestroni, G.J.; Esquifino, A.I.; Hardeland, R.; Cardinali, D.P. Role of melatonin in neurodegenerative diseases. Neurotox. Res 2005, 7, 293–318. [Google Scholar]
- Antolin, I.; Rodriquez, C.; Sainz, R.M.; Mayo, J.C.; Aria, H.; Kotter, M. Neurohormone melatonin prevents cell damage: Effect on gene expression for antioxidant enzymes. FASEB J 1996, 10, 882–890. [Google Scholar]
- Hashimoto, S.; Minami, N.; Takakura, R. Low oxygen tension during in vitro maturation is beneficial for supporting the subsequent development of bovine cumulus-oocyte complexes. Mol. Reprod. Dev 2000, 57, 353–360. [Google Scholar]
- Costa, E.J.; Lopes, R.H.; Lamy-Freund, M.T. Permeability of pure lipid bilayers to melatonin. J. Pineal Res 1995, 19, 123–126. [Google Scholar]
- Ekmekcioglu, C. Melatonin receptors in humans: Biological role and clinical relevance. Biomed. Pharmacother 2006, 60, 97–108. [Google Scholar]
- Hung, M.W.; Tipoe, G.L.; Poon, A.M.; Reiter, R.J.; Fung, M.L. Protective effect of melatonin against hippocampal injury of rats with intermittent hypoxia. J. Pineal Res 2008, 44, 214–221. [Google Scholar]
- Cheung, R.T.; Tipoe, G.L.; Tam, S.; Ma, E.S.; Zou, L.Y.; Chan, P.S. Preclinical evaluation of pharmacokinetics and safety of melatonin in propylene glycol for intravenous administration. J. Pineal Res 2006, 41, 337–343. [Google Scholar]
- Cheung, R.T. The utility of melatonin in reducing cerebral damage resulting from ischemia and reperfusion. J. Pineal Res 2003, 34, 153–160. [Google Scholar]
- Hardeland, R. Antioxidative protection by melatonin: Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005, 27, 119–130. [Google Scholar]
- Reiter, R.J. Oxidative damage in the central nervous system: Protection by melatonin. Prog. Neurobiol 1998, 56, 359–384. [Google Scholar]
- Kristensen, B.W.; Noraberg, J.; Zimmer, J. Comparison of excitotoxic profiles of ATPA, AMPA, KA and NMDA in organotypic hippocampal slice cultures. Brain Res 2001, 917, 21–44. [Google Scholar]
- Kim, E.J.; Won, R.; Sohn, J.-H.; Chung, M.-A.; Nam, T.S.; Lee, H.-J.; Lee, B.H. Anti-oxidant effect of ascorbic and dehydroascorbic acids in hippocampal slice culture. Biochem. Biophys. Res. Commun 2008, 366, 8–14. [Google Scholar]
- Won, R.; Lee, K.H.; Lee, B.H. Coenzyme Q10 protects neurons against neurotoxicity in hippocampal slice culture. Neuroreport 2011, 22, 721–726. [Google Scholar]
- Jung, N.Y.; Lee, K.H.; Won, R.; Lee, B.H. Neuroprotective effects of α-tocotrienol on kainic acid-induced neurotoxicity in organotypic hippocampal slice cultures. Int. J. Mol. Sci 2013, 14, 18256–18268. [Google Scholar]
- Zaja-Milatovic, S.; Gupta, R.C.; Aschner, M.; Montine, T.J.; Milatovic, D. Pharmacologic suppression of oxidative damage and dendritic degeneration following kainic acid-induced excitotoxicity in mouse cerebrum. Neurotoxicology 2008, 29, 621–627. [Google Scholar]
- Kotler, M.; Rodriquez, C.; Sainz, R.M.; Antolin, I.; Menendez-Pelaez, A. Melatonin increases gene expression for antioxidant enzymes in rat brain cortex. J. Pineal Res 1998, 24, 83–89. [Google Scholar]
- Juliana, B.H.; Rudimar, L.F.; Ana, P.H.; Ricardo, A.C.; Andressa, B.; Maria, M.C. Amyloid-β neurotoxicity in organotypic culture is attenuated by melatonin: Involvement of GSK-3β, tau and neuroinflammation. J. Pineal Res 2010, 48, 230–238. [Google Scholar]
- Chung, S.Y.; Han, S.H. Melatonin attenuates kainic acid-induced hippocampal neurodeneration and oxidative stress through microglial inhibition. J. Pineal Res 2003, 34, 95–102. [Google Scholar]
- Holopainen, I.E.; Jarvela, J.; Lopez-Picon, F.R.; Pelliniemi, L.J.; Kukko-Lukjanov, T.K. Mechanisms of kainate-induced region-specific neuronal death in immature organotypic hippocampal slice cultures. Neurochem. Int 2004, 45, 1–10. [Google Scholar]
- Noraberg, J.; Kristensen, B.W.; Zimmer, J. Markers for neuronal degeneration in organotypic slice cultures. Brain Res. Protoc 1999, 3, 278–290. [Google Scholar]
- Vornov, J.J.; Tasker, R.C.; Coyle, J.T. Direct observation of the agonist-specific regional vulnerability to glutamate, NMDA, and kainate neurotoxicity in organotypic hippocampal cultures. Exp. Neurol 1991, 114, 11–22. [Google Scholar]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med 2002, 33, 337–349. [Google Scholar]
- Flavia, R.; Laura, P.; Milena, D.N.; Claudia, C.; Sergio, A.; Annalida, B. Rapid and transient stimulation of intracellular reactive oxygen species by melatonin in normal and tumor leukocytes. Toxicol. Appl. Pharmacol 2009, 239, 37–45. [Google Scholar]
- Werz, O.; Szellas, D.; Steinhilber, D. Reactive oxygen species released from granulocytes stimulate 5-lipoxygenase activity in a B-lymphocytic cell line. Eur. J. Biochem 2000, 267, 1263–1269. [Google Scholar]
- Liu, W.; Liu, R.; Schreiber, S.S.; Baudry, M. Role of polyamine metabolism in kainic acid excitotoxicity in organotypic hippocampal slice cultures. J. Neurochem 2001, 79, 976–984. [Google Scholar]
- Monyer, H.; Hartley, D.M.; Choi, D.W. 21-Aminosteroids attenuate excitotoxic neuronal injury in cortical cell cultures. Neuron 1990, 5, 121–126. [Google Scholar]
- Ries, W.L.; Key, L.L., Jr.; Rodriguiz, R.M. Nitroblue tetrazolium reduction and bone resorption by osteoclasts in vitro inhibited by a manganese-based superoxide dismutase mimic. J. Bone Miner. Res 1992, 7, 931–939. [Google Scholar]
- Yalcin, A.; Kilinc, E.; Kocturk, S.; Resmi, H.; Sozmen, E.Y. Effect of melatonin cotreatment against kainic acid on coenzyme Q10, lipid peroxidation and Trx mRNA in rat hippocampus. Int. J. Neurosci 2004, 114, 1085–1097. [Google Scholar]
- Stoppini, L.; Buchs, P.A.; Muller, D.A. Simple method for organotypic cultures of nervous tissue. J. Neurosci. Methods 1991, 37, 173–182. [Google Scholar]





© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, H.A.; Lee, K.H.; Lee, B.H. Neuroprotective Effect of Melatonin against Kainic Acid-Induced Oxidative Injury in Hippocampal Slice Culture of Rats. Int. J. Mol. Sci. 2014, 15, 5940-5951. https://doi.org/10.3390/ijms15045940
Kim HA, Lee KH, Lee BH. Neuroprotective Effect of Melatonin against Kainic Acid-Induced Oxidative Injury in Hippocampal Slice Culture of Rats. International Journal of Molecular Sciences. 2014; 15(4):5940-5951. https://doi.org/10.3390/ijms15045940
Chicago/Turabian StyleKim, Hyung A, Kyung Hee Lee, and Bae Hwan Lee. 2014. "Neuroprotective Effect of Melatonin against Kainic Acid-Induced Oxidative Injury in Hippocampal Slice Culture of Rats" International Journal of Molecular Sciences 15, no. 4: 5940-5951. https://doi.org/10.3390/ijms15045940
APA StyleKim, H. A., Lee, K. H., & Lee, B. H. (2014). Neuroprotective Effect of Melatonin against Kainic Acid-Induced Oxidative Injury in Hippocampal Slice Culture of Rats. International Journal of Molecular Sciences, 15(4), 5940-5951. https://doi.org/10.3390/ijms15045940