Novel and Functional DNA Sequence Variants within the GATA6 Gene Promoter in Ventricular Septal Defects

Abstract
:1. Introduction
2. Results and Discussion
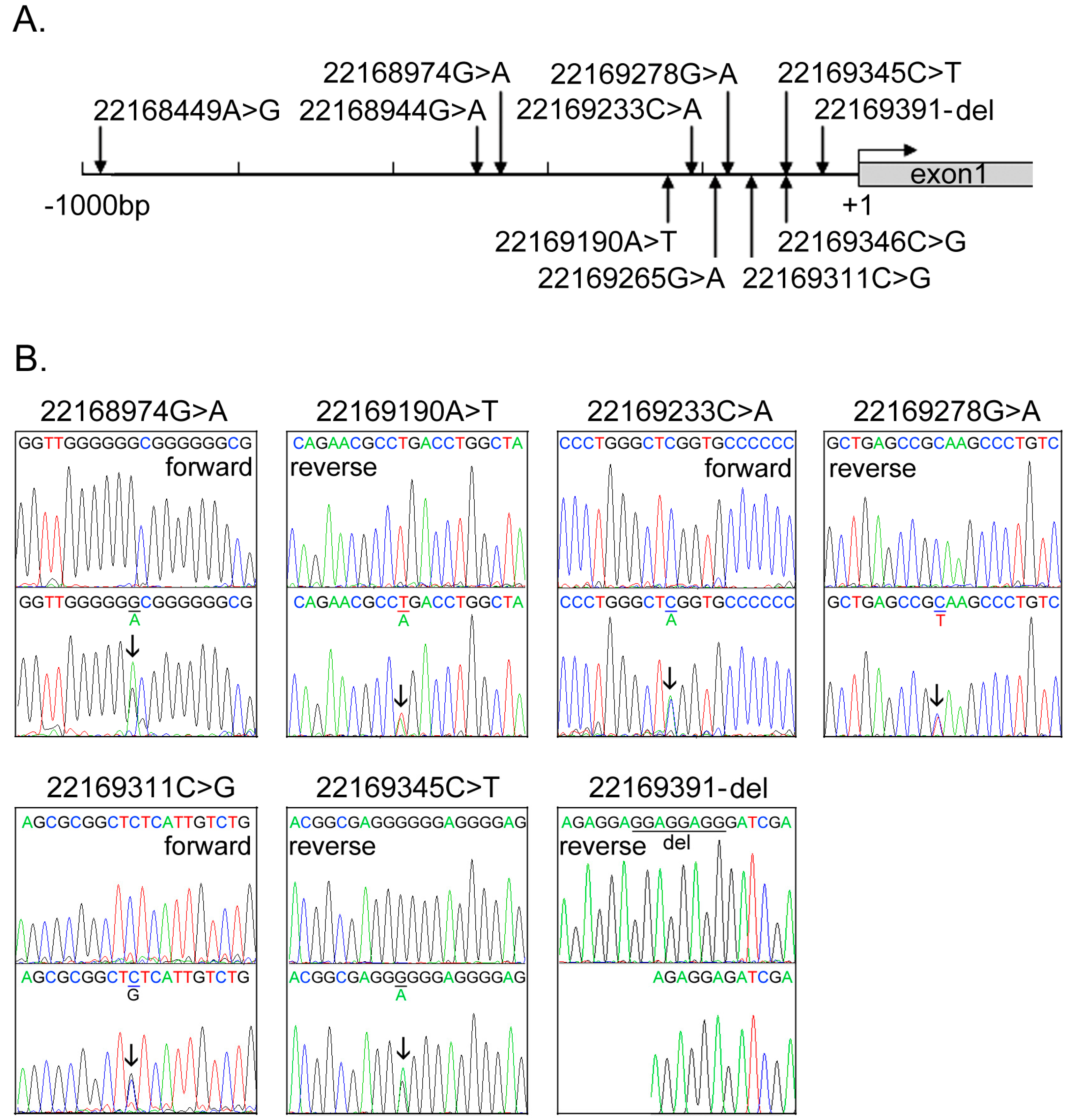
2.1. DNA Sequence Variants (DSVs) Identified in Ventricular Septal Defect (VSD) Patients and Controls

{kind=link}
{kind=link}
| DSVs | Genotype | Location a | Controls ( n = 365) | VSD ( n = 359) | p Value |
|---|---|---|---|---|---|
| g.22168449A>G (rs189133474) | AG | −994 bp | 10 | 6 | 0.329 |
| g.22168944G>A (rs144923558) | GG | −499 bp | 347 | 333 | 0.212 |
| GA | 18 | 24 | |||
| AA | 0 | 2 | |||
| g.22168974G>A | GA | −469 bp | 1 | 0 | – |
| g.22169190A>T | AT | −253 bp | 0 | 1 | – |
| g.22169233C>A | CA | −210 bp | 1 | 0 | – |
| g.22169265G>A (rs146748749) | GG | −178 bp | 347 | 333 | 0.212 |
| GA | 18 | 24 | |||
| AA | 0 | 2 | |||
| g.22169278G>A | GA | −165 bp | 1 | 0 | – |
| g.22169311C>G | CG | −132 bp | 0 | 1 | – |
| g.22169345C>T | CT | −96 bp | 5 | 7 | 0.541 |
| g.22169346C>G (rs139399350) | CC | −97 bp | 316 | 313 | 0.330 |
| CG | 48 | 43 | |||
| GG | 1 | 3 | |||
| g.22169391-del | CCTCCTCC/- | −45–52 bp | 1 | 0 | – |
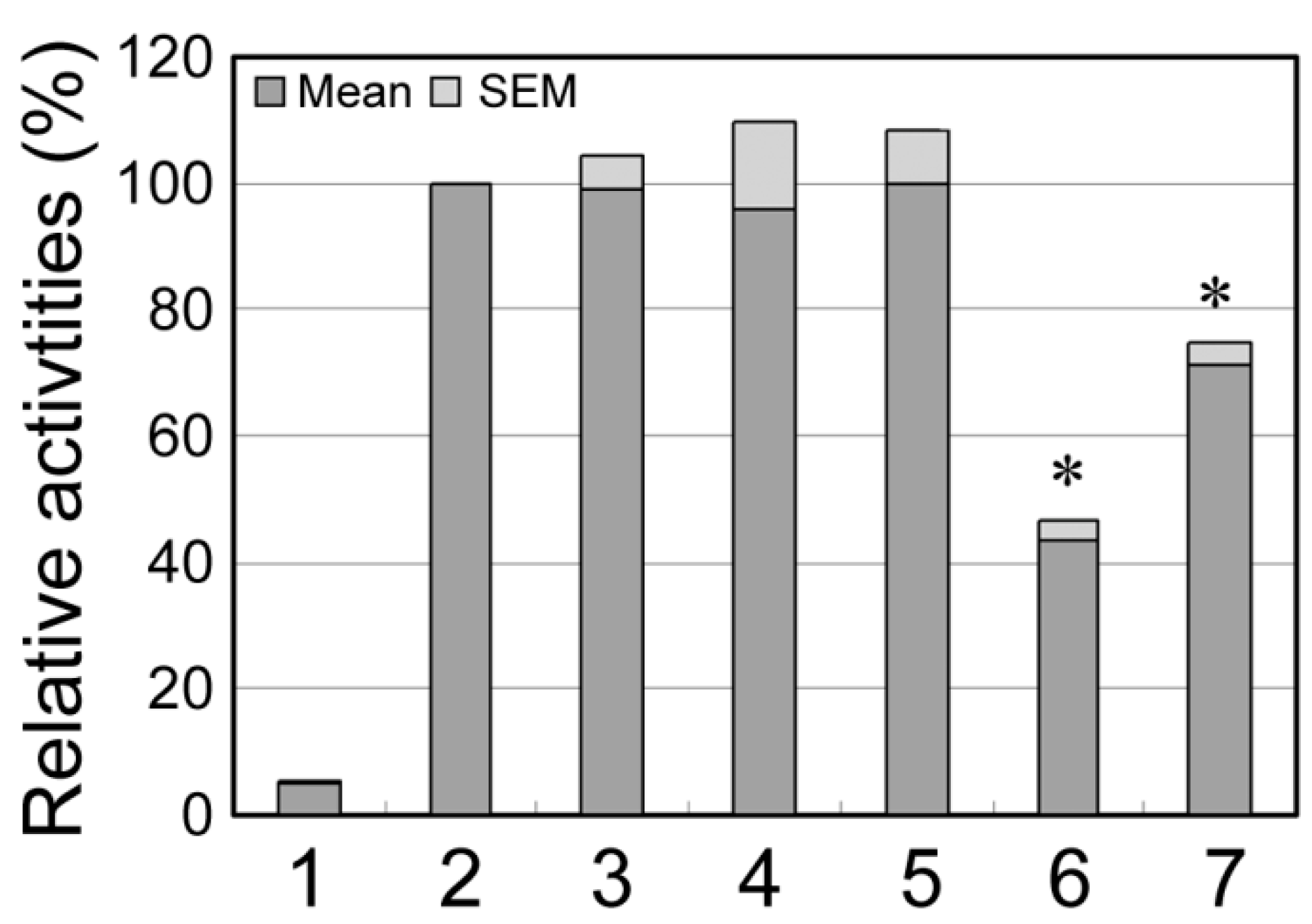
2.2. Functional Analysis of the DSVs

2.3. Discussion
3. Materials and Methods
3.1. Patients and Controls
3.2. Sequence Analysis
| Primers | Sequences | Location | Products |
|---|---|---|---|
| Sequencing | |||
| GATA6-F1 | 5'-ACCAGAGCCTAAACGCTTTC-3' | 22168197 | 695 bp |
| GATA6-R1 | 5'-ACCCTATCTCGGGATGCTAC-3' | 22168891 | |
| GATA6-F2 | 5'-CCGAAACCACCACGACCTGAG-3' | 22168849 | 663 bp |
| GATA6-R2 | 5'-TGGGCTCCTGATTGGACTCACC-3' | 22169511 | |
| Functioning | |||
| GATA6-F | 5'-(KpnI)-ACGCCTCTTGTCCTAAAGTCTC-3' | 22168318 | 1173 bp |
| GATA6-R | 5'-(HindIII)-CGAGCCCTAAACAAACAGC-3' | 22169490 |
3.3. Functional Analysis
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hoffman, J.I.; Kaplan, S. The incidence of congenital heart disease. J. Am. Coll. Cardiol. 2002, 39, 1890–1900. [Google Scholar] [CrossRef]
- Bruneau, B.G. The developmental genetics of congenital heart disease. Nature 2008, 451, 943–948. [Google Scholar] [CrossRef]
- Kathiresan, S.; Srivastava, D. Genetics of human cardiovascular disease. Cell 2012, 148, 1242–1257. [Google Scholar] [CrossRef]
- Burch, J.B. Regulation of GATA gene expression during vertebrate development. Semin. Cell Dev. Biol. 2005, 16, 71–81. [Google Scholar] [CrossRef]
- Peterkin, T.; Gibson, A.; Loose, M.; Patient, R. The roles of GATA4, GATA5 and GATA6 in vertebrate heart development. Semin. Cell Dev. Biol. 2005, 16, 83–94. [Google Scholar] [CrossRef]
- Pikkarainen, S.; Tokola, H.; Kerkelä, R.; Ruskoaho, H. GATA transcription factors in the developing and adult heart. Cardiovasc. Res. 2004, 63, 196–207. [Google Scholar] [CrossRef]
- Morrisey, E.E.; Ip, H.S.; Lu, M.M.; Parmacek, M.S. GATA6: A zinc finger transcription factor that is expressed in multiple cell lineages derived from lateral mesoderm. Dev. Biol. 1996, 177, 309–322. [Google Scholar] [CrossRef]
- Suzuki, E.; Evans, T.; Lowry, J.; Truong, L.; Bell, D.W.; Testa, J.R.; Walsh, K. The human GATA-6 gene: Structure, chromosomal location, and regulation of expression by tissue-specific and mitogen-responsive signals. Genomics 1996, 38, 283–290. [Google Scholar] [CrossRef]
- Brewer, A.; Gove, C.; Davies, A.; McNulty, C.; Barrow, D.; Koutsourakis, M.; Farzaneh, F.; Pizzey, J.; Bomford, A.; Patient, R. The human and mouse GATA6 genes utilize two promoters and two initiation codons. J. Biol. Chem. 1999, 274, 38004–38016. [Google Scholar] [CrossRef]
- Laforest, B.; Nemer, M. GATA5 interacts with GATA4 and GATA6 in outflow tract development. Dev. Biol. 2011, 358, 368–378. [Google Scholar] [CrossRef]
- Morrisey, E.E.; Tang, Z.; Sigrist, K.; Lu, M.M.; Jiang, F.; Ip, H.S.; Parmacek, M.S. GATA6 regulates HNF4 and is required for differentiation of visceral endoderm in the mouse embryo. Genes Dev. 1998, 12, 3579–3590. [Google Scholar] [CrossRef]
- Xin, M.; Davis, C.A.; Molkentin, J.D.; Lien, C.L.; Duncan, S.A.; Richardson, J.A.; Olson, E.N. A threshold of GATA4 and GATA6 expression is required for cardiovascular development. Proc. Nat. Acad. Sci. USA 2006, 103, 11189–11194. [Google Scholar]
- Van Berlo, J.H.; Elrod, J.W.; van den Hoogenhof, M.M.; York, A.J.; Aronow, B.J.; Duncan, S.A.; Molkentin, J.D. The transcription factor GATA6 regulates pathological cardiac hypertrophy. Circ. Res. 2010, 107, 1032–1040. [Google Scholar] [CrossRef]
- Lepore, J.J.; Mericko, P.A.; Cheng, L.; Lu, M.M.; Morrisey, E.E.; Parmacek, M.S. GATA-6 regulates semaphorin 3C and is required in cardiac neural crest for cardiovascular morphogenesis. J. Clin. Investig. 2006, 116, 929–939. [Google Scholar]
- Turbendian, H.K.; Gordillo, M.; Tsai, S.Y.; Lu, J.; Kang, G.; Liu, T.C.; Tang, A.; Liu, S.; Fishman, G.I.; Evans, T. GATA factors efficiently direct cardiac fate from embryonic stem cells. Development 2013, 140, 1639–1644. [Google Scholar] [CrossRef]
- Huang, R.T.; Xue, S.; Xu, Y.J.; Yang, Y.Q. Somatic mutations in the GATA6 gene underlie sporadic tetralogy of Fallot. Int. J. Mol. Med. 2013, 31, 51–58. [Google Scholar]
- Kodo, K.; Nishizawa, T.; Furutani, M.; Arai, S.; Yamamura, E.; Joo, K.; Takahashi, T.; Matsuoka, R.; Yamagishi, H. GATA6 mutations cause human cardiac outflow tract defects by disrupting semaphorin-plexin signaling. Proc. Nat. Acad. Sci. USA 2009, 106, 13933–13938. [Google Scholar] [CrossRef]
- Kodo, K.; Nishizawa, T.; Furutani, M.; Arai, S.; Ishihara, K.; Oda, M.; Makino, S.; Fukuda, K.; Takahashi, T.; Matsuoka, R.; et al. Genetic analysis of essential cardiac transcription factors in 256 patients with non-syndromic congenital heart defects. Circ. J. 2012, 76, 1703–1711. [Google Scholar] [CrossRef]
- Lin, X.; Huo, Z.; Liu, X.; Zhang, Y.; Li, L.; Zhao, H.; Yan, B.; Liu, Y.; Yang, Y.; Chen, Y.H. A novel GATA6 mutation in patients with tetralogy of Fallot or atrial septal defect. J. Hum. Genet. 2010, 55, 662–667. [Google Scholar] [CrossRef]
- Maitra, M.; Koenig, S.N.; Srivastava, D.; Garg, V. Identification of GATA6 sequence variants in patients with congenital heart defects. Pediat. Res. 2010, 68, 281–285. [Google Scholar]
- Suzuki, S.; Nakao, A.; Sarhat, A.R.; Furuya, A.; Matsuo, K.; Tanahashi, Y.; Kajino, H.; Azuma, H. A case of pancreatic agenesis and congenital heart defects with a novel GATA6 nonsense mutation: Evidence of haploinsufficiency due to nonsense-mediated mRNA decay. Am. J. Med. Genet. A 2014, 164A, 476–479. [Google Scholar]
- Tomita-Mitchell, A.; Mahnke, D.K.; Struble, C.A.; Tuffnell, M.E.; Stamm, K.D.; Hidestrand, M.; Harris, S.E.; Goetsch, M.A.; Simpson, P.M.; Bick, D.P.; et al. Human gene copy number spectra analysis in congenital heart malformations. Physiol. Genomics 2012, 44, 518–541. [Google Scholar] [CrossRef]
- Wang, J.; Luo, X.J.; Xin, Y.F.; Liu, Y.; Liu, Z.M.; Wang, Q.; Li, R.G.; Fang, W.Y.; Wang, X.Z.; Yang, Y.Q. Novel GATA6 mutations associated with congenital ventricular septal defect or tetralogy of fallot. DNA Cell Biol. 2012, 31, 1610–1617. [Google Scholar]
- Yu, L.; Bennett, J.T.; Wynn, J.; Carvill, G.L.; Cheung, Y.H.; Shen, Y.; Mychaliska, G.B.; Azarow, K.S.; Crombleholme, T.M.; Chung, D.H.; et al. Whole exome sequencing identifies de novo mutations in GATA6 associated with congenital diaphragmatic hernia. J. Med. Genet. 2014, 51, 197–202. [Google Scholar]
- Zheng, G.F.; Wei, D.; Zhao, H.; Zhou, N.; Yang, Y.Q.; Liu, X.Y. A novel GATA6 mutation associated with congenital ventricular septal defect. Int. J. Mol. Med. 2012, 29, 1065–1071. [Google Scholar]
- Allen, H.L.; Flanagan, S.E.; Shaw-Smith, C.; de Franco, E.; Akerman, I.; Caswell, R.; International Pancreatic Agenesis Consortium; Ferrer, J.; Hattersley, A.T.; Ellard, S. GATA6 haploinsufficiency causes pancreatic agenesis in humans. Nat. Genet. 2011, 44, 20–22. [Google Scholar] [CrossRef]
- De Franco, E.; Shaw-Smith, C.; Flanagan, S.E.; Shepherd, M.H.; International Pancreatic Agenesis Consortium; Hattersley, A.T.; Ellard, S. GATA6 mutations cause a broad phenotypic spectrum of diabetes from pancreatic agenesis to adult-onset diabetes without exocrine insufficiency. Diabetes 2013, 62, 993–997. [Google Scholar]
- Yorifuji, T.; Kawakita, R.; Hosokawa, Y.; Fujimaru, R.; Yamaguchi, E.; Tamagawa, N. Dominantly inherited diabetes mellitus caused by GATA6 haploinsufficiency: Variable intrafamilial presentation. J. Med. Genet. 2012, 49, 642–643. [Google Scholar] [CrossRef]
- Van Berlo, J.H.; Aronow, B.J.; Molkentin, J.D. Parsing the roles of the transcription factors GATA4 and GATA6 in the adult cardiac hypertrophic response. PLoS One 2013, 8, e84591. [Google Scholar] [CrossRef]
- Heinemeyer, T.; Wingender, E.; Reuter, I.; Hermjakob, H.; Kel, A.E.; Kel, O.V.; Ignatieva, E.V.; Ananko, E.A.; Podkolodnaya, O.A.; Kolpakov, F.A.; et al. Databases on Transcriptional Regulation: TRANSFAC, TRRD, and COMPEL. Nucleic Acids Res. 1998, 26, 364–370. [Google Scholar]
- Lee, T.I.; Young, R.A. Transcriptional regulation and its misregulation in disease. Cell 2013, 152, 1237–1251. [Google Scholar] [CrossRef]
- Zaidi, S.; Choi, M.; Wakimoto, H.; Ma, L.; Jiang, J.; Overton, J.D.; Romano-Adesman, A.; Bjornson, R.D.; Breitbart, R.E.; Brown, K.K.; et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature 2013, 498, 220–223. [Google Scholar] [CrossRef]
- Cordell, H.J.; Bentham, J.; Topf, A.; Zelenika, D.; Heath, S.; Mamasoula, C.; Cosgrove, C.; Blue, G.; Granados-Riveron, J.; Setchfield, K.; et al. Genome-wide association study of multiple congenital heart disease phenotypes identifies a susceptibility locus for atrial septal defect at chromosome 4p16. Nat. Genet. 2013, 45, 822–824. [Google Scholar]
- Jimenez, P.; Saner, K.; Mayhew, B.; Rainey, W.E. GATA6 is expressed in the human adrenal and regulates transcription of genes required for adrenal androgen biosynthesis. Endocrinology 2003, 144, 4285–4288. [Google Scholar]
- Kamnasaran, D.; Guha, A. Expression of GATA6 in the human and mouse central nervous system. Brain Res. Dev. Brain Res. 2005, 160, 90–95. [Google Scholar] [CrossRef]
- Sun-Wada, G.H.; Manabe, S.; Yoshimizu, T.; Yamaguchi, C.; Oka, T.; Wada, Y.; Futai, M. Upstream regions directing heart-specific expression of the GATA6 gene during mouse early development. J. Biochem. 2000, 127, 703–709. [Google Scholar] [CrossRef]
- Molkentin, J.D.; Antos, C.; Mercer, B.; Taigen, T.; Miano, J.M.; Olson, E.N. Direct activation of a GATA6 cardiac enhancer by NKX2-5: Evidence for a reinforcing regulatory network of NKX2-5 and GATA transcription factors in the developing heart. Dev. Biol. 2000, 217, 301–309. [Google Scholar] [CrossRef]
- Fu, B.; Luo, M.; Lakkur, S.; Lucito, R.; Iacobuzio-Donahue, C.A. Frequent genomic copy number gain and over-expression of GATA6 in pancreatic carcinoma. Cancer Biol. Ther. 2008, 7, 1593–1601. [Google Scholar] [CrossRef]
- Ghatnekar, A.; Chrobak, I.; Reese, C.; Stawski, L.; Seta, F.; Wirrig, E.; Paez-Cortez, J.; Markiewicz, M.; Asano, Y.; Harley, R.; et al. Endothelial GATA6 deficiency promotes pulmonary arterial hypertension. Am. J. Pathol. 2013, 182, 2391–2406. [Google Scholar] [CrossRef]
- Ho, C.K.M.; Wood, J.R.; Stewart, D.R.; Ewens, K.; Ankener, W.; Wickenheisser, J.; Nelson-Degrave, V.; Zhang, Z.; Legro, R.S.; Dunaif, A.; et al. Increased transcription and increased messenger ribonucleic acid (mRNA) stability contribute to increased GATA6 mRNA abundance in polycystic ovary syndrome theca cells. J. Clin. Endocr. Metab. 2005, 90, 6596–6602. [Google Scholar] [CrossRef]
- Kamnasaran, D.; Qian, B.; Hawkins, C.; Stanford, W.L.; Guha, A. GATA6 is an astrocytoma tumor suppressor gene identified by gene trapping of mouse glioma model. Proc. Nat. Acad. Sci. USA 2007, 104, 8053–8058. [Google Scholar] [CrossRef]
- Vähätalo, R.; Asikainen, T.M.; Karikoski, R.; Kinnula, V.L.; White, C.W.; Andersson, S.; Heikinheimo, M.; Myllärniemi, M. Expression of transcription factor GATA6 in alveolar epithelial cells is linked to neonatal lung disease. Neonatology 2011, 99, 231–240. [Google Scholar] [CrossRef]
- Bruneau, B.G. Signaling and transcriptional networks in heart development and regeneration. Cold Spring Harb. Perspect. Biol. 2013, 5, a008292. [Google Scholar] [CrossRef]
- McCulley, D.J.; Black, B.L. Transcription factor pathways and congenital heart disease. Curr. Top. Dev. Biol. 2012, 100, 253–277. [Google Scholar] [CrossRef]
- Chen, J.; Wang, D.Z. MicroRNAs in cardiovascular development. J. Mol. Cell. Cardiol. 2012, 52, 949–957. [Google Scholar] [CrossRef]
- Srivastava, D. Making or breaking the heart: from lineage determination to morphogenesis. Cell 2006, 126, 1037–1048. [Google Scholar] [CrossRef]
- Charron, F.; Paradis, P.; Bronchain, O.; Nemer, G.; Nemer, M. Cooperative interaction between GATA4 and GATA6 regulates myocardial gene expression. Mol. Cell Biol. 1999, 19, 4355–4365. [Google Scholar]
- Zhao, R.; Watt, A.J.; Battle, M.A.; Li, J.; Bondow, B.J.; Duncan, S.A. Loss of both GATA4 and GATA6 blocks cardiac myocyte differentiation and results in acardia in mice. Dev. Biol. 2008, 317, 614–619. [Google Scholar] [CrossRef]
- Maitra, M.; Schluterman, M.K.; Nichols, H.A.; Richardson, J.A.; Lo, C.W.; Srivastava, D.; Garg, V. Interaction of GATA4 and GATA6 with TBX5 is critical for normal cardiac development. Dev. Biol. 2009, 326, 368–377. [Google Scholar] [CrossRef]
- Lien, C.L.; Wu, C.; Mercer, B.; Webb, R.; Richardson, J.A.; Olson, E.N. Control of early cardiac-specific transcription of NKX2-5 by a GATA-dependent enhancer. Development 1999, 126, 75–84. [Google Scholar]
- Searcy, R.D.; Vincent, E.B.; Liberatore, C.M.; Yutzey, K.E. A GATA-dependent NKX2-5 regulatory element activates early cardiac gene expression in transgenic mice. Development 1998, 125, 4461–4470. [Google Scholar]
- Fischer, A.; Klattig, J.; Kneitz, B.; Diez, H.; Maier, M.; Holtmann, B.; Englert, C.; Gessler, M. Hey basic helix-loop-helix transcription factors are repressors of GATA4 and GATA6 and restrict expression of the GATA target gene ANF in fetal hearts. Mol. Cell. Biol. 2005, 25, 8960–8970. [Google Scholar]
- Sakata, Y.; Kamei, C.N.; Nakagami, H.; Bronson, R.; Liao, J.K.; Chin, M.T. Ventricular septal defect and cardiomyopathy in mice lacking the transcription factor CHF1/Hey2. Proc. Natl. Acad. Sci. USA 2002, 99, 16197–16202. [Google Scholar]
- Van der Bom, T.; Zomer, A.C.; Zwinderman, A.H.; Meijboom, F.J.; Bouma, B.J.; Mulder, B.J. The changing epidemiology of congenital heart disease. Nat. Rev. Cardiol. 2011, 8, 50–60. [Google Scholar] [CrossRef]
- Verheugt, C.L.; Uiterwaal, C.S.; van der Velde, E.T.; Meijboom, F.J.; Pieper, P.G.; van Dijk, A.P.; Vliegen, H.W.; Grobbee, D.E.; Mulder, B.J. Mortality in adult congenital heart disease. Eur. Heart J. 2010, 31, 1220–1229. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, C.; Li, X.; Pang, S.; Chen, W.; Qin, X.; Huang, W.; Zeng, C.; Yan, B. Novel and Functional DNA Sequence Variants within the GATA6 Gene Promoter in Ventricular Septal Defects. Int. J. Mol. Sci. 2014, 15, 12677-12687. https://doi.org/10.3390/ijms150712677
Li C, Li X, Pang S, Chen W, Qin X, Huang W, Zeng C, Yan B. Novel and Functional DNA Sequence Variants within the GATA6 Gene Promoter in Ventricular Septal Defects. International Journal of Molecular Sciences. 2014; 15(7):12677-12687. https://doi.org/10.3390/ijms150712677
Chicago/Turabian StyleLi, Chunyu, Xianke Li, Shuchao Pang, Wei Chen, Xianyun Qin, Wenhui Huang, Changqing Zeng, and Bo Yan. 2014. "Novel and Functional DNA Sequence Variants within the GATA6 Gene Promoter in Ventricular Septal Defects" International Journal of Molecular Sciences 15, no. 7: 12677-12687. https://doi.org/10.3390/ijms150712677
APA StyleLi, C., Li, X., Pang, S., Chen, W., Qin, X., Huang, W., Zeng, C., & Yan, B. (2014). Novel and Functional DNA Sequence Variants within the GATA6 Gene Promoter in Ventricular Septal Defects. International Journal of Molecular Sciences, 15(7), 12677-12687. https://doi.org/10.3390/ijms150712677