Effect of ?-Glucan Supplementation on Acute Postprandial Changes in Fatty Acid Profile of Lymph and Serum in Pigs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
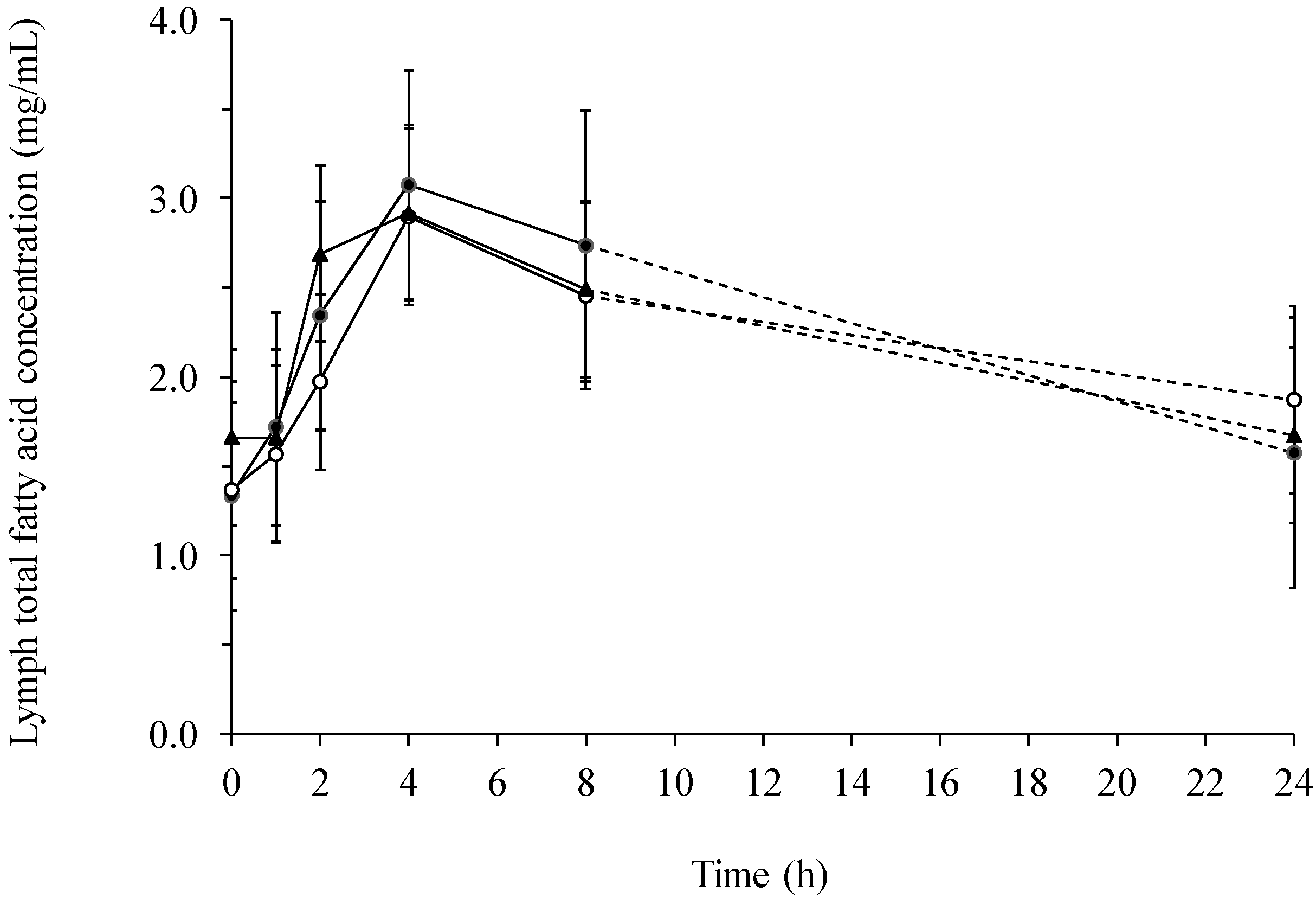
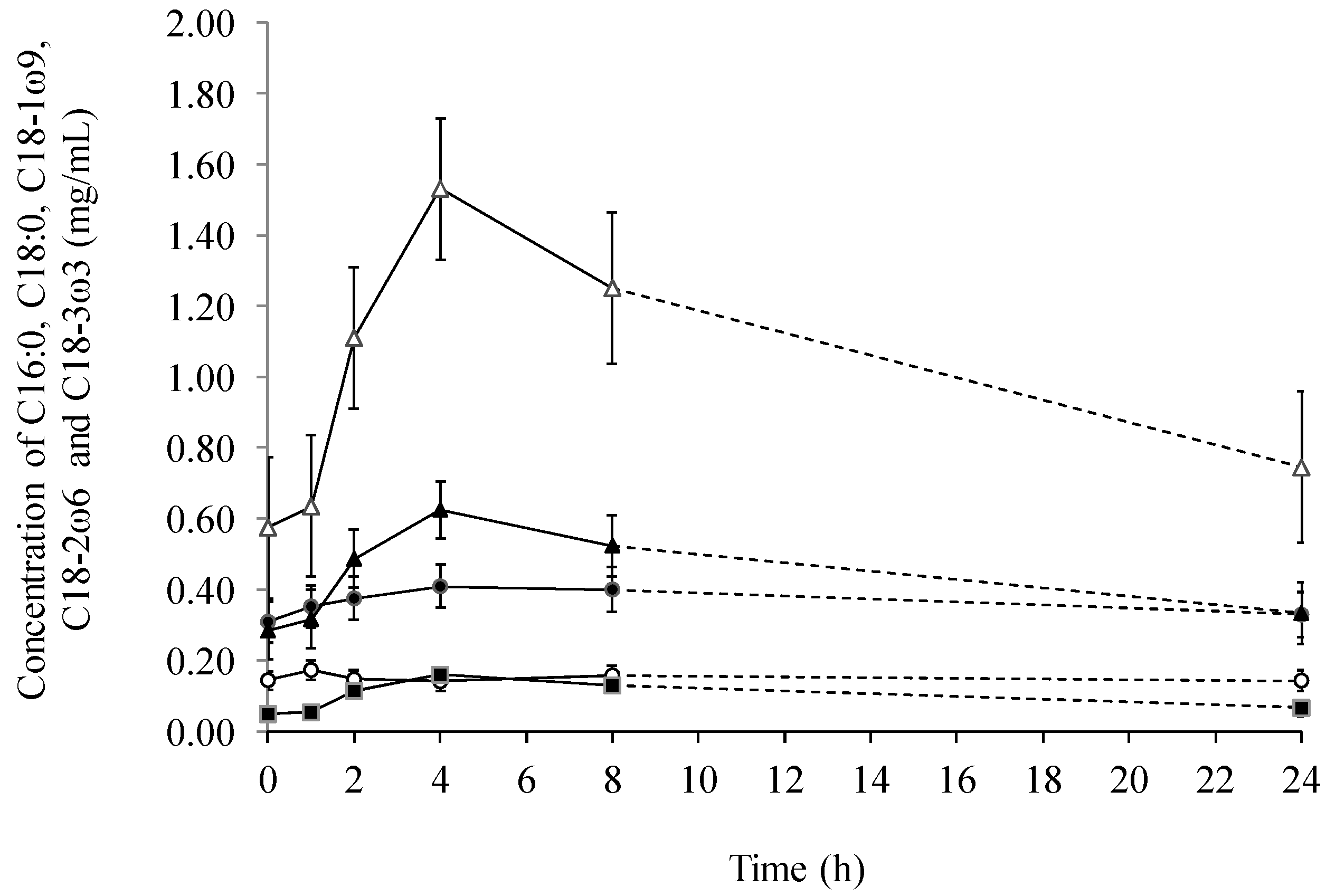
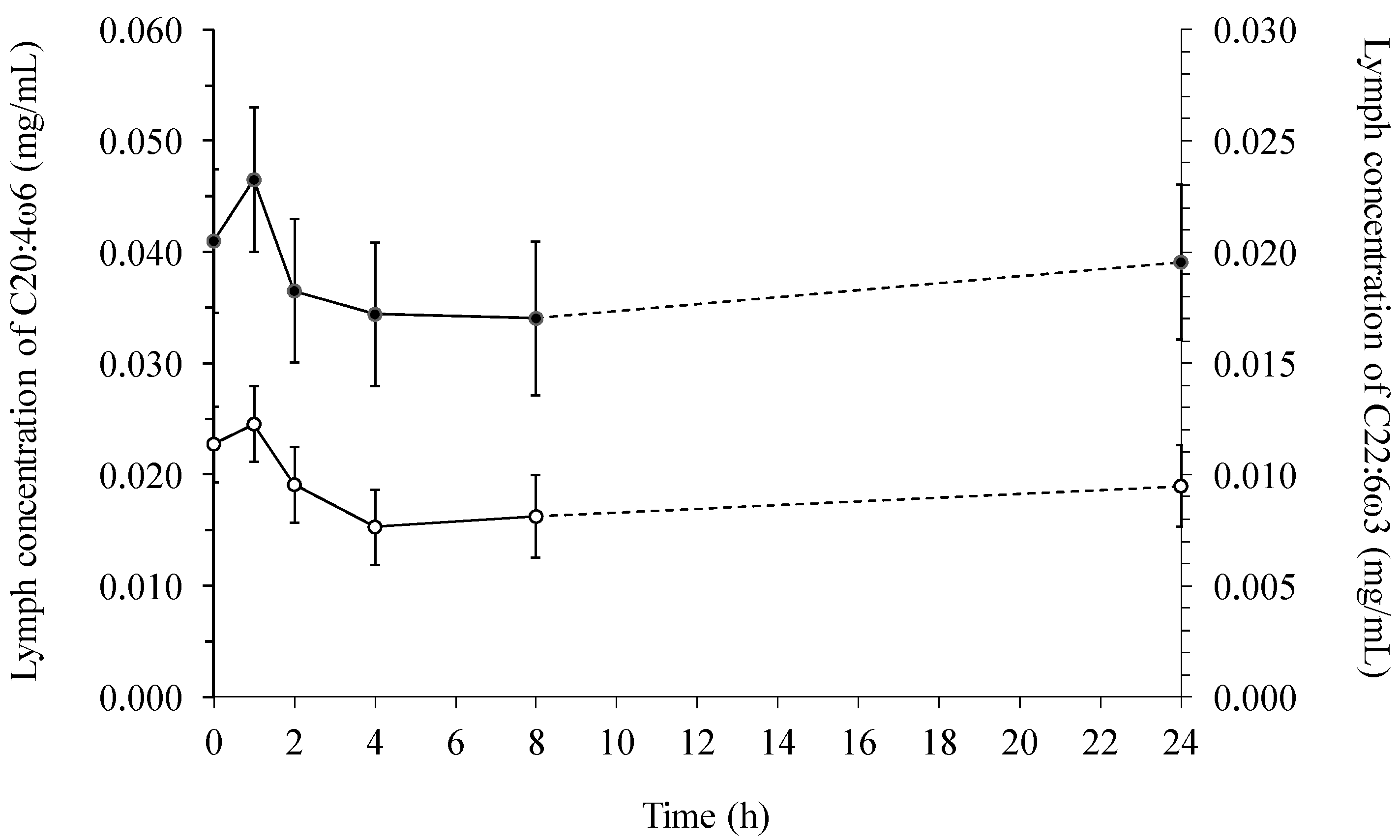
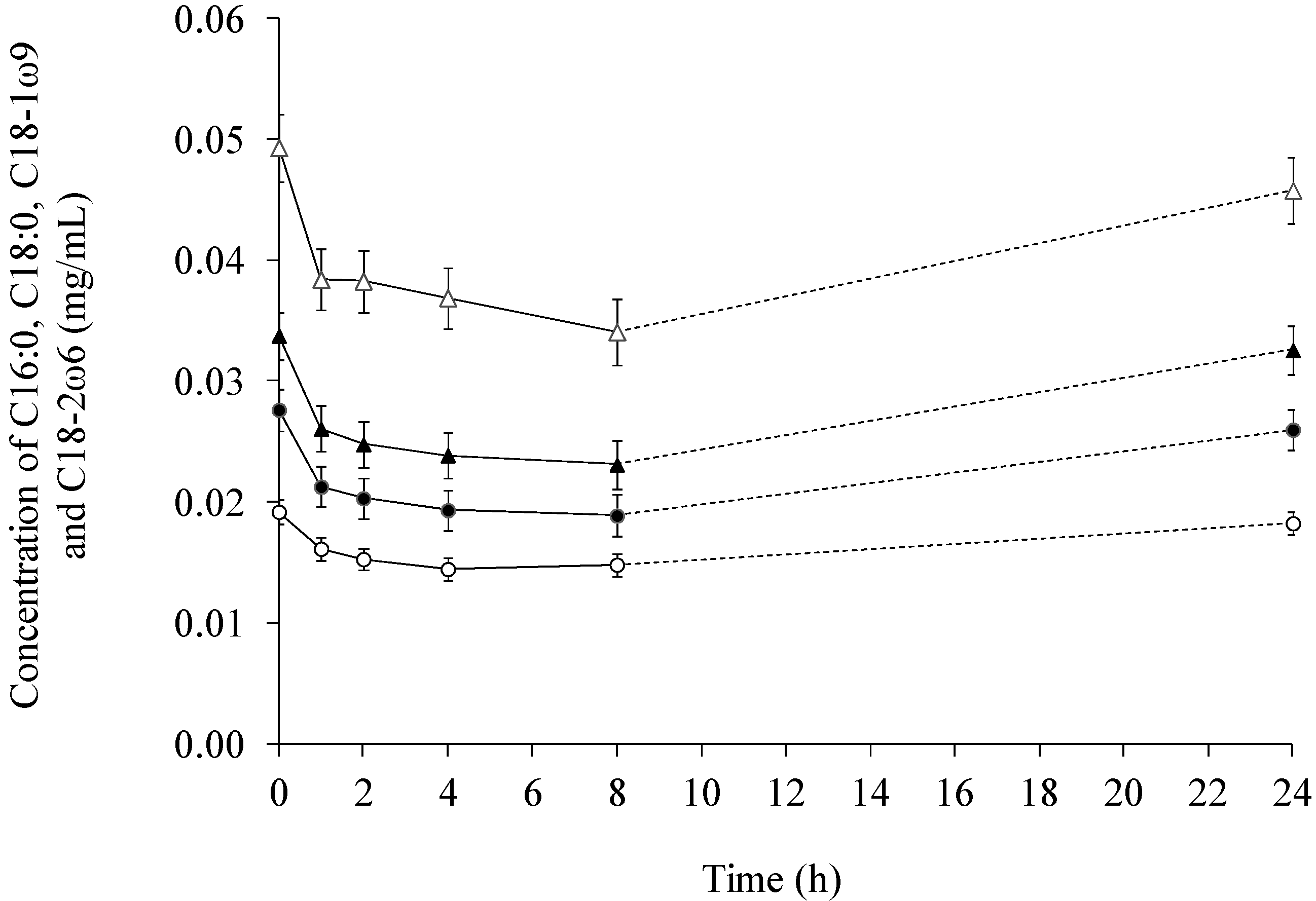
2.1. Fatty Acids in Lymph



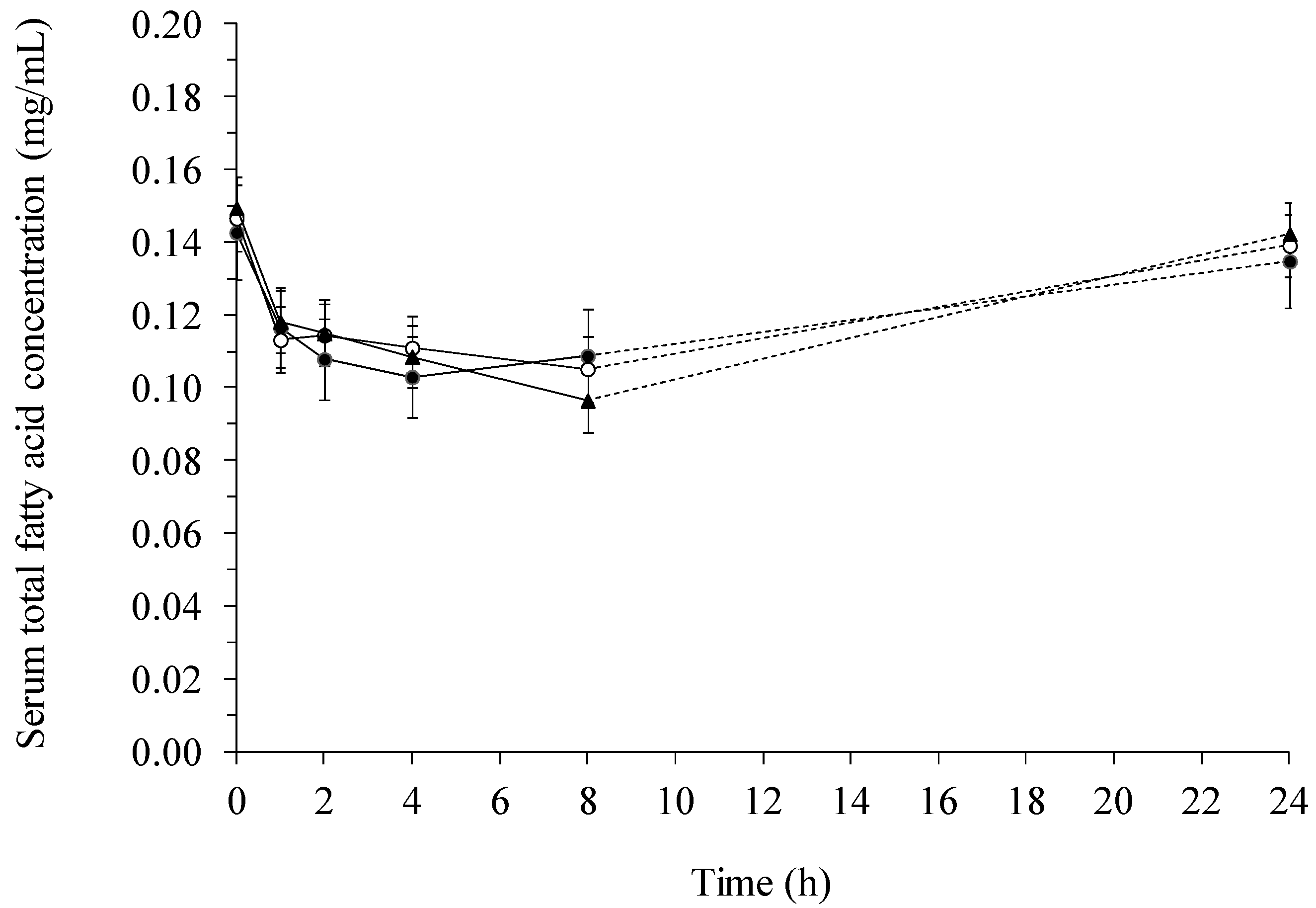
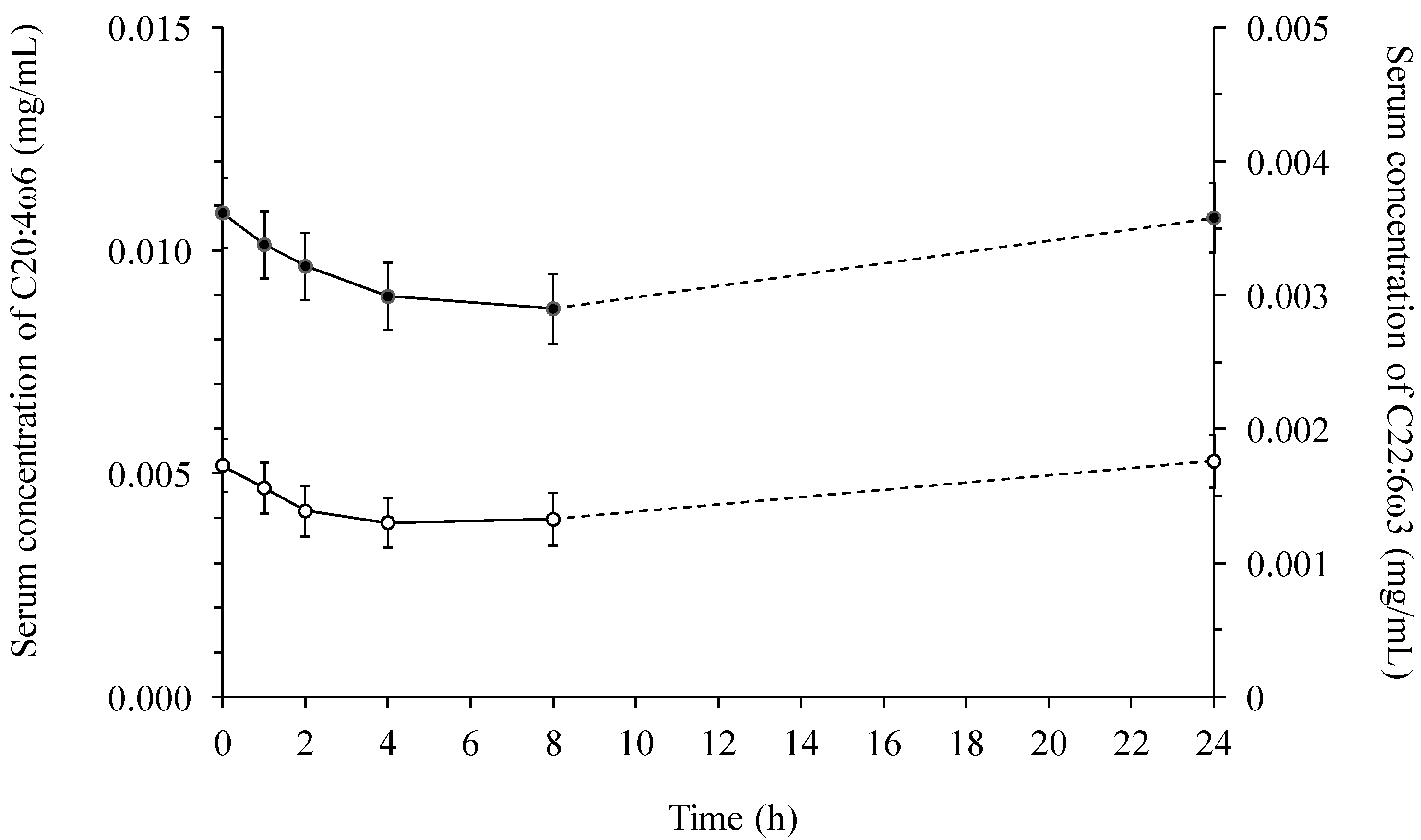
2.2. Fatty Acid Contents in Serum



3. Discussion
4. Experimental Section
4.1. Animals and Surgery
4.2. Feeding and Sampling
4.3. Chemical Analysis
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sniderman, A.D.; Williams, K.; Contois, J.H.; Monroe, H.M.; McQueen, M.J.; de Graaf, J.; Furberg, C.D. A meta-analysis of low-density lipoprotein cholesterol, non-high-density lipoprotein cholesterol, and apolipoprotein B as markers of cardiovascular risk. Circ. Cardiovasc. Qual. Outcomes 2011, 4, 337–345. [Google Scholar] [CrossRef]
- Brown, L.; Rosner, B.; Willett, W.W.; Sacks, F.M. Cholesterol-lowering effects of dietary fiber: A meta-analysis. Am. J. Clin. Nutr. 1999, 69, 30–42. [Google Scholar]
- Queenan, K.M.; Stewart, M.L.; Smith, K.N.; Thomas, W.; Fulcher, R.G.; Slavin, J.L. Concentrated oat β-glucan, a fermentable fiber, lowers serum cholesterol in hypercholesterolemic adults in a randomized controlled trial. Nutr. J. 2008, 6, 6. [Google Scholar] [CrossRef]
- Behall, K.M.; Scholfield, D.J.; Hallfrisch, J. Diets containing barley significantly reduce lipids in mildly hypercholesterolemic men and women. Am. J. Clin. Nutr. 2004, 80, 1185–1193. [Google Scholar]
- Wilson, T.A.; Nicolosi, R.J.; Delaney, B.; Chadwell, K.; Moolchandani, V.; Kotyla, T.; Ponduru, S.; Zheng, G.H.; Hess, R.; Knutson, N.; et al. Reduced and high molecular weight barley β-glucans decrease plasma total and non-HDL-cholesterol in hypercholesterolemic Syrian golden hamsters. J. Nutr. 2004, 134, 2617–2622. [Google Scholar]
- Bourdon, I.; Yokoyama, W.; Davis, P.; Hudson, C.; Backus, R.; Richter, D.; Knuckles, B.; Schneeman, B.O. Postprandial lipid, glucose, insulin, and cholecystokinin responses in men fed barley pasta enriched with beta-glucan. Am. J. Clin. Nutr. 1999, 69, 55–63. [Google Scholar]
- Talati, R.; Baker, W.L.; Pabilonia, M.S.; White, C.M.; Coleman, C.I. The effects of barley-derived soluble fiber on serum lipids. Ann. Fam. Med. 2009, 7, 157–163. [Google Scholar]
- Bell, S.; Goldman, V.M.; Bistrian, B.R.; Arnold, A.H.; Ostroff, G.; Forse, R.A. Effect of β-glucan from oats and yeast on serum lipids. Crit. Rev. Food Sci. Nutr. 1999, 39, 189–202. [Google Scholar] [CrossRef]
- Bartnikowska, E.; Zabielski, R. Dynamics of absorption of dietary cholesterol from the alimentary tract as influenced by pectin in pigs. J. Anim. Physiol. Anim. Nutr. 1990, 64, 174–179. [Google Scholar] [CrossRef]
- Larsen, F.H.; Jorgensen, H.; Engelsen, S.B.; Laerke, H.N. Metabolic profiling of lymph from pigs fed with β-glucan by high-resolution (1)H NMR spectroscopy. Livest. Sci. 2010, 133, 38–41. [Google Scholar]
- Knothe, G.; Kenar, J.A. Determination of the fatty acid profile by H-1-NMR spectroscopy. Eur. J. Lipid Sci. Technol. 2004, 106, 88–96. [Google Scholar] [CrossRef]
- Casiraghi, M.C.; Garsetti, M.; Testolin, G.; Brighenti, F. Post-prandial responses to cereal products enriched with barley β-glucan. J. Am. Coll. Nutr. 2006, 25, 313–320. [Google Scholar] [CrossRef]
- Ikeda, I.; Tomari, Y.; Sugano, M. Interrelated effects of dietary fiber and fat on lymphatic cholesterol and triglyceride absorption in rats. J. Nutr. 1989, 119, 1383–1387. [Google Scholar]
- Drozdowski, L.A.; Reimer, R.A.; Temelli, F.; Bell, R.C.; Vasanthan, T.; Thomson, A.B.R. β-Glucan extracts inhibit the in vitro intestinal uptake of long-chain fatty acids and cholesterol and down-regulate genes involved in lipogenesis and lipid transport in rats. J. Nutr. Biochem. 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Zabielski, R.; Pierzynowski, S.G. A model for long-term sampling of lymph from the jejunal lymphatic trunk in pigs and sheep. J. Anim. Physiol. Anim. Nutr. 1992, 68, 146–150. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar]
- Jensen, S.K. Improved Bligh and Dyer extraction procedure. Lipid Technol. 2008, 20, 280–281. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lærke, H.N.; Mikkelsen, L.S.; Jørgensen, H.; Jensen, S.K. Effect of ?-Glucan Supplementation on Acute Postprandial Changes in Fatty Acid Profile of Lymph and Serum in Pigs. Int. J. Mol. Sci. 2014, 15, 13881-13891. https://doi.org/10.3390/ijms150813881
Lærke HN, Mikkelsen LS, Jørgensen H, Jensen SK. Effect of ?-Glucan Supplementation on Acute Postprandial Changes in Fatty Acid Profile of Lymph and Serum in Pigs. International Journal of Molecular Sciences. 2014; 15(8):13881-13891. https://doi.org/10.3390/ijms150813881
Chicago/Turabian StyleLærke, Helle Nygaard, Lasse Sommer Mikkelsen, Henry Jørgensen, and Søren Krogh Jensen. 2014. "Effect of ?-Glucan Supplementation on Acute Postprandial Changes in Fatty Acid Profile of Lymph and Serum in Pigs" International Journal of Molecular Sciences 15, no. 8: 13881-13891. https://doi.org/10.3390/ijms150813881
APA StyleLærke, H. N., Mikkelsen, L. S., Jørgensen, H., & Jensen, S. K. (2014). Effect of ?-Glucan Supplementation on Acute Postprandial Changes in Fatty Acid Profile of Lymph and Serum in Pigs. International Journal of Molecular Sciences, 15(8), 13881-13891. https://doi.org/10.3390/ijms150813881