Association between Phosphatase Related Gene Variants and Coronary Artery Disease: Case-Control Study and Meta-Analysis

Abstract
:1. Introduction
2. Results
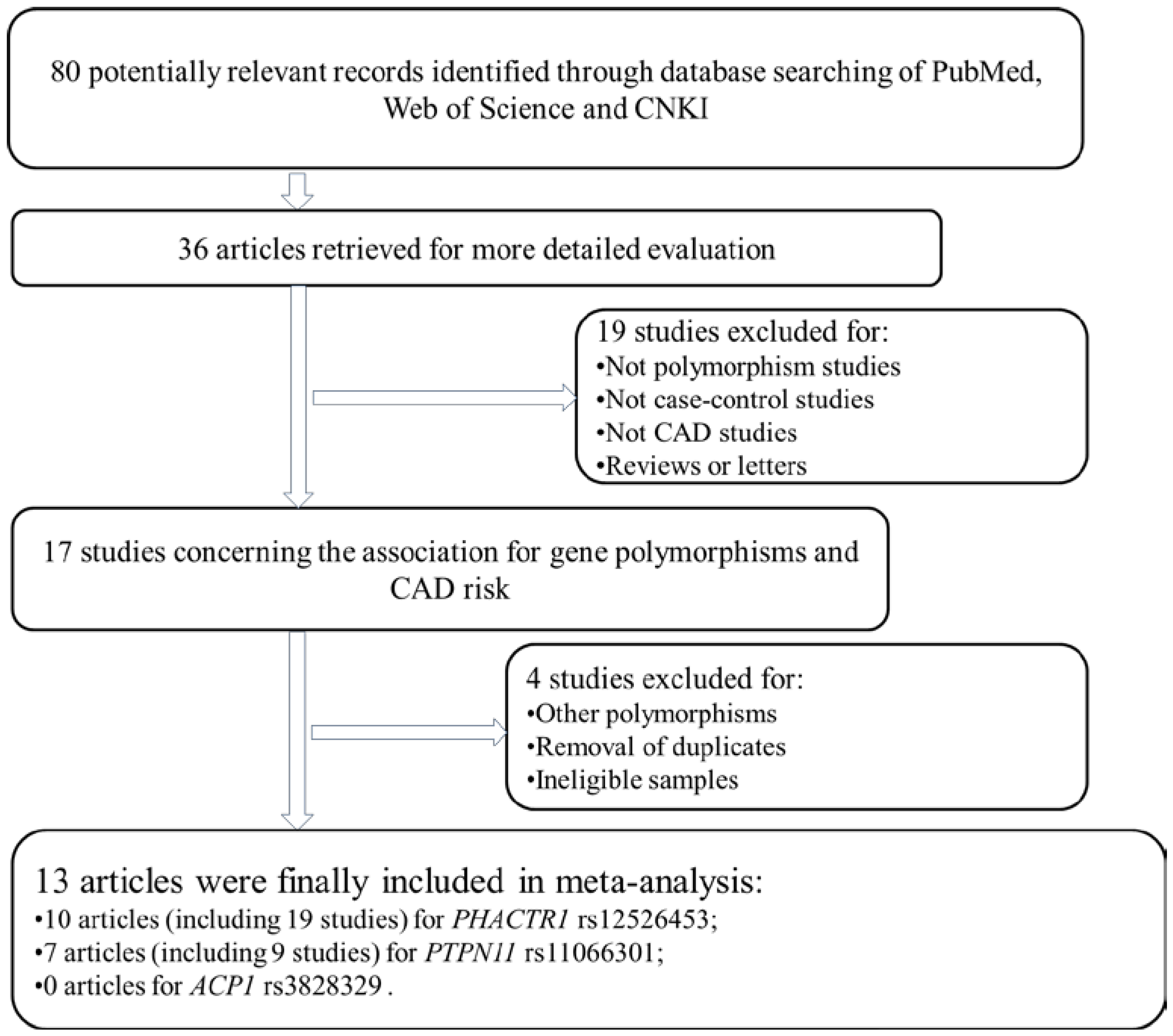
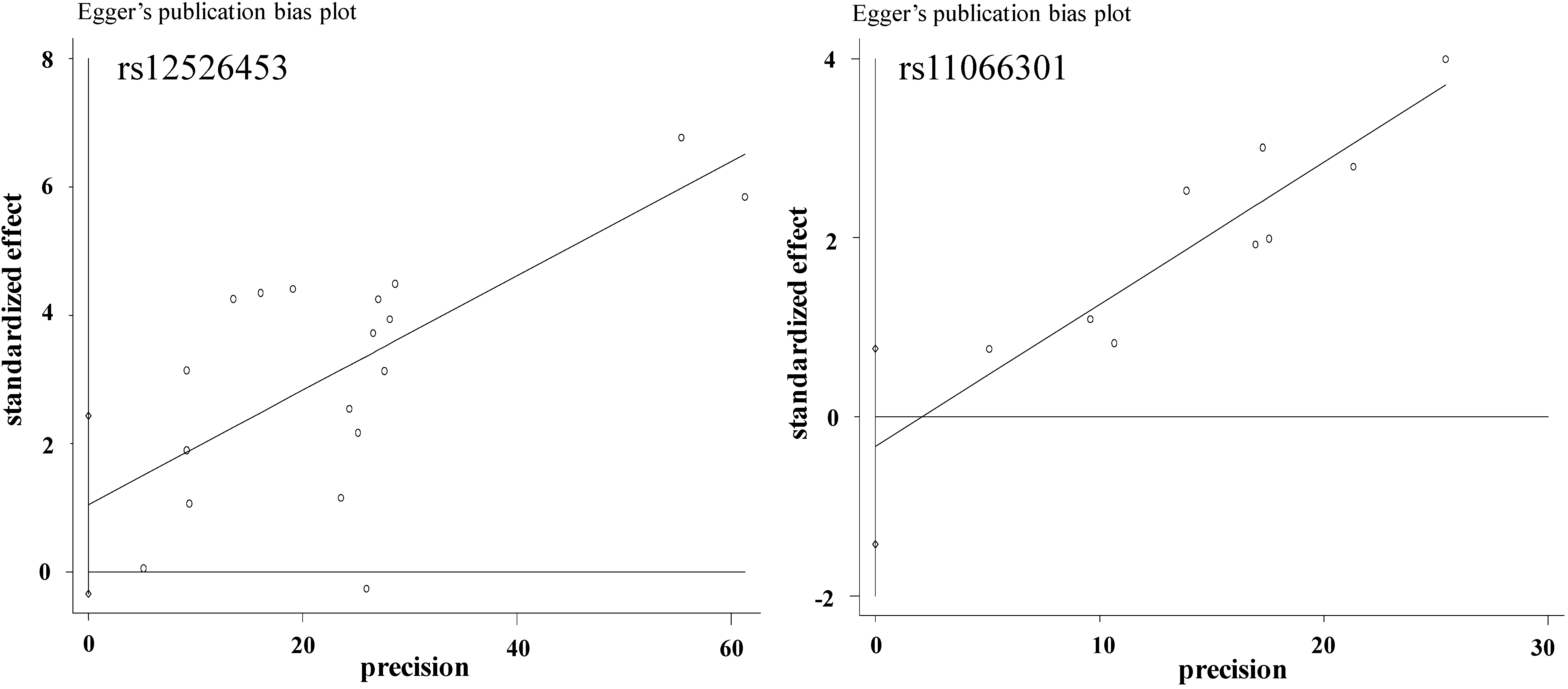
2.1. Literature Analysis

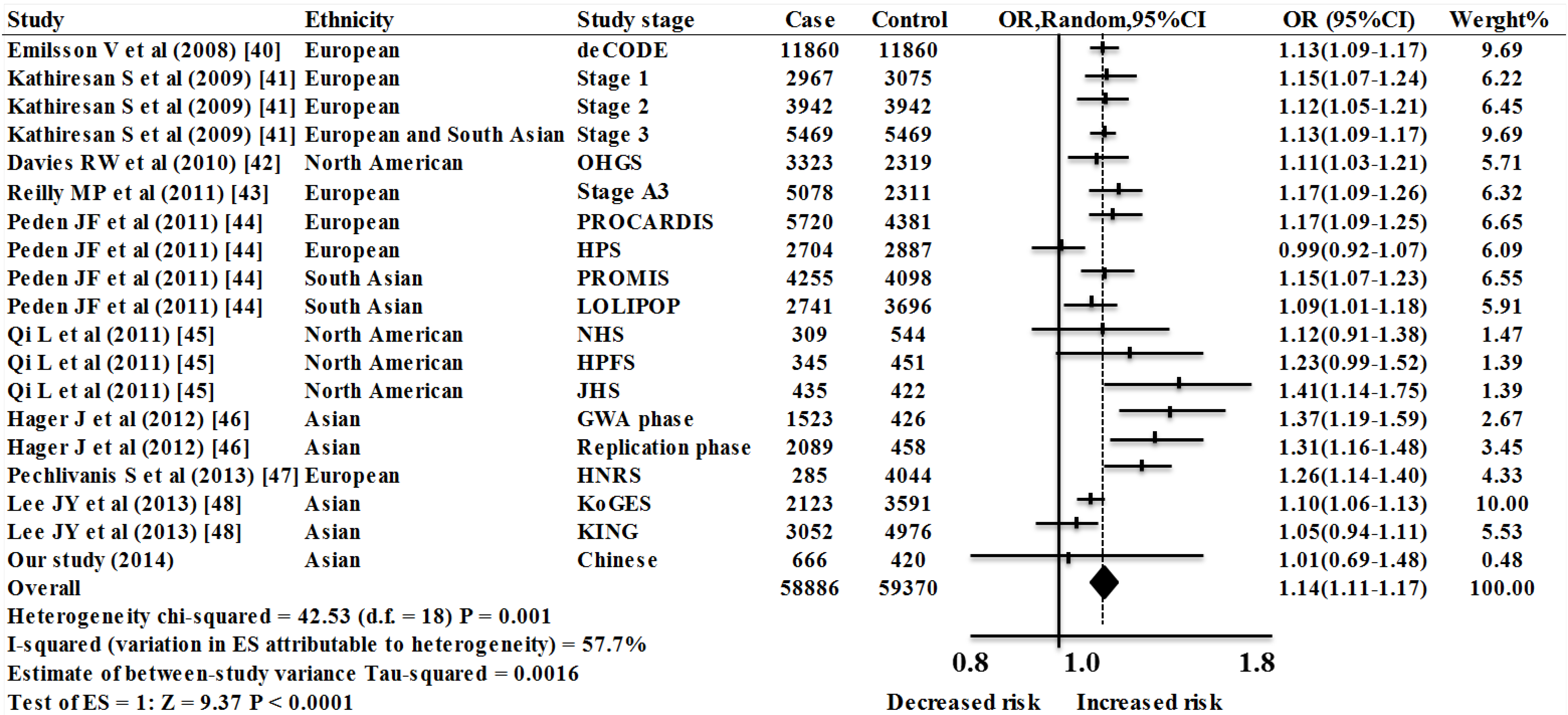
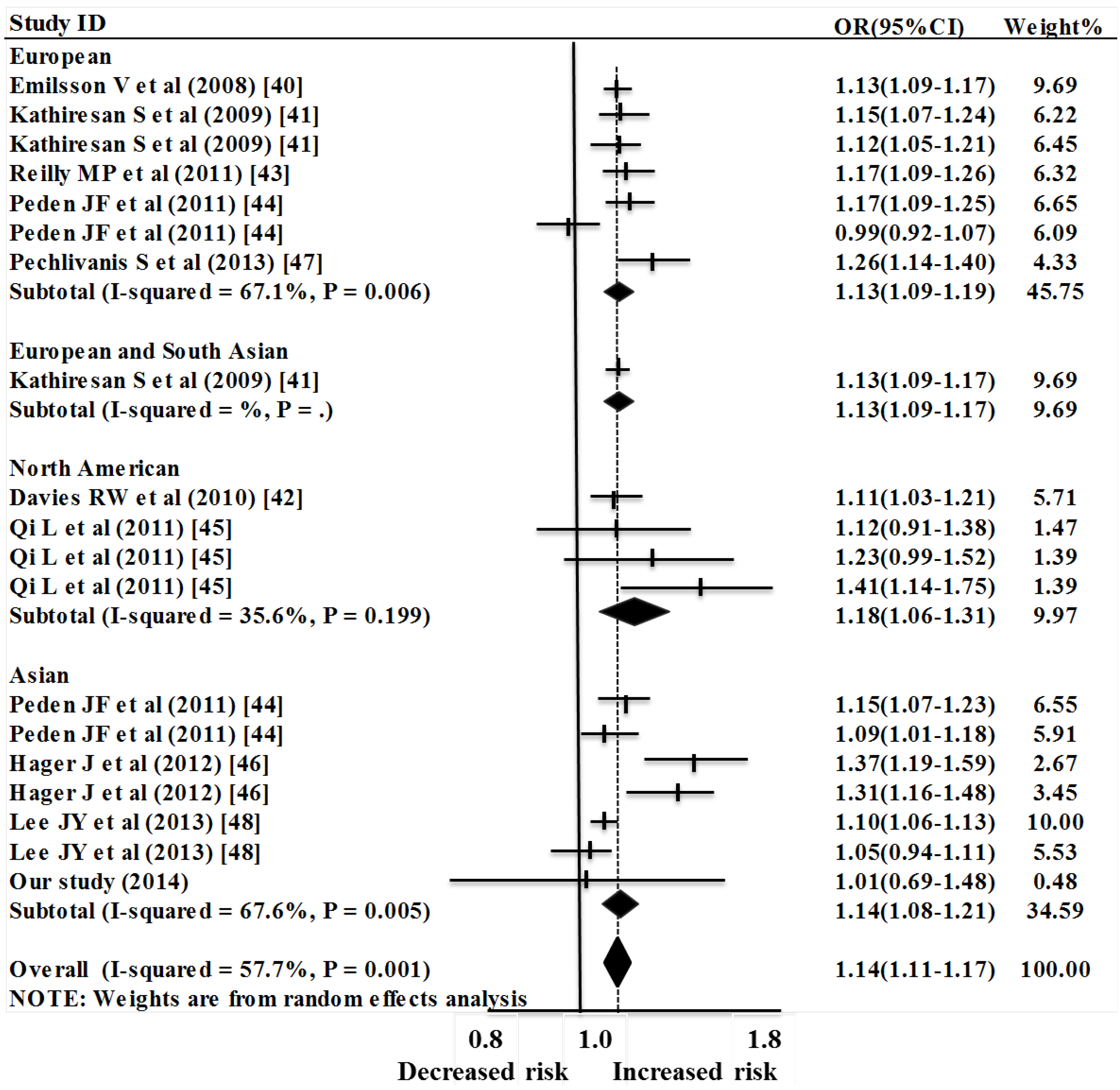
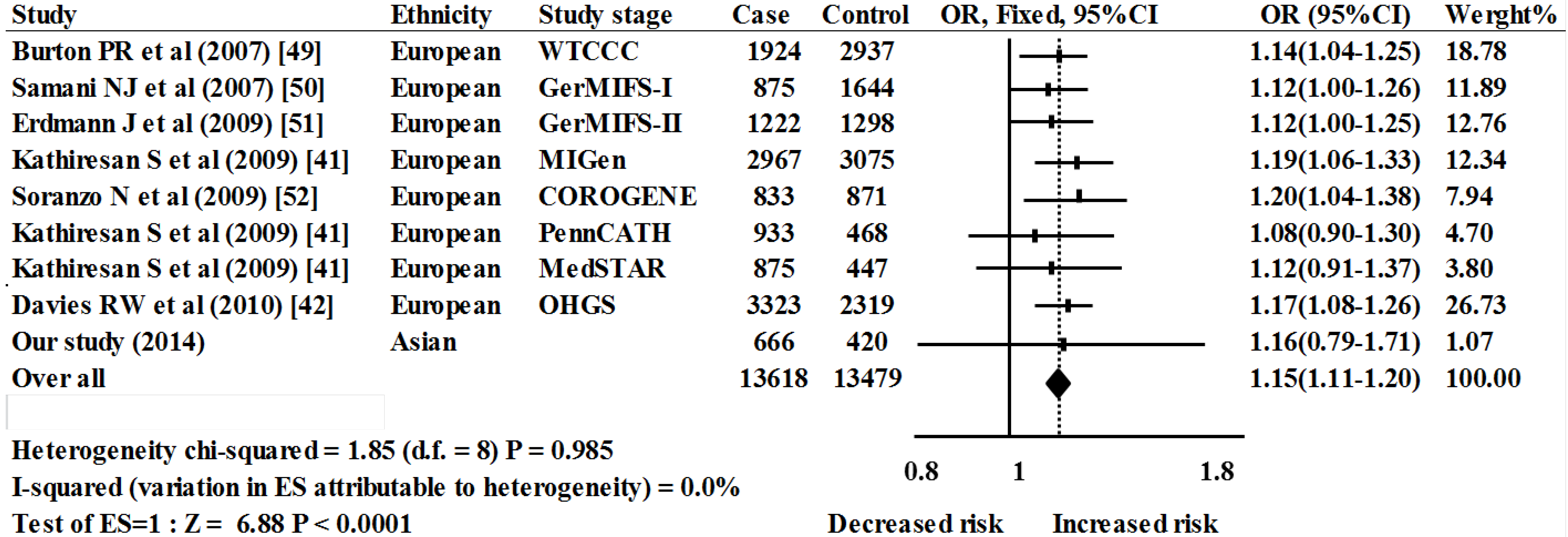
2.2. Meta-Analysis Results





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP/Group | Genotype | χ2 | P (df = 2) | Allele | χ2 | P (df = 1) | HWE | OR (95% CI) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| rs12526453 | CC (%) | CG (%) | GG (%) | C (%) | G (%) | ||||||
| case | 594 (89.2) | 72 (10.8) | 0 (0.0) | 1260 (94.6) | 72 (5.4) | 0.25 | |||||
| control | 375 (89.3) | 45 (10.7) | 0 (0.0) | 0.00 | 1.00 | 795 (94.6) | 45 (5.4) | 0.00 | 1.00 | 0.62 | 1.01 (0.69–1.48) |
| rs11066301 | AA (%) | AG (%) | GG (%) | A (%) | G (%) | ||||||
| case | 606 (91.0) | 54 (8.1) | 6 (0.9) | 1266 (95.0) | 66 (5.0) | 0.003 | |||||
| control | 378 (90.0) | 36 (8.6) | 6 (1.4) | 0.74 | 0.69 | 792 (94.3) | 48 (5.7) | 0.60 | 0.44 | 0.06 | 1.16 (0.79–1.71) |
| rs3828329 | CC (%) | CT (%) | TT (%) | C (%) | T (%) | ||||||
| case | 360 (54.1) | 270 (40.5) | 36 (5.4) | 990 (74.3) | 342 (25.7) | 0.13 | |||||
| control | 270 (64.3) | 138 (32.9) | 12 (2.8) | 12.48 | 0.002 | 678 (80.7) | 162 (19.3) | 11.80 | 0.0006 | 0.34 | 1.45 (1.17–1.78) |
| SNP/Group | Dominant | χ2 | P (df = 1) | OR (95% CI) | Recessive | χ2 | P (df = 1) | OR (95% CI) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| rs12526453 | CC | CG + GG | CC + CG | GG | ||||||
| case | 594 | 72 | 666 | 0 | ||||||
| control | 375 | 45 | 0.00 | 1.00 | 1.01 (0.69–1.48) | 420 | 0 | NA | NA | NA |
| rs11066301 | AA | AG + GG | AA + AG | GG | ||||||
| case | 606 | 60 | 660 | 6 | ||||||
| control | 378 | 42 | 0.30 | 0.58 | 0.89 (059–1.39) | 414 | 6 | NA | NA | 0.63 (0.20–1.96) |
| rs3828329 | CC | CT + TT | CC + CT | TT | ||||||
| case | 360 | 306 | 630 | 36 | ||||||
| control | 270 | 150 | 11.07 | 0.0009 | 1.53 (1.19–1.97) | 408 | 12 | 3.96 | 0.047 | 1.94 (1.00–3.78) |
| Gender | Group | Genotype(n) | Allele(n) | Additive | Dominant | Recessive | |||
|---|---|---|---|---|---|---|---|---|---|
| CC/CT/TT | C/T | P (df = 1) | OR (95% CI) | P (df = 1) | OR (95% CI) | P (df = 1) | OR (95% CI) | ||
| male | cases (N = 400) | 242/172/20 | 656/212 | 0.109 | 1.25 (0.95–1.65) | 0.120 | 1.29 (0.93–1.80) | 0.337 | 1.53 (0.64–3.68) |
| controls (N = 252) | 142/80/7 | 364/94 | |||||||
| female | cases (N = 261) | 118/98/16 | 334/130 | 0.001 | 1.80 (1.29–2.50) | 0.0008 | 1.96 (1.32–2.92) | 0.044 | 2.76 (1.00–7.66) |
| controls (N = 177) | 128/58/5 | 314/68 | |||||||
| Gender | Age | Group | Genotype(n) | X2 | P (df = 2) | Allele(n) | X2 | P (df = 1) | OR (95% CI) |
|---|---|---|---|---|---|---|---|---|---|
| CC/CT/TT | C/T | ||||||||
| All | <65 | cases (N = 338) | 177/148/13 | 502/174 | |||||
| controls (N = 247) | 154/88/5 | 6.41 | 0.041 | 396/98 | 5.57 | 0.018 | 1.40 (1.06–1.85) | ||
| ≥65 | cases (N = 328) | 183/122/23 | 488/168 | ||||||
| controls (N = 173) | 116/50/7 | 6.34 | 0.042 | 282/64 | 6.44 | 0.011 | 1.52 (1.10–2.10) | ||
| male | <65 | cases (N = 202) | 104/89/9 | 297/107 | |||||
| controls (N = 136) | 84/49/3 | 3.99 | 0.136 | 217/55 | 3.50 | 0.061 | 1.42 (0.98–2.05) | ||
| ≥65 | cases (N = 232) | 138/83/11 | 359/105 | ||||||
| controls (N = 91) | 58/31/2 | 1.30 | 0.522 | 147/35 | 0.89 | 0.345 | 1.23 (0.80–1.88) | ||
| female | <65 | cases (N = 136) | 73/59/4 | 205/67 | |||||
| controls (N = 111) | 70/39/2 | 2.30 | 0.316 | 179/43 | 1.96 | 0.161 | 1.36 (0.88–2.10) | ||
| ≥65 | cases (N = 96) | 45/39/12 | 129/63 | ||||||
| controls (N = 80) | 58/19/3 | 12.58 | 0.002 | 135/25 | 10.56 | 0.001 | 2.27 (1.37–3.75) |
2.3. Case-Control Study
3. Discussion
4. Experimental Section
4.1. Literature Review and Data Extraction
| Study Population | Studies, n | OR (95% CI) | Weight % | Z | P(z) | I2 | p | τ2 | Heterogeneity Statistic |
|---|---|---|---|---|---|---|---|---|---|
| European | 7 | 1.14 (1.09–1.19) | 45.75 | 5.60 | <0.0001 | 67.1% | 0.006 | 0.0023 | 18.24 |
| North American | 4 | 1.18 (1.06–1.31) | 9.97 | 3.14 | 0.002 | 35.6% | 0.199 | 0.0041 | 4.66 |
| Asian | 7 | 1.14 (1.08–1.21) | 34.59 | 4.58 | <0.0001 | 67.6% | 0.005 | 0.0034 | 18.52 |
| European and South Asian | 1 | 1.13 (1.09–1.17) | 9.69 | 6.76 | <0.0001 | NA | NA | 0.00 | 0.00 |
| Overall | 19 | 1.14 (1.11–1.17) | 100.00 | 9.37 | <0.0001 | 57.7% | 0.001 | 0.0016 | 42.53 |
4.2. Case-Control Study Sample Collection
4.3. SNP Genotyping
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Zhang, X.H.; Lu, Z.L.; Liu, L. Coronary heart disease in China. Heart 2008, 94, 1126–1131. [Google Scholar]
- Visvikis-Siest, S.; Marteau, J.B. Genetic variants predisposing to cardiovascular disease. Curr. Opin. Lipidol. 2006, 17, 139–151. [Google Scholar]
- Roberts, R.; Stewart, A.F. 9p21 and the genetic revolution for coronary artery disease. Clin. Chem. 2012, 58, 104–112. [Google Scholar]
- Roberts, R.; Chen, L.; Wells, G.A.; Stewart, A.F. Recent success in the discovery of coronary artery disease genes. Can. J. Physiol. Pharmacol. 2011, 89, 609–615. [Google Scholar]
- Ren, H.; Panchatcharam, M.; Mueller, P.; Escalante-Alcalde, D.; Morris, A.J.; Smyth, S.S. Lipid phosphate phosphatase (LPP3) and vascular development. Biochim. Biophys. Acta 2013, 1831, 126–132. [Google Scholar]
- Schunkert, H.; Konig, I.R.; Kathiresan, S.; Reilly, M.P.; Assimes, T.L.; Holm, H.; Preuss, M.; Stewart, A.F.; Barbalic, M.; Gieger, C.; et al. Large-scale association analysis identifies 13 new susceptibility loci for coronary artery disease. Nat. Genet. 2011, 43, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Teruel, M.; Martin, J.E.; Gonzalez-Juanatey, C.; Lopez-Mejias, R.; Miranda-Filloy, J.A.; Blanco, R.; Balsa, A.; Pascual-Salcedo, D.; Rodriguez-Rodriguez, L.; Fernández-Gutierrez, B.; et al. Association of acid phosphatase locus 1*C allele with the risk of cardiovascular events in rheumatoid arthritis patients. Arthritis Res. Ther. 2011, 13, R116. [Google Scholar] [CrossRef] [Green Version]
- Mount, P.F.; Kemp, B.E.; Power, D.A. Regulation of endothelial and myocardial NO synthesis by multi-site eNOS phosphorylation. J. Mol. Cell. Cardiol. 2007, 42, 271–279. [Google Scholar]
- Dudzinski, D.M.; Michel, T. Life history of eNOS: Partners and pathways. Cardiovasc. Res. 2007, 75, 247–260. [Google Scholar]
- Hager, J.; Kamatani, Y.; Cazier, J.B.; Youhanna, S.; Ghassibe-Sabbagh, M.; Platt, D.E.; Abchee, A.B.; Romanos, J.; Khazen, G.; Othman, R.; et al. Genome-wide association study in a Lebanese cohort confirms PHACTR1 as a major determinant of coronary artery stenosis. PLoS One 2012, 7, e38663. [Google Scholar]
- Elchebly, M.; Payette, P.; Michaliszyn, E.; Cromlish, W.; Collins, S.; Loy, A.L.; Normandin, D.; Cheng, A.; Himms-Hagen, J.; Chan, C.C.; et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science 1999, 283, 1544–1548. [Google Scholar] [CrossRef]
- Raugei, G.; Ramponi, G.; Chiarugi, P. Low molecular weight protein tyrosine phosphatases: Small, but smart. Cell. Mol. Life Sci. 2002, 59, 941–949. [Google Scholar]
- Gloria-Bottini, F.; Bottini, N.; Renzetti, G.; Bottini, E. ACP1 and Th class of immunological disease: Evidence of interaction with gender. Int. Arch. Allergy Immunol. 2007, 143, 170–176. [Google Scholar]
- Alho, I.; Clara Bicho, M.; Carvalho, R.; da Silva, A.P.; Costa, L.; Bicho, M. Low molecular weight protein tyrosine phosphatase genetic polymorphism and susceptibility to cancer development. Cancer Genet. Cytogenet. 2008, 181, 20–24. [Google Scholar]
- Gloria-Bottini, F.; Banci, M.; Saccucci, P.; Neri, A.; Bottini, E.; Magrini, A. p53 codon 72 polymorphism and coronary artery disease: Evidence of interaction with ACP(1). Med. Sci. Monit. 2012, 18, CR712–CR715. [Google Scholar]
- Banci, M.; Saccucci, P.; D’Annibale, F.; Dofcaci, A.; Trionfera, G.; Magrini, A.; Bottini, N.; Bottini, E.; Gloria-Bottini, F. ACP1 genetic polymorphism and coronary artery disease: An association study. Cardiology 2009, 113, 236–242. [Google Scholar]
- Hirano, T. Interleukin 6 in autoimmune and inflammatory diseases: A personal memoir. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 717–730. [Google Scholar]
- Timmerman, I.; Hoogenboezem, M.; Bennett, A.M.; Geerts, D.; Hordijk, P.L.; van Buul, J.D. The tyrosine phosphatase SHP2 regulates recovery of endothelial adherens junctions through control of beta-catenin phosphorylation. Mol. Biol. Cell 2012, 23, 4212–4225. [Google Scholar]
- Mannell, H.; Krotz, F. SHP-2 regulates growth factor dependent vascular signalling and function. Mini Rev. Med. Chem. 2014, 14, 471–483. [Google Scholar]
- Seki, N.; Hashimoto, N.; Suzuki, Y.; Mori, S.; Amano, K.; Saito, Y. Role of SRC homology 2-containing tyrosine phosphatase 2 on proliferation of rat smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1081–1085. [Google Scholar]
- Won, K.J.; Lee, H.M.; Lee, C.K.; Lin, H.Y.; Na, H.; Lim, K.W.; Roh, H.Y.; Sim, S.; Song, H.; Choi, W.S.; et al. Protein tyrosine phosphatase SHP-2 is positively involved in platelet-derived growth factor-signaling in vascular neointima formation via the reactive oxygen species-related pathway. J. Pharmacol. Sci. 2011, 115, 164–175. [Google Scholar]
- Sturla, L.M.; Zinn, P.O.; Ng, K.; Nitta, M.; Kozono, D.; Chen, C.C.; Kasper, E.M. Src homology domain-containing phosphatase 2 suppresses cellular senescence in glioblastoma. Br. J. Cancer 2011, 105, 1235–1243. [Google Scholar]
- Mali, R.S.; Chan, R.; Kapur, R. Targeting SHP2 phosphatase in myeloproliferative neoplasms. Oncotarget 2012, 3, 1049–1051. [Google Scholar]
- Jiang, J.; Jia, Z.F.; Kong, F.; Jin, M.S.; Wang, Y.P.; Tian, S.; Suo, J.; Cao, X. Association of polymorphism of PTPN 11 encoding SHP-2 with gastric atrophy but not gastric cancer in Helicobacter pylori seropositive Chinese population. BMC Gastroenterol. 2012, 12, 89. [Google Scholar] [CrossRef]
- Dong, S.; Li, F.Q.; Zhang, Q.; Lv, K.Z.; Yang, H.L.; Gao, Y.; Yu, J.R. Expression and clinical significance of SHP2 in gastric cancer. J. Int. Med. Res. 2012, 40, 2083–2089. [Google Scholar]
- Dunlap, J.; Beadling, C.; Warrick, A.; Neff, T.; Fleming, W.H.; Loriaux, M.; Heinrich, M.C.; Kovacsovics, T.; Kelemen, K.; Leeborg, N.; et al. Multiplex high-throughput gene mutation analysis in acute myeloid leukemia. Hum. Pathol. 2012, 43, 2167–2176. [Google Scholar]
- Aceto, N.; Bentires-Alj, M. Targeting protein-tyrosine phosphatases in breast cancer. Oncotarget 2012, 3, 514–515. [Google Scholar]
- Bentires-Alj, M.; Paez, J.G.; David, F.S.; Keilhack, H.; Halmos, B.; Naoki, K.; Maris, J.M.; Richardson, A.; Bardelli, A.; Sugarbaker, D.J.; et al. Activating mutations of the noonan syndrome-associated SHP2/PTPN11 gene in human solid tumors and adult acute myelogenous leukemia. Cancer Res. 2004, 64, 8816–8820. [Google Scholar]
- Ikram, M.K.; Sim, X.; Jensen, R.A.; Cotch, M.F.; Hewitt, A.W.; Ikram, M.A.; Wang, J.J.; Klein, R.; Klein, B.E.; Breteler, M.M.; et al. Four novel Loci (19q13, 6q24, 12q24, and 5q14) influence the microcirculation in vivo. PLoS Genet. 2010, 6, e1001184. [Google Scholar] [CrossRef]
- Petoumenos, K.; Worm, S.W. HIV infection, aging and cardiovascular disease: Epidemiology and prevention. Sex. Health 2011, 8, 465–473. [Google Scholar]
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Adams, R.J.; Berry, J.D.; Brown, T.M.; Carnethon, M.R.; Dai, S.; de Simone, G.; Ford, E.S.; et al. Heart disease and stroke statistics—2011 update: A report from the American Heart Association. Circulation 2011, 123, e18–e209. [Google Scholar] [CrossRef]
- Rai, M.; Baker, W.L.; Parker, M.W.; Heller, G.V. Meta-analysis of optimal risk stratification in patients >65 years of age. Am. J. Cardiol. 2012, 110, 1092–1099. [Google Scholar]
- Weiss, L.A.; Pan, L.; Abney, M.; Ober, C. The sex-specific genetic architecture of quantitative traits in humans. Nat. Genet. 2006, 38, 218–222. [Google Scholar]
- Huang, Y.; Zhou, J.; Ye, H.; Xu, L.; Le, Y.; Yang, X.; Xu, W.; Huang, X.; Lian, J.; Duan, S. Relationship between chemokine (C-X-C motif) ligand 12 gene variant (rs1746048) and coronary heart disease: Case-control study and meta-analysis. Gene 2013, 521, 38–44. [Google Scholar]
- Zhou, J.; Huang, Y.; Huang, R.S.; Wang, F.; Xu, L.; Le, Y.; Yang, X.; Xu, W.; Huang, X.; Lian, J.; et al. A case-control study provides evidence of association for a common SNP rs974819 in PDGFD to coronary heart disease and suggests a sex-dependent effect. Thromb. Res. 2012, 130, 602–606. [Google Scholar]
- Zhang, L.N.; Liu, P.P.; Zhou, J.; Huang, R.S.; Yuan, F.; Fei, L.J.; Huang, Y.; Xu, L.; Hao, L.M.; Qiu, X.J.; et al. Positive correlation between variants of lipid metabolismrelated genes and coronary heart disease. Mol. Med. Rep. 2013, 8, 260–266. [Google Scholar]
- Peng, P.; Lian, J.; Huang, R.S.; Xu, L.; Huang, Y.; Ba, Y.; Yang, X.; Huang, X.; Dong, C.; Zhang, L.; et al. Meta-analyses of KIF6 Trp719Arg in coronary heart disease and statin therapeutic effect. PLoS One 2012, 7, e50126. [Google Scholar]
- Gloria-Bottini, F.; Banci, M.; Saccucci, P.; Papetti, F.; Neri, A.; Pietroiusti, A.; Magrini, A.; Bottini, E. The interaction of ACP1, ADA1, diabetes and gender in coronary artery disease. Am. J. Med. Sci. 2010, 340, 103–108. [Google Scholar] [CrossRef]
- Alonso, A.; Sasin, J.; Bottini, N.; Friedberg, I.; Osterman, A.; Godzik, A.; Hunter, T.; Dixon, J.; Mustelin, T. Protein tyrosine phosphatases in the human genome. Cell 2004, 117, 699–711. [Google Scholar]
- Chong, Z.Z.; Wang, S.; Shang, Y.C.; Maiese, K. Targeting cardiovascular disease with novel SIRT1 pathways. Future Cardiol. 2012, 8, 89–100. [Google Scholar]
- Park, J.B.; Kang, D.Y.; Yang, H.M.; Cho, H.J.; Park, K.W.; Lee, H.Y.; Kang, H.J.; Koo, B.K.; Kim, H.S. Serum alkaline phosphatase is a predictor of mortality, myocardial infarction, or stent thrombosis after implantation of coronary drug-eluting stent. Eur. Heart J. 2013, 34, 920–931. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, X.; Zhan, Q.; Duan, X.; Yang, Q.; Xia, J. Association of PTEN genetic polymorphisms with atherosclerotic cerebral infarction in the Han Chinese population. J. Clin. Neurosci. 2012, 19, 1641–1645. [Google Scholar]
- Saccucci, P.; Banci, M.; Cozzoli, E.; Neri, A.; Magrini, A.; Bottini, E.; Gloria-Bottini, F. Atherosclerosis and PTPN22: A study in coronary artery disease. Cardiology 2011, 119, 54–56. [Google Scholar]
- Saccucci, P.; Banci, M.; Amante, A.; Bottini, E.; Gloria-Bottini, F. Coronary artery disease: Evidence of interaction between PTPN22 and p53 genetic polymorphisms. Cardiology 2011, 120, 166–168. [Google Scholar]
- Zhou, X.; Xu, W.; Chen, J. The 981C>T polymorphism in protein tyrosine phosphatase 1B is associated with decreased risk of coronary artery disease in Chinese Han population. Atherosclerosis 2011, 218, 147–150. [Google Scholar]
- Tasic, I.; Milojkovic, M.; Sunder-Plassmann, R.; Lazarevic, G.; Tasic, N.M.; Stefanovic, V. The association of PC-1 (ENPP1) K121Q polymorphism with metabolic syndrome in patients with coronary heart disease. Clin. Chim. Acta 2007, 377, 237–242. [Google Scholar]
- Polzonetti, V.; Passini, V.; Lucarini, N. Association between ACP(1) genetic polymorphism and favism. Genet. Mol. Res. 2011, 10, 878–884. [Google Scholar]
- Gloria-Bottini, F.; Spina, C.; Nicotra, M.; Saccucci, P.; Ambrosi, S.; Bottini, E. Acid phosphatase locus 1 genetic polymorphism and cancer grading. Am. J. Med. Sci. 2012, 344, 32–34. [Google Scholar]
- Gloria-Bottini, F.; Saccucci, P.; Magrini, A.; Bottini, E. Is there a role of ACP1-ADA1 genetic complex in immune reaction? Association with T1D and with past malarial morbidity. Am. J. Med. Sci. 2010, 340, 268–270. [Google Scholar]
- Shu, Y.H.; Hartiala, J.; Xiang, A.H.; Trigo, E.; Lawrence, J.M.; Allayee, H.; Buchanan, T.A.; Bottini, N.; Watanabe, R.M. Evidence for sex-specific associations between variation in acid phosphatase locus 1 (ACP1) and insulin sensitivity in Mexican-Americans. J. Clin. Endocrinol. Metab. 2009, 94, 4094–4102. [Google Scholar]
- Emilsson, V.; Thorleifsson, G.; Zhang, B.; Leonardson, A.S.; Zink, F.; Zhu, J.; Carlson, S.; Helgason, A.; Walters, G.B.; Gunnarsdottir, S.; et al. Genetics of gene expression and its effect on disease. Nature 2008, 452, 423–428. [Google Scholar] [CrossRef]
- Qi, L.; Parast, L.; Cai, T.; Powers, C.; Gervino, E.V.; Hauser, T.H.; Hu, F.B.; Doria, A. Genetic susceptibility to coronary heart disease in type 2 diabetes: 3 independent studies. J. Am. Coll. Cardiol. 2011, 58, 2675–2682. [Google Scholar]
- Pechlivanis, S.; Muhleisen, T.W.; Mohlenkamp, S.; Schadendorf, D.; Erbel, R.; Jockel, K.H.; Hoffmann, P.; Nothen, M.M.; Scherag, A.; Moebus, S; et al. Risk loci for coronary artery calcification replicated at 9p21 and 6q24 in the Heinz Nixdorf Recall Study. BMC Med. Genet. 2013, 14, 23. [Google Scholar] [CrossRef]
- Kathiresan, S.; Voight, B.F.; Purcell, S.; Musunuru, K.; Ardissino, D.; Mannucci, P.M.; Anand, S.; Engert, J.C.; Samani, N.J.; Schunkert, H.; et al. Genome-wide association of early-onset myocardial infarction with single nucleotide polymorphisms and copy number variants. Nat. Genet. 2009, 41, 334–341. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lee, B.S.; Shin, D.J.; Woo Park, K.; Shin, Y.A.; Joong Kim, K.; Heo, L.; Young Lee, J.; Kyoung Kim, Y.; Jin Kim, Y.; et al. A genome-wide association study of a coronary artery disease risk variant. J. Hum. Genet. 2013, 58, 120–126. [Google Scholar] [CrossRef]
- Brasil, A.S.; Pereira, A.C.; Wanderley, L.T.; Kim, C.A.; Malaquias, A.C.; Jorge, A.A.; Krieger, J.E.; Bertola, D.R. PTPN11 and KRAS gene analysis in patients with Noonan and Noonan-like syndromes. Genet. Test. Mol. Biomark. 2010, 14, 425–432. [Google Scholar]
- Papadopoulou, A.; Issakidis, M.; Gole, E.; Kosma, K.; Fryssira, H.; Fretzayas, A.; Nicolaidou, P.; Kitsiou-Tzeli, S. Phenotypic spectrum of 80 Greek patients referred as Noonan syndrome and PTPN11 mutation analysis: The value of initial clinical assessment. Eur. J. Pediatr. 2012, 171, 51–58. [Google Scholar]
- Sarkozy, A.; Conti, E.; Seripa, D.; Digilio, M.C.; Grifone, N.; Tandoi, C.; Fazio, V.M.; di Ciommo, V.; Marino, B.; Pizzuti, A.; et al. Correlation between PTPN11 gene mutations and congenital heart defects in Noonan and LEOPARD syndromes. J. Med. Genet. 2003, 40, 704–708. [Google Scholar] [CrossRef]
- Daly, M.E. Determinants of platelet count in humans. Haematologica 2011, 96, 10–13. [Google Scholar]
- Jia, Z.F.; Cao, X.Y.; Cao, D.H.; Kong, F.; Kharbuja, P.; Jiang, J. Polymorphisms of PTPN11 gene could influence serum lipid levels in a sex-specific pattern. Lipids Health Dis. 2013, 12, 72. [Google Scholar] [CrossRef]
- Zaid, G.; Yehudai, D.; Rosenschein, U.; Zeina, A.R. Coronary artery disease in an asymptomatic population undergoing a multidetector computed tomography (MDCT) coronary angiography. Open Cardiovasc. Med. J. 2010, 4, 7–13. [Google Scholar]
- Higgs, Z.C.; Macafee, D.A.; Braithwaite, B.D.; Maxwell-Armstrong, C.A. The Seldinger technique: 50 years on. Lancet 2005, 366, 1407–1409. [Google Scholar]
- Gabriel, S.; Ziaugra, L.; Tabbaa, D. SNP genotyping using the Sequenom MassARRAY iPLEX platform. Curr. Protoc. Hum. Genet. 2009. [Google Scholar] [CrossRef]
- Huang, Y.; Yu, X.; Wang, L.; Zhou, S.; Sun, J.; Feng, N.; Nie, S.; Wu, J.; Gao, F.; Fei, B.; et al. Four genetic polymorphisms of lymphotoxin-alpha gene and cancer risk: A systematic review and meta-analysis. PLoS One 2013, 8, e82519. [Google Scholar]
- Lian, J.; Xu, L.; Huang, Y.; Le, Y.; Jiang, D.; Yang, X.; Xu, W.; Huang, X.; Dong, C.; Ye, M.; et al. Meta-analyses of HFE variants in coronary heart disease. Gene 2013, 527, 167–173. [Google Scholar] [CrossRef]
- Lian, J.; Huang, Y.; Huang, R.S.; Xu, L.; Le, Y.; Yang, X.; Xu, W.; Huang, X.; Ye, M.; Zhou, J.; et al. Meta-analyses of four eosinophil related gene variants in coronary heart disease. J. Thromb. Thrombolysis 2013, 36, 394–401. [Google Scholar]
- Huang, Y.; Lian, J.; Huang, R.S.; Wang, F.; Xu, L.; Le, Y.; Yang, X.; Xu, W.; Huang, X.; Ye, M.; et al. Positive association between rs10918859 of the NOS1AP gene and coronary heart disease in male Han Chinese. Genet. Test. Mol. Biomark. 2013, 17, 25–29. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Han, X.; Zhang, L.; Zhang, Z.; Zhang, Z.; Wang, J.; Yang, J.; Niu, J. Association between Phosphatase Related Gene Variants and Coronary Artery Disease: Case-Control Study and Meta-Analysis. Int. J. Mol. Sci. 2014, 15, 14058-14076. https://doi.org/10.3390/ijms150814058
Han X, Zhang L, Zhang Z, Zhang Z, Wang J, Yang J, Niu J. Association between Phosphatase Related Gene Variants and Coronary Artery Disease: Case-Control Study and Meta-Analysis. International Journal of Molecular Sciences. 2014; 15(8):14058-14076. https://doi.org/10.3390/ijms150814058
Chicago/Turabian StyleHan, Xia, Lijun Zhang, Zhiqiang Zhang, Zengtang Zhang, Jianchun Wang, Jun Yang, and Jiamin Niu. 2014. "Association between Phosphatase Related Gene Variants and Coronary Artery Disease: Case-Control Study and Meta-Analysis" International Journal of Molecular Sciences 15, no. 8: 14058-14076. https://doi.org/10.3390/ijms150814058
APA StyleHan, X., Zhang, L., Zhang, Z., Zhang, Z., Wang, J., Yang, J., & Niu, J. (2014). Association between Phosphatase Related Gene Variants and Coronary Artery Disease: Case-Control Study and Meta-Analysis. International Journal of Molecular Sciences, 15(8), 14058-14076. https://doi.org/10.3390/ijms150814058