Expression Profiling of Exosomal miRNAs Derived from Human Esophageal Cancer Cells by Solexa High-Throughput Sequencing

Abstract
:1. Introduction
2. Results
2.1. Characterization of Exosomes Released by Esophageal Cancer Cells

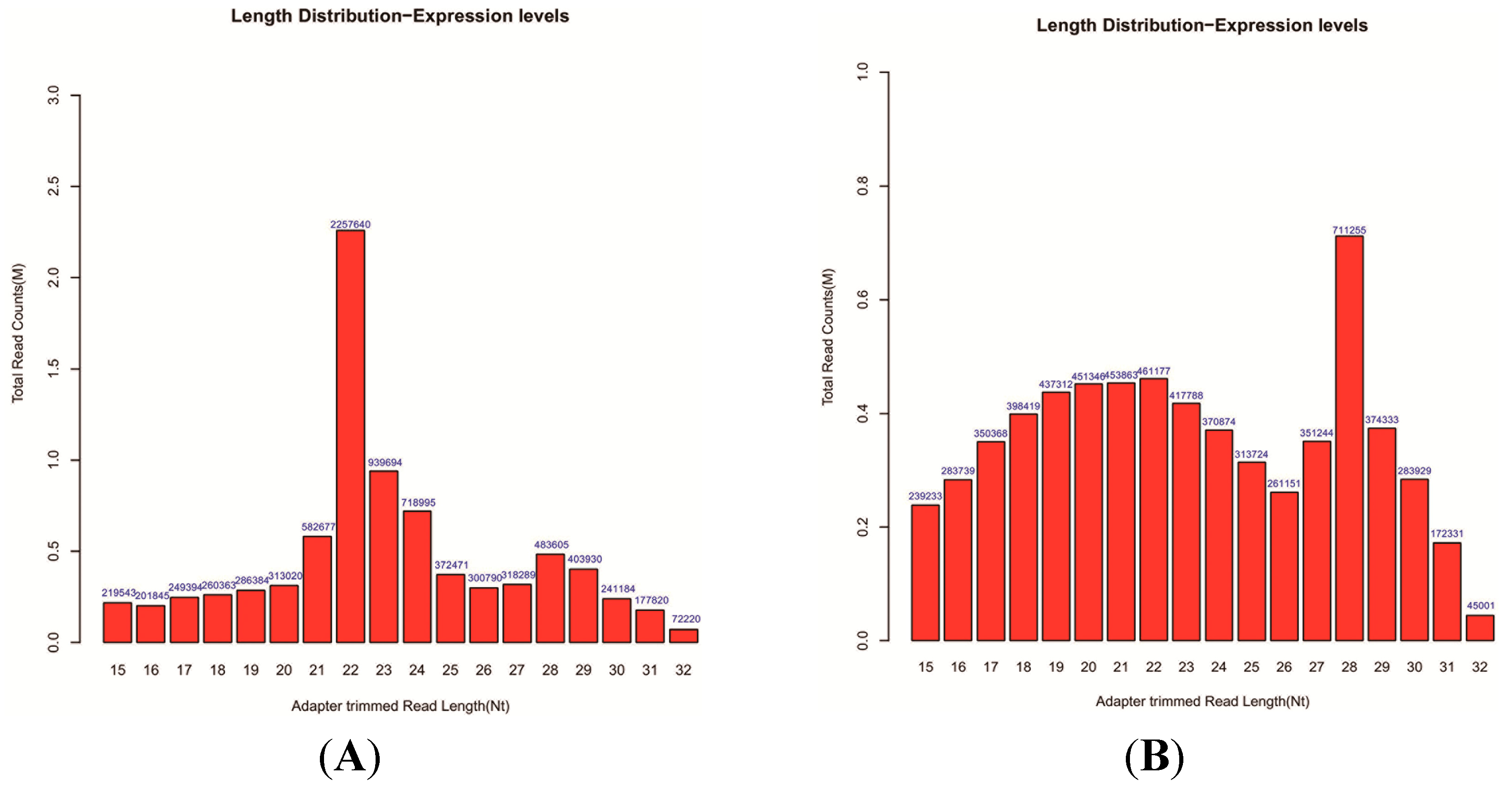
2.2. Overview of Small RNA Sequencing Data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
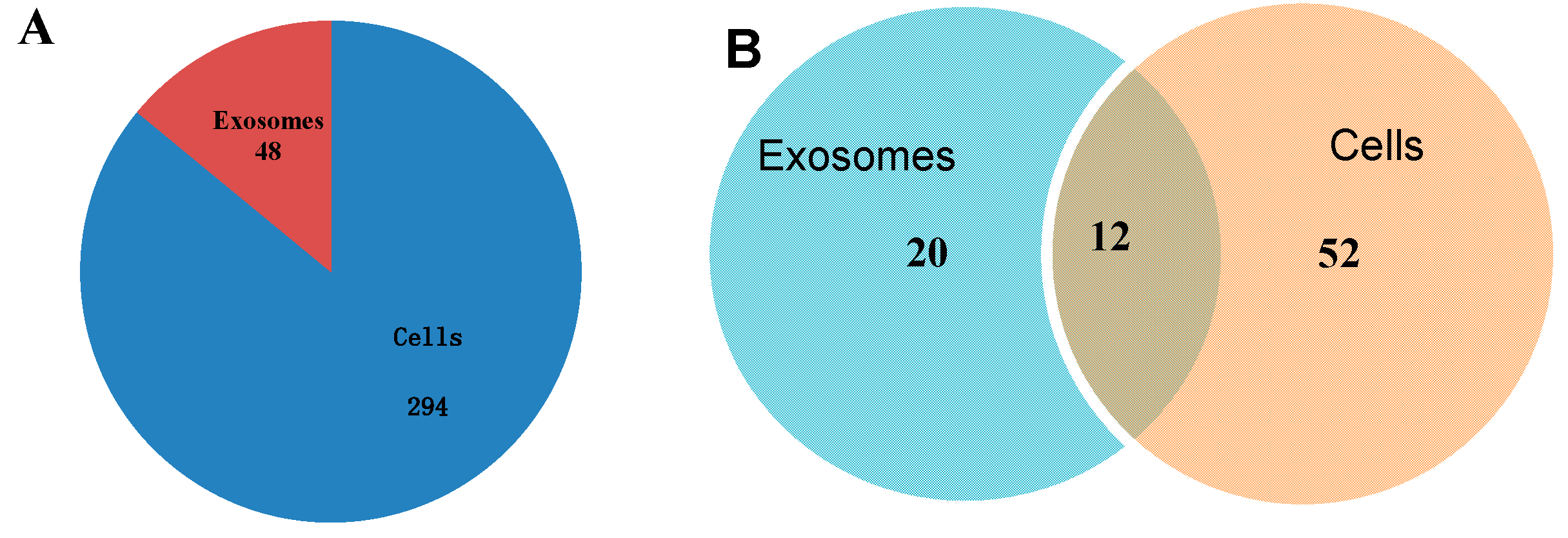{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Small RNAs | Cells | Exosomes | ||
|---|---|---|---|---|
| Number of Unique Reads | Total Number of Reads | Number of Unique Reads | Total Number of Reads | |
| Clean Reads | 13,088,424 | 7,736,476 | ||
| Adaptor-Trimmed Reads (≥15 bp) | 1,919,950 | 9,595,761 | 1,226,905 | 7,193,132 |
| Protein-Coding mRNA | 59,655 (3.11%) | 92,372 (0.96%) | 5988 (0.49%) | 21,742 (0.30%) |
| Repbase | 31,421 (1.64%) | 106,725 (1.11%) | 5066 (0.41%) | 14,454 (0.20%) |
| miRNA | 56,421 (2.94%) | 3,163,192 (32.96%) | 7726 (0.63%) | 94,141 (1.31%) |
| rRNA | 260,307 (13.56%) | 1,423,180 (14.83%) | 85,303 (6.95%) | 542,830 (7.55%) |
| tRNA | 80,635 (4.20%) | 856,084 (9.02%) | 119,500 (9.74%) | 2,325,394 (32.33%) |
| snRNA | 27,935 (1.45%) | 297,457 (3.10%) | 1823 (0.15%) | 5472 (0.08%) |
| Unannotated Reads | 1,403,502 (73.10%) | 3,647,636 (38.01%) | 1,001,224 (81.61%) | 4,188,375 (58.23%) |

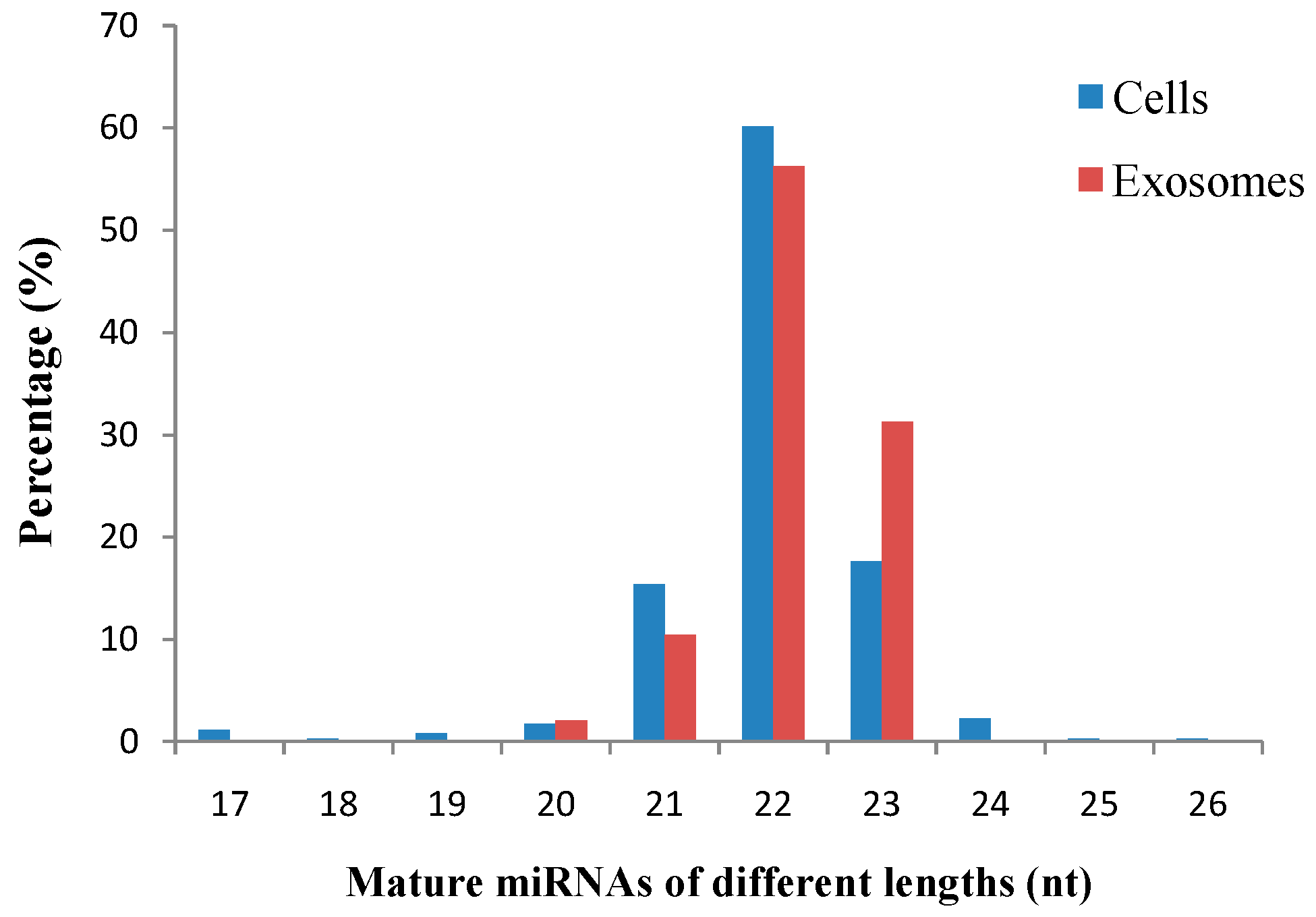
2.3. Identification of Known miRNAs



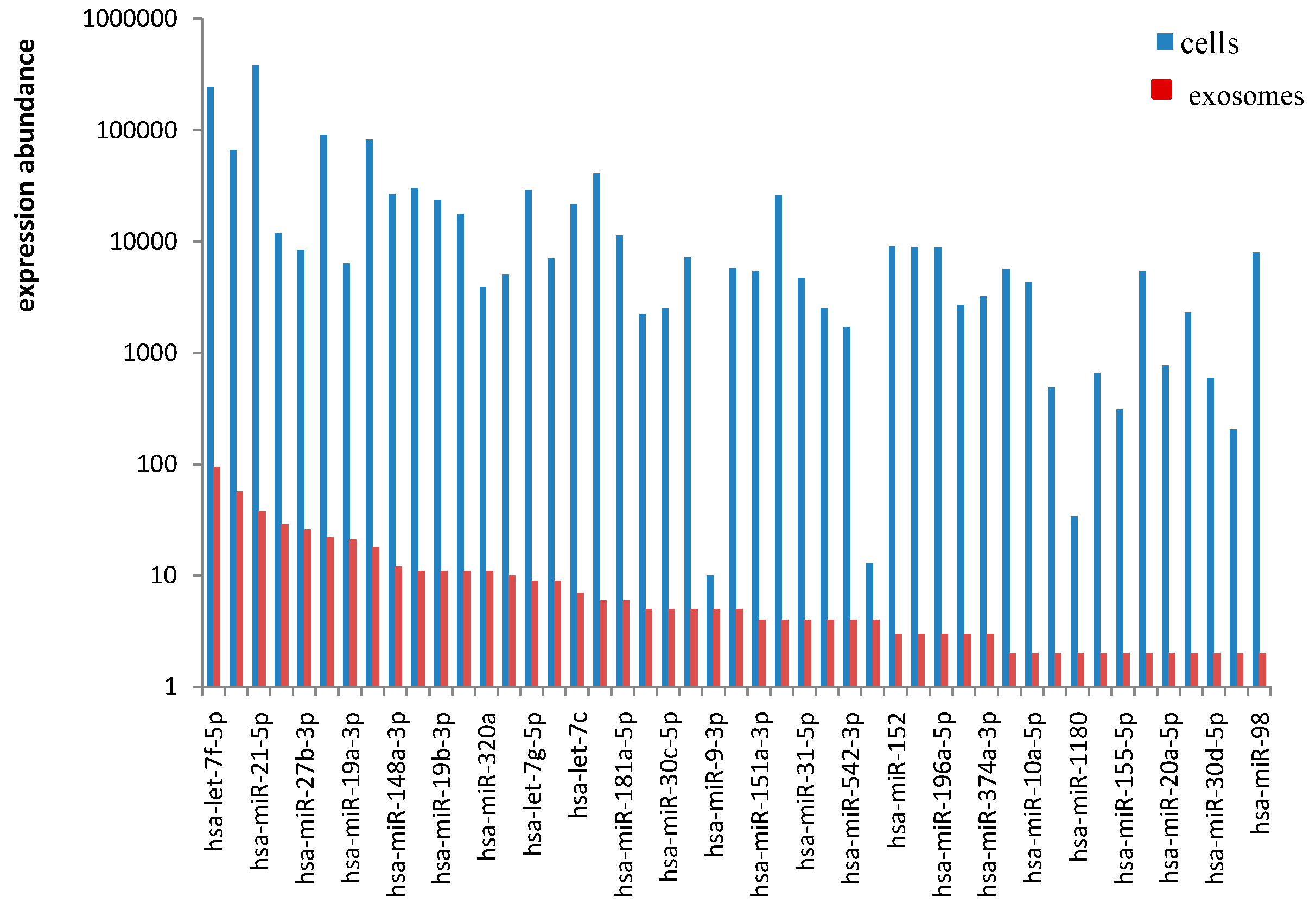
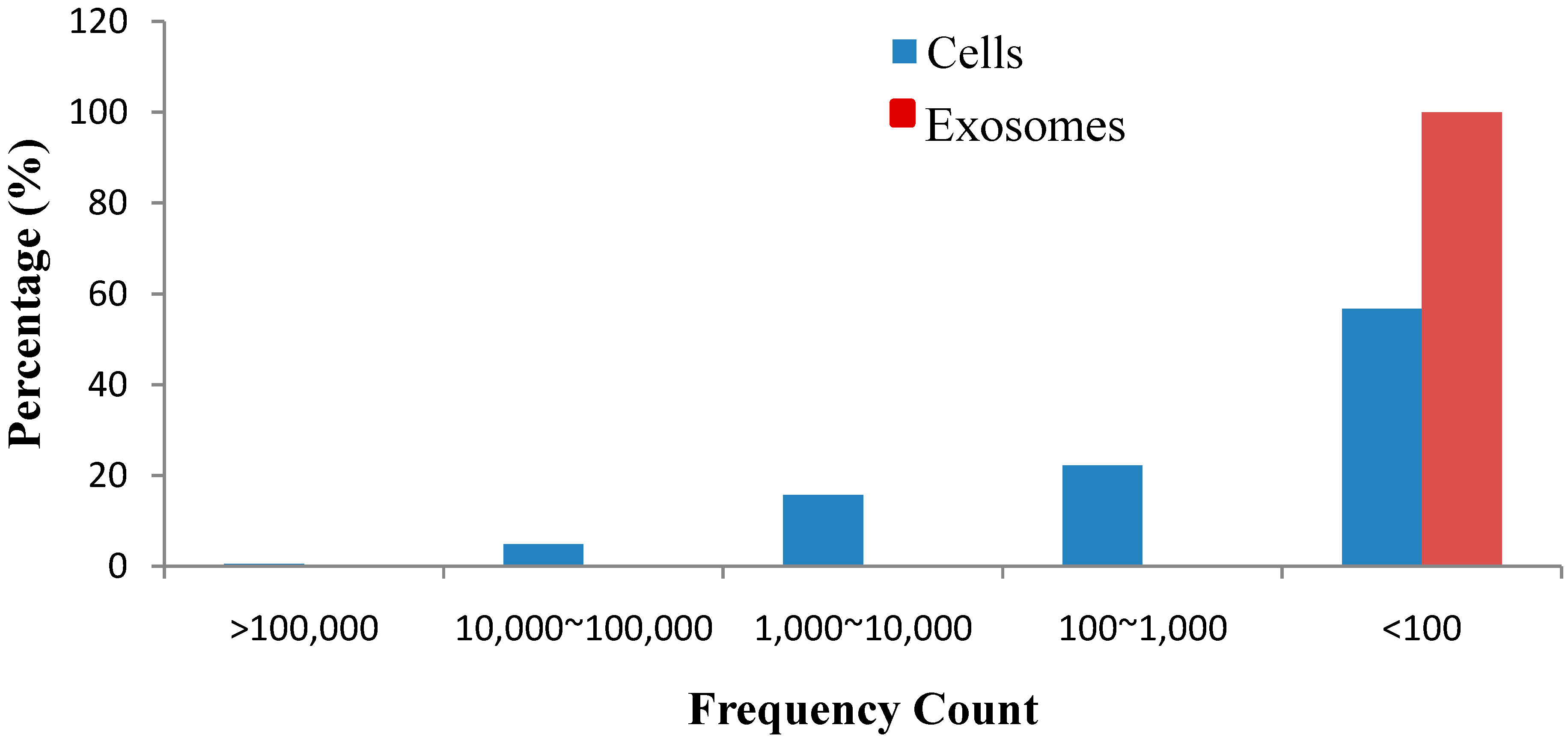
2.4. EC9706 and Their Corresponding Exosomes Contain a Subset of Dysregulated miRNA

| miRNA Annotation | Pre-miRNA Arm (5p or 3p) | Mature MicroRNA Seed | Transcript Sequence | Is miRBase Mature miRNA the Most Abundant Sequence? | Intracellular Transcript Number |
|---|---|---|---|---|---|
| hsa-miR-21-5p | 5p | AGCTTA | TAGCTTATCAGACTGATGTTGA | Yes | 382,634 |
| hsa-let-7f-5p | 5p | GAGGTA | TGAGGTAGTAGATTGTATAGTT | Yes | 243,882 |
| hsa-let-7b-5p | 5p | GAGGTA | TGAGGTAGTAGGTTGTGTGGTT | Yes | 91,479 |
| hsa-miR-100-5p | 5p | ACCCGT | AACCCGTAGATCCGAACTTGTG | Yes | 82,325 |
| hsa-let-7a-5p | 5p | GAGGTA | TGAGGTAGTAGGTTGTATAGTT | Yes | 66,589 |
| hsa-miR-125b-5p | 5p | CCCTGA | TCCCTGAGACCCTAACTTGTGA | Yes | 41,096 |
| hsa-let-7i-5p | 5p | GAGGTA | TGAGGTAGTAGTTTGTGCTGTT | Yes | 30,233 |
| hsa-let-7g-5p | 5p | GAGGTA | TGAGGTAGTAGTTTGTACAGTT | Yes | 28,900 |
| hsa-miR-148a-3p | 3p | CAGTGC | TCAGTGCACTACAGAACTTTGT | Yes | 26,923 |
| hsa-miR-24-3p | 3p | GGCTCA | TGGCTCAGTTCAGCAGGAACAG | Yes | 26,085 |
| hsa-miR-19b-3p | 3p | GTGCAA | TGTGCAAATCCATGCAAAACTGA | Yes | 23,649 |
| hsa-let-7c | 5p | GAGGTA | TGAGGTAGTAGGTTGTATGGTT | Yes | 21,557 |
| hsa-miR-25-3p | 3p | ATTGCA | CATTGCACTTGTCTCGGTCTGA | Yes | 17,757 |
| hsa-miR-182-5p | 5p | TTGGCA | TTTGGCAATGGTAGAACTCACACT | Yes | 15,213 |
| hsa-miR-425-5p | 5p | ATGACA | AATGACACGATCACTCCCGTTGA | No | 12,236 |
| hsa-miR-26a-5p | 5p | TCAAGT | TTCAAGTAATCCAGGATAGGCT | Yes | 11,993 |
| hsa-miR-181a-5p | 5p | ACATTC | AACATTCAACGCTGTCGGTGAGT | Yes | 11,329 |
| hsa-miR-99a-5p | 5p | ACCCGT | AACCCGTAGATCCGATCTTGTG | Yes | 10,476 |
| hsa-miR-103a-3p | 3p | GCAGCA | AGCAGCATTGTACAGGGCTATGA | Yes | 10,305 |
| miRNA Annotation | Pre-miRNA Arm (5p or 3p) | Mature MicroRNA Seed | Transcript Sequence | Is miRBase Mature miRNA the Most Abundant Sequence? | Extracellular Transcript Number | |
|---|---|---|---|---|---|---|
| hsa-let-7f-5p | 5p | GAGGTA | TGAGGTAGTAGATTGTATAGTT | Yes | 95 | |
| hsa-let-7a-5p | 5p | GAGGTA | TGAGGTAGTAGGTTGTATAGTT | Yes | 57 | |
| hsa-miR-21-5p | 5p | AGCTTA | TAGCTTATCAGACTGATGTTGA | Yes | 38 | |
| hsa-miR-26a-5p | 5p | TCAAGT | TTCAAGTAATCCAGGATAGGCT | Yes | 29 | |
| hsa-miR-27b-3p | 3p | TCACAG | TTCACAGTGGCTAAGTTCTGC | Yes | 26 | |
| hsa-let-7b-5p | 5p | GAGGTA | TGAGGTAGTAGGTTGTGTGGTT | Yes | 22 | |
| hsa-miR-19a-3p | 3p | GTGCAA | TGTGCAAATCTATGCAAAACTGA | Yes | 21 | |
| hsa-miR-100-5p | 5p | ACCCGT | AACCCGTAGATCCGAACTTGTG | Yes | 18 | |
| hsa-miR-148a-3p | 3p | CAGTGC | TCAGTGCACTACAGAACTTTGT | Yes | 12 | |
| hsa-let-7i-5p | 5p | GAGGTA | TGAGGTAGTAGTTTGTGCTGTT | Yes | 11 | |
| hsa-miR-19b-3p | 3p | GTGCAA | TGTGCAAATCCATGCAAAACTGA | Yes | 11 | |
| hsa-miR-25-3p | 3p | ATTGCA | CATTGCACTTGTCTCGGTCTGA | Yes | 11 | |
| hsa-miR-320a | 3p | AAAGCT | AAAAGCTGGGTTGAGAGGGCGA | Yes | 11 | |
| hsa-miR-423-5p | 5p | GAGGGG | TGAGGGGCAGAGAGCGAGACTTT | Yes | 10 | |
| hsa-let-7g-5p | 5p | GAGGTA | TGAGGTAGTAGTTTGTACAGTT | Yes | 9 | |
| hsa-miR-92a-3p | 3p | ATTGCA | TATTGCACTTGTCCCGGCCTGT | Yes | 9 | |
| hsa-let-7c | 5p | GAGGTA | TGAGGTAGTAGGTTGTATGGTT | Yes | 7 | |
| hsa-miR-125b-5p | 5p | CCCTGA | TCCCTGAGACCCTAACTTGTGA | Yes | 6 | |
| hsa-miR-181a-5p | 5p | ACATTC | AACATTCAACGCTGTCGGTGAGT | Yes | 6 | |
| Mature MicroRNA ID | Pre-miRNA Arm (5p or 3p) | Mature MicroRNA Seed | Most Abundant Sequence (isomiR) | Is miRBase Mature miRNA the Most Abundant Sequence? | Cell Counts | Exosome Counts |
|---|---|---|---|---|---|---|
| hsa-miR-chr12_3064 | 3p | GAGAGG | GGAGAGGTGGATGAGTGGTTTA | No | 0 | 339 |
| hsa-miR-chr1_9487 | 3p | GAGAGG | GGAGAGGTGGATGAGTGGTTTA | No | 0 | 339 |
| hsa-miR-chr15_4885 | 3p | TTCAAG | GTTCAAGTCCAGCTGGG | Yes | 30 | 306 |
| hsa-miR-chr3_14243 | 3p | TTCAAG | GTTCAAGTCCAGCTGGG | Yes | 30 | 306 |
| hsa-miR-chr18_7463 | 3p | ACGTGA | CACGTGAAACCCTGTCTGAAT | No | 14 | 137 |
| hsa-miR-chr2_11998 | 5p | AGGACT | AAGGCAGGACTGGTGACTGGGGTG | No | 7 | 59 |
| hsa-miR-chrY_24624 | 3p | TCCCTG | GTGTCCCTGGTTCGAGCCC | No | 0 | 55 |
| hsa-miR-chr6_18451 | 3p | GTCGTG | GGTCGTGGGTTCGAGC | No | 0 | 37 |
| hsa-miR-chr20_11216 | 3p | GTTCGA | GGTTCAAATCCTGTCTTCT | No | 2 | 28 |
| hsa-miR-chr1_9933 | 5p | TCTTTG | ATCTCTTTGAGTTCTCACCA | No | 2 | 19 |
2.5. Prediction of Novel miRNAs
| Candidate miRNA ID | Precursor Coordinate | Consensus Mature Sequence | Length | Pre-miRNA Arm (5p or 3p) | Cell Counts | Exosome Counts |
|---|---|---|---|---|---|---|
| hsa-miR-chr18_7463 | chr18:42911426..42911487: + | CACGTGAAACCCTGTCT | 17 | 3p | 14 | 137 |
| hsa-miR-chr2_11998 | chr2:9376082..9376166: + | CAGGACTGGGGACTGGGGTG | 20 | 5p | 7 | 59 |
| hsa-miR-chr20_11216 | chr20:39392006..39392060: − | GGTTCGAATCCTGTCTTCT | 19 | 3p | 2 | 28 |
| hsa-miR-chr1_9933 | chr1:45874140..45874224: − | CTCTTTGAGTTCTCACCA | 18 | 5p | 2 | 19 |
| hsa-miR-chr5_17564 | chr5:33712846..33712885: − | GTACTCAAGAGGCTGAAGA | 19 | 5p | 7 | 18 |
| hsa-miR-chr1_8755 | chr1:62567393..62567462: + | TCAAATCCTGTCTGACC | 17 | 3p | 2 | 17 |
| hsa-miR-chr7_20613 | chr7:25989517..25989563: − | TCAGTGCACTACAGAACTTTGT | 22 | 5p | 26,923 | 12 |
| hsa-miR-chr11_1761 | chr11:31858407..31858458: − | GCATGGGTGGTTCAGTGGTAGAATT | 25 | 5p | 202 | 5 |
| hsa-miR-chr19_8205 | chr19:13947044..13947124: − | TGGCTCAGTTCAGCAGGAACAG | 22 | 5p | 26,085 | 4 |
| hsa-miR-chr16_5626 | chr16:14397874..14397964: + | AACTGGCCCTCAAAGTCCCGCT | 22 | 5p | 5460 | 2 |
| Candidate miRNA ID | Precursor Coordinate | Consensus Mature Sequence | Length | Pre-miRNA Arm (5p or 3p) | Cell Counts | Exosome Counts |
|---|---|---|---|---|---|---|
| hsa-miR-chr15_4885 | chr15:43193729..43193791: + | GTTCAAGTCCAGCTGGG | 17 | 3p | 30 | 306 |
| hsa-miR-chr3_14243 | chr3:101824879..101824941: + | GTTCAAGTCCAGCTGGG | 17 | 3p | 30 | 306 |
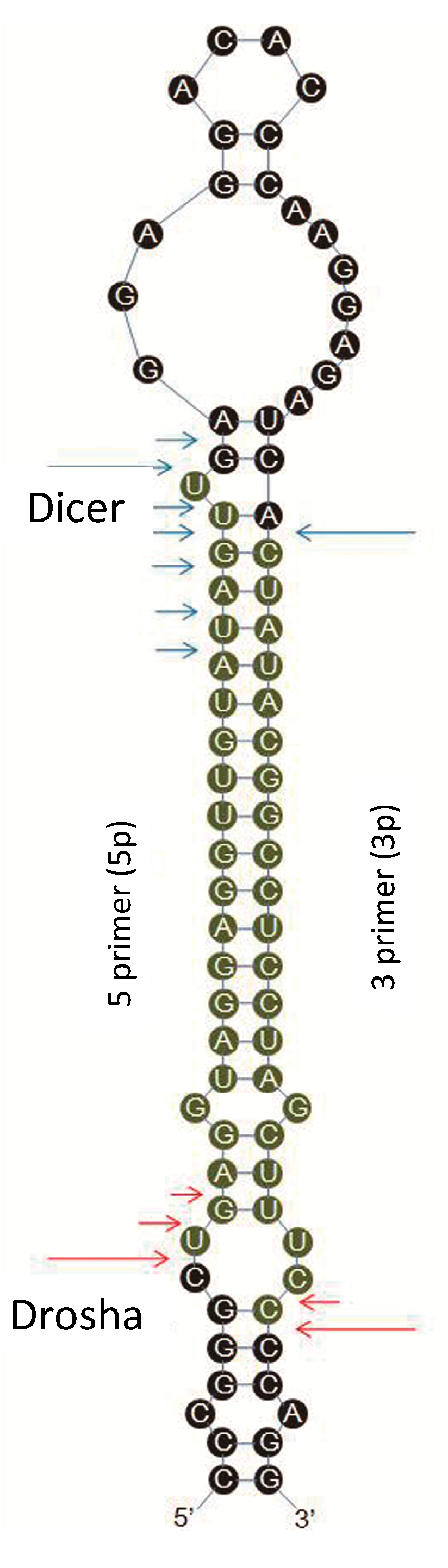
2.6. Sequence Variations in miRNAs

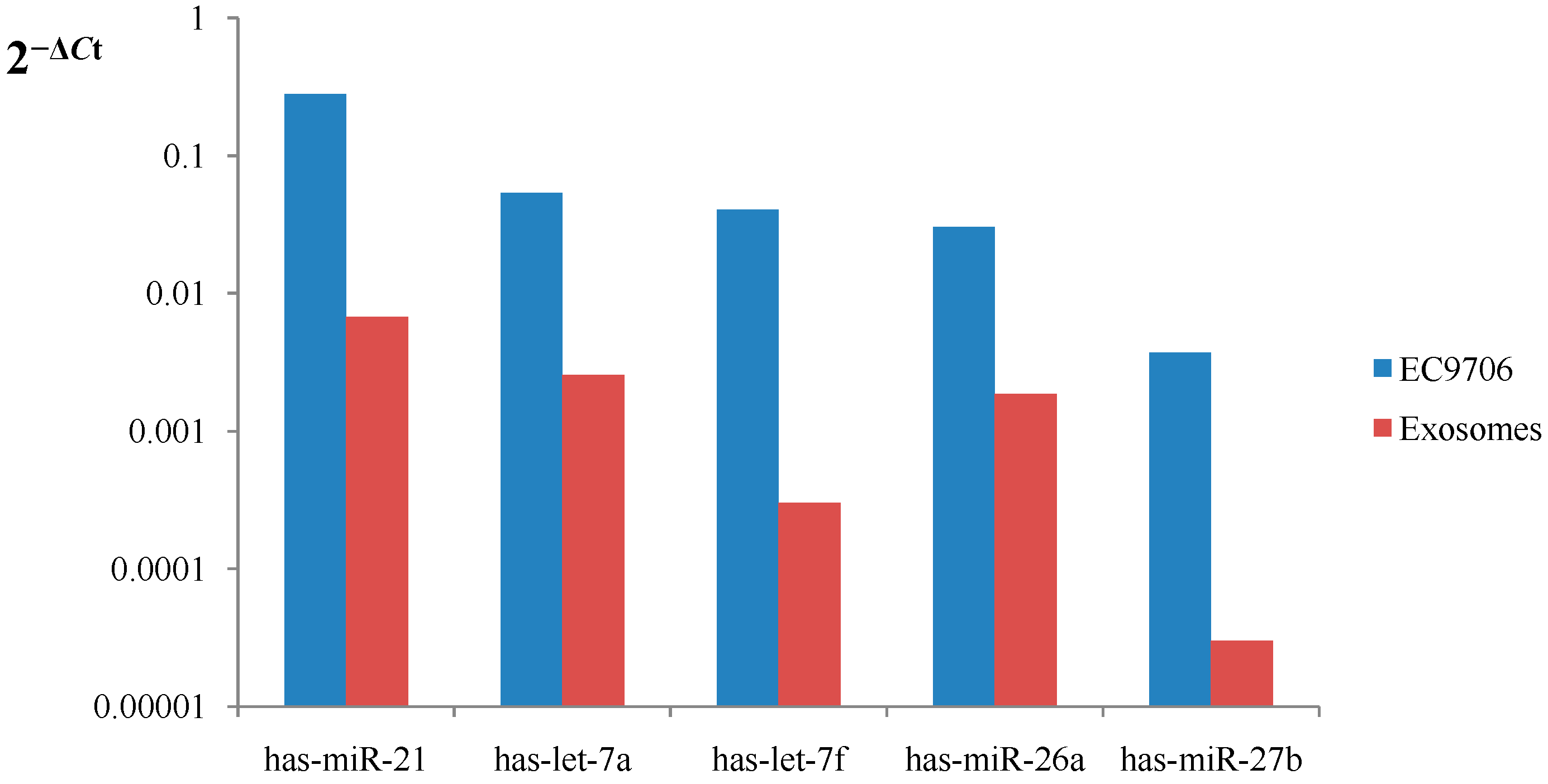
2.7. miRNA Validation Assays by qRT-PCR

3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. Exosome Isolation
4.3. Transmission Electron Microscopy
4.4. Western Blot
4.5. RNA Isolation and Analysis
4.6. Small RNA Libraries Construction and Solexa Sequencing
4.7. Bioinformation Analysis of Small RNA Data
4.8. Expression Patterns of Known and Novel miRNAs
4.9. miRNA Variants
4.10. Measurement of miRNA Expression Level by qRT-PCR

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteomics 2010, 73, 1907–1920. [Google Scholar]
- Zhang, H.; Xie, Y.; Li, W.; Chibbar, R.; Xiong, S.; Xiang, J. CD4+ T cell-released exosomes inhibit CD8+ cytotoxic T-lymphocyte responses and antitumor immunity. Cell Mol. Immunol. 2011, 8, 23–30. [Google Scholar]
- Zumaquero, E.; Muñoz, P.; Cobo, M.; Lucena, G.; Pavón, E.J.; Martín, A.; Navarro, P.; García-Pérez, A.; Ariza-Veguillas, A.; Malavasi, F.; et al. Exosomes from human lymphoblastoid B cells express enzymatically active CD38 that is associated with signaling complexes containing CD81, Hsc-70 and Lyn. Exp. Cell. Res. 2010, 316, 2692–2706. [Google Scholar]
- Park, J.A.; Sharif, A.S.; Tschumperlin, D.J.; Lau, L.; Limbrey, R.; Howarth, P.; Drazen, J.M. Tissue factor–bearing exosome secretion from human mechanically stimulated bronchial epithelial cells in vitro and in vivo. J. Allergy Clin. Immunol. 2012, 130, 1375–1383. [Google Scholar] [CrossRef]
- Tauro, B.J.; Greening, D.W.; Mathias, R.A.; Ji, H.; Mathivanan, S.; Scott, A.M.; Simpson, R.J. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods 2012, 56, 293–304. [Google Scholar]
- Kogure, T.; Patel, T. Isolation of extracellular nanovesicle microRNA from liver cancer cells in culture. Methods Mol. Biol. 2013, 1024, 11–18. [Google Scholar]
- Kruger, S.; Elmageed, Z.Y.; Hawke, D.H.; Wörner, P.M.; Jansen, D.A.; Abdel-Mageed, A.B.; Alt, E.U.; Izadpanah, R. Molecular characterization of exosome-like vesicles from breast cancer cells. BMC Cancer 2014, 14, 44. [Google Scholar] [CrossRef]
- Lässer, C.; Alikhani, V.S.; Ekström, K.; Eldh, M.; Paredes, P.T.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human saliva, plasma and breast milk exosomes contain RNA: Uptake by macrophages. J. Transl. Med. 2011, 9, 9. [Google Scholar]
- Wada, J.; Onishi, H.; Suzuki, H.; Yamasaki, A.; Nagai, S.; Morisaki, T.; Katano, M. Surface-bound TGF-β1 on effusion-derived exosomes participates in maintenance of number and suppressive function of regulatory T-cells in malignant effusions. Anticancer Res. 2010, 30, 3747–3757. [Google Scholar]
- Leväne, B.; Bhakta, N.R.; Torregrosa Paredes, P.; Barbeau, R.; Hiltbrunner, S.; Pollack, J.L.; Magnus Sköld, C.; Svartengren, M.; Grunewald, J.; Gabrielsson, S.; et al. Altered microRNA profiles in bronchoalveolar lavage fluid exosomes in asthmatic patients. J. Allergy Clin. Immunol. 2013, 131, 894–903. [Google Scholar]
- Alvarez, S.; Suazo, C.; Boltansky, A.; Ursu, M.; Carvajal, D.; Innocenti, G.; Vukusich, A.; Hurtado, M.; Villanueva, S.; Carreño, J.E.; et al. Urinary exosomes as a source of kidney dysfunction biomarker in renal transplantation. Transplant. Proc. 2013, 45, 3719–3723. [Google Scholar]
- Ogawa, Y.; Taketomi, Y.; Murakami, M.; Tsujimoto, M.; Yanoshita, R. Small RNA transcriptomes of two types of exosomes in human whole saliva determined by next generation sequencing. Biol. Pharm. Bull. 2013, 36, 66–75. [Google Scholar]
- Van Niel, G.; Porto-Carreiro, I.; Simoes, S.; Raposo, G. Exosomes: A common pathway for a specialized function. J. Biochem. 2006, 140, 13–21. [Google Scholar]
- Silva, J.; Garcia, V.; Rodriguez, M.; Compte, M.; Cisneros, E.; Veguillas, P.; Garcia, J.M.; Dominguez, G.; Campos-Martin, Y.; Cuevas, J.; et al. Analysis of exosome release and its prognostic value in human colorectal cancer. Genes Chromosom. Cancer 2012, 51, 409–418. [Google Scholar]
- Inal, J.M.; Kosgodage, U.; Azam, S.; Stratton, D.; Antwi-Baffour, S.; Lange, S. Blood/plasma secretome and microvesicles. Biochim. Biophys. Acta 2013, 1834, 2317–2325. [Google Scholar]
- Ohshima, K.; Inoue, K.; Fujiwara, A.; Hatakeyama, K.; Kanto, K.; Watanabe, Y.; Muramatsu, K.; Fukuda, Y.; Ogura, S.; Yamaguchi, K.; et al. Let-7 microRNA family is selectively secreted into the extracellular environment via exosomes in a metastatic gastric cancer cell line. PLoS One 2010, 5, e13247. [Google Scholar]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef]
- Tanaka, Y.; Kamohara, H.; Kinoshita, K.; Kurashige, J.; Ishimoto, T.; Iwatsuki, M.; Watanabe, M.; Baba, H. Clinical impact of serum exosomal microrna-21 as a clinical biomarker in human esophageal squamous cell carcinoma. Cancer 2013, 119, 1159–1167. [Google Scholar]
- Keller, S.; König, A.K.; Marmé, F.; Runz, S.; Wolterink, S.; Koensgen, D.; Mustea, A.; Sehouli, J.; Altevogt, P. Systemic presence and tumor-growth promoting effect of ovarian carcinoma released exosomes. Cancer Lett. 2009, 278, 73–81. [Google Scholar]
- Yi, S.; Gao, Z.X.; Zhao, H.; Zeng, C.; Luo, W.; Chen, B.; Wang, W.M. Identification and characterization of microRNAs involved in growth of blunt snout bream (Megalobrama amblycephala) by Solexa sequencing. BMC Genomics 2013, 14, 754. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, J.H.; Zheng, Y.S.; Zhang, P.; Chen, X.; Wu, J.; Xu, L.; Luo, X.Q.; Ke, Z.Y.; Zhou, H.; et al. Genome-wide analysis of small RNA and novel microRNA discovery in human acute lymphoblastic leukemia based on extensive sequencing approach. PLoS One 2009, 4, e6849. [Google Scholar]
- Jenjaroenpun, P.; Kremenska, Y.; Nair, V.M.; Kremenskoy, M.; Joseph, B.; Kurochkin, I.V. Characterization of RNA in exosomes secreted by human breast cancer cell lines using next-generation sequencing. Peer J. 2013, 1, e201. [Google Scholar]
- Song, C.; Wang, C.; Zhang, C.; Korir, N.K.; Yu, H.; Ma, Z.; Fang, J. Deep sequencing discovery of novel and conserved microRNAs in trifoliate (Citrus trifoliata). BMC Genomics 2010, 11, 431. [Google Scholar] [CrossRef]
- Pigati, L.; Yaddanapudi, S.C.; Iyengar, R.; Kim, D.J.; Hearn, S.A.; Danforth, D.; Hastings, M.L.; Duelli, D.M. Selective release of microRNA species from normal and malignant mammary epithelial cells. PLoS One 2010, 5, e13515. [Google Scholar]
- Yao, J.; Wang, Y.; Wang, W.; Wang, N.; Li, H. Solexa sequencing analysis of chicken pre-adipocyte microRNAs. Biosci. Biotechnol. Biochem. 2011, 75, 54–61. [Google Scholar]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar]
- Fernandez-Valverde, S.L.; Taft, R.J.; Mattick, J.S. Dynamic isomiR regulation in Drosophila development. RNA 2010, 16, 1881–1888. [Google Scholar]
- Xie, S.S.; Li, X.Y.; Liu, T.; Cao, J.H.; Zhong, Q.; Zhao, S.H. Discovery of porcine microRNAs in multiple tissues by a Solexa deep sequencing approach. PLoS One 2011, 6, e16235. [Google Scholar]
- Sakai, N.S.; Samia-Aly, E.; Barbera, M.; Fitzgerald, R.C. A review of the current understanding and clinical utility of miRNAs in esophageal cancer. Semin. Cancer Biol. 2013, 23, 512–521. [Google Scholar]
- Hessvik, N.P.; Phuyal, S.; Brech, A.; Sandvig, K.; Llorente, A. Profiling of microRNAs in exosomes released from PC-3 prostate cancer cells. Biochim. Biophys. Acta 2012, 1819, 1154–1163. [Google Scholar] [CrossRef]
- Komatsu, S.; Ichikawa, D.; Takeshita, H.; Morimura, R.; Hirajim, S.; Tsujiura, M.; Kawaguchi, T.; Miyamae, M.; Nagata, H.; Konishi, H.; et al. Circulating miR-18a: A sensitive cancer screening biomarker in human cancer. In Vivo 2014, 28, 293–297. [Google Scholar]
- Ogata-Kawata, H.; Izumiya, M.; Kurioka, D.; Honma, Y.; Yamada, Y.; Furuta, K.; Gunji, T.; Ohta, H.; Okamoto, H.; Sonoda, H.; et al. Circulating exosomal microRNAs as biomarkers of colon cancer. PLoS One 2014, 9, e92921. [Google Scholar]
- Takeshita, N.; Hoshino, I.; Mori, M.; Akutsu, Y.; Hanari, N.; Yoneyama, Y.; Ikeda, N.; Isozaki, Y.; Maruyama, T.; Akanuma, N.; et al. Serum microRNA expression profile: miR-1246 as a novel diagnostic and prognostic biomarker for oesophageal squamous cell carcinoma. Br. J. Cancer 2013, 108, 644–652. [Google Scholar]
- Taylor, D.D.; Gercel-Taylor, C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol. Oncol. 2008, 110, 13–21. [Google Scholar]
- Bellingham, S.A.; Coleman, B.M.; Hill, A.F. Small RNA deep sequencing reveals a distinct miRNA signature released in exosomes from prion-infected neuronal cells. Nucleic Acids Res. 2012, 40, 10937–10949. [Google Scholar]
- Chiba, M.; Kimura, M.; Asari, S. Exosomes secreted from human colorectal cancer cell lines contain mRNAs, microRNAs and natural antisense RNAs, that can transfer into the human hepatoma HepG2 and lung cancer A549 cell lines. Oncol. Rep. 2012, 28, 1551–1558. [Google Scholar]
- Li, P.; Mao, W.M.; Zheng, Z.G.; Dong, Z.M.; Ling, Z.Q. Down-regulation of PTEN expression modulated by dysregulated miR-21 contributes to the progression of esophageal cancer. Dig. Dis. Sci. 2013, 58, 3483–3493. [Google Scholar]
- Vogt, P.K.; Gymnopoulos, M.; Hart, J.R. PI 3-kinase and cancer: Changing accents. Curr. Opin. Genet. Dev. 2009, 19, 12–17. [Google Scholar]
- Yan, L.X.; Wu, Q.N.; Zhang, Y.; Li, Y.Y.; Liao, D.Z.; Hou, J.H.; Fu, J.; Zeng, M.S.; Yun, J.P.; Wu, Q.L.; et al. Knockdown of miR-21 in human breast cancer cell lines inhibits proliferation, in vitro migration and in vivo tumor growth. Breast Cancer Res. 2011, 13, R2. [Google Scholar] [CrossRef]
- Kobayashi, M.; Salomon, C.; Tapia, J.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E. Ovarian cancer cell invasiveness is associated with discordant exosomal sequestration of Let-7 miRNA and miR-200. J. Transl. Med. 2014, 12, 4. [Google Scholar]
- Colamaio, M.; Calì, G.; Sarnataro, D.; Borbone, E.; Pallante, P.; Decaussin-Petrucci, M.; Nitsch, L.; Croce, C.M.; Battista, S.; Fusco, A. Let-7a down-regulation plays a role in thyroid neoplasias of follicular histotype affecting cell adhesion and migration through its ability to target the FXYD5 (Dysadherin) gene. J. Clin. Endocrinol. Metab. 2012, 97, E2168–E2178. [Google Scholar] [CrossRef]
- Gao, J.; Li, L.; Wu, M.; Liu, M.; Xie, X.; Guo, J.; Tang, H.; Xie, X. miR-26a inhibits proliferation and migration of breast cancer through repression of MCL-1. PLoS One 2013, 8, e65138. [Google Scholar]
- Zhang, B.; Liu, X.X.; He, J.R.; Zhou, C.X.; Guo, M.; He, M.; Li, M.F.; Chen, G.Q.; Zhao, Q. Pathologically decreased miR-26a antagonizes apoptosis and facilitates carcinogenesis by targeting MTDH and EZH2 in breast cancer. Carcinogenesis 2011, 32, 2–9. [Google Scholar]
- Namwat, N.; Chusorn, P.; Loilome, W.; Techasen, A.; Puetkasichonpasutha, J.; Pairojkul, C.; Khuntikeo, N.; Yongvanit, P. Expression profiles of oncomir miR-21 and tumor suppressor let-7a in the progression of opisthorchiasis-associated cholangiocarcinoma. Asian Pac. J. Cancer Prev. 2012, 13, 65–69. [Google Scholar]
- Kida, K.; Nakajjima, M.; Mohri, T.; Oda, Y.; Takagi, S.; Fukam, T.; Yokoi, T. PPARα is regulated by miR-21 and miR-27b in human liver. Pharm. Res. 2011, 28, 2467–2476. [Google Scholar]
- Henson, B.J.; Bhattacharjee, S.; O’Dee, D.M.; Feingold, E.; Gollin, S.M. Decreased expression of miR-125b and miR-100 in oral cancer cells contributes to malignancy. Genes Chromosom. Cancer 2009, 48, 569–582. [Google Scholar]
- Cloonan, N.; Wani, S.; Xu, Q.; Gu, J.; Lea, K.; Heater, S.; Barbacioru, C.; Steptoe, A.L.; Martin, H.C.; Nourbakhsh, E.; et al. MicroRNAs and their isomiRs function cooperatively to target common biological pathways. Genome Biol. 2011, 12, R126. [Google Scholar]
- Camussi, G.; Dereqibus, M.C.; Bruno, S.; Cantaluppi, V.; Biancone, L. Exosomes/microvesicles as a mechanism of cell-to-cell communication. Kidney Int. 2010, 78, 838–848. [Google Scholar]
- Kosaka, N.; Iquchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar]
- Manterola, L.; Guruceaga, E.; Gállego Pérez-Larraya, J.; González-Huarriz, M.; Jauregui, P.; Tejada, S.; Diez-Valle, R.; Segura, V.; Samprón, N.; Barrena, C.; et al. A small noncoding RNA signature found in exosomes of GBM patient serum as a diagnostic tool. Neuro Oncol. 2014, 16, 520–527. [Google Scholar]
- Que, R.; Ding, G.; Chen, J.; Cao, L. Analysis of serum exosomal microRNAs and clinicopathologic features of patients with pancreatic adenocarcinoma. World J. Surg. Oncol. 2013, 11, 219. [Google Scholar]
- Rabinowits, G.; Gerçel-Taylor, C.; Day, J.M.; Taylor, D.D.; Kloecker, G.H. Exosomal microRNA: A diagnostic marker for lung cancer. Clin. Lung Cancer 2009, 10, 42–46. [Google Scholar] [CrossRef]
- Kroh, E.M.; Parkin, R.K.; Mitchell, P.S.; Tewari, M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010, 50, 298–301. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liao, J.; Liu, R.; Yin, L.; Pu, Y. Expression Profiling of Exosomal miRNAs Derived from Human Esophageal Cancer Cells by Solexa High-Throughput Sequencing. Int. J. Mol. Sci. 2014, 15, 15530-15551. https://doi.org/10.3390/ijms150915530
Liao J, Liu R, Yin L, Pu Y. Expression Profiling of Exosomal miRNAs Derived from Human Esophageal Cancer Cells by Solexa High-Throughput Sequencing. International Journal of Molecular Sciences. 2014; 15(9):15530-15551. https://doi.org/10.3390/ijms150915530
Chicago/Turabian StyleLiao, Juan, Ran Liu, Lihong Yin, and Yuepu Pu. 2014. "Expression Profiling of Exosomal miRNAs Derived from Human Esophageal Cancer Cells by Solexa High-Throughput Sequencing" International Journal of Molecular Sciences 15, no. 9: 15530-15551. https://doi.org/10.3390/ijms150915530
APA StyleLiao, J., Liu, R., Yin, L., & Pu, Y. (2014). Expression Profiling of Exosomal miRNAs Derived from Human Esophageal Cancer Cells by Solexa High-Throughput Sequencing. International Journal of Molecular Sciences, 15(9), 15530-15551. https://doi.org/10.3390/ijms150915530