1. Introduction
Exosomes are small (40–100 nm) membrane vesicles that are derived from the invagination of endosomal compartments called multivesicular bodies and then released into the extracellular environment by most cell types [
1]. Recent studies have reported that exosomes exist in the culture supernatant of numerous eukaryotic cells, including cytotoxic T-lymphocytes [
2], B-lymphocytes [
3], epithelial cells [
4] and tumor cells [
5,
6,
7]. Exosomes have also been found present in physiological and pathological fluids, including plasma, urine, saliva, breast milk, malignant effusions and bronchoalveolar lavage fluid [
8,
9,
10,
11]. Exosomes mediate the disposal of obsolete membranes from original cells [
12], aid in intercellular communication, modulate selected cellular activities [
13] or modify their growth and mobility capacities [
14] and stimulate the proliferation, survival and adhesion of target cells [
15]. However, the biological functions of exosomes remain unclear. A recent study has shown that cellular gene products, such as proteins, mRNAs and microRNAs (miRNAs), are packaged inside exosomes and are delivered into recipient cells, where they exert their respective biological effects [
16]. The expression of peripheral blood miRNAs from tumor cells can remain stable, because the extracellular nucleic acids in the bloodstream are highly protected by exosomes, microvesicle-like structures that can resist the degradation of various enzymes [
17].
Esophageal cancer is a common type of cancer worldwide. However, esophageal cancer with distant metastasis and local invasion is still associated with a poor prognosis. To date, endoscopic and radiologic examinations are still being used as early detection methods. Simple and non-invasive diagnostic methods, such as blood and urine esophageal squamous cell carcinoma (ESCC) screening, are currently unavailable [
18]. Tumor-derived exosomes may become systemic by distribution through the blood stream; hence, exosomes in the blood or other body fluids of tumor patients can be used for clinical testing [
19]. The miRNA profiles of tumor-derived circulating exosomes isolated from body fluids may be used as novel diagnostic biomarkers when tumor cell materials are inaccessible. This strategy can also aid in the development of novel tumor therapeutics.
Next-generation RNA sequencing technology has rapidly progressed in recent years and is now widely used in various biological applications. This technology allows the rapid, sensitive and precise procurement of all RNA subtypes, as well as unannotated novel miRNA candidates or low-abundance RNAs that are weakly expressed [
20,
21,
22]. On the basis of small RNA digital analysis through Solexa high-throughput sequencing, the commercially available high-throughput sequencing technology allows the identification of numerous short RNAs in a sample. miRNA sequences are 19–25 nt long, thus, the Solexa platform is apparently a suitable choice for miRNA discovery [
23]. This technology can be used to explore the specific miRNA expression and distribution characteristics in tumor-derived exosomes and to discover novel miRNAs [
21]. In the present study, Solexa high-throughput sequencing was used to explore the miRNA expression profile and distribution characteristics of exosomes derived from human esophageal cancer cells.
We utilized the next-generation RNA sequencing technology to construct two small-RNA cDNA libraries from human esophageal cancer cells and esophageal cancer-derived exosomes. This technology can describe the expression of miRNAs in both intracellular and extracellular environments of esophageal cancer. We identified miRNAs in the two libraries and analyzed differentially-expressed miRNAs through high-throughput sequencing and bioinformatics analysis. We validated several miRNAs and their expression profiles in esophageal cancer cells and esophageal cancer-derived exosomes through stem-loop RT-PCR. This study provided insights into the involvement of exosome-mediated miRNAs in regulating an esophageal cancer microenvironment. We identified and profiled the RNA species present in exosomes and original cells; our results suggested that exosomal miRNAs can be used as potential esophageal cancer-specific biomarkers.
3. Discussion
miRNAs are important members of small, well-conserved and non-coding RNAs. Their function in cancer pathogenesis as oncogenes or tumor suppressors has been investigated and attracted attention for several years [
29]. With the development of next-generation RNA sequencing technology, an increasing number of miRNAs in tumor-derived exosomes has been discovered [
22,
30]. However, the roles of miRNAs in esophageal cancer-derived exosomes remain unknown. Serum
miR-18a level was significantly higher in patients with esophageal, pancreatic or hepatocellular cancer. The areas under the receiver-operating characteristic curve (ROC) were 0.944, 0.936 and 0.881 in esophageal, pancreatic and hepatocellular, respectively, which showed that serum
miR-18a may serve as a screening biomarker [
31]. Researchers have characterized the miRNA profiles of colorectal cancer (CRC) serum exosomes using microarray analyses to identify several special miRNAs up-regulated in CRC patients and found that the levels of seven miRNAs are significantly down-regulated after surgical resection of tumors. Through ROC analysis,
miR-23a and
miR-1246 were detected to have higher sensitivities of 95% and 90%, respectively, for Stage I samples compared with known tumor markers (CA19-9 and CEA). This finding supports the promising application of
miR-23a and
miR-1246 as diagnostic biomarkers [
32]. Another study on the serum microRNA expression profile of esophageal cancer showed that the expression level of
miR-1246 is higher in ESCC patients than in controls and that this expression level decreases after surgical resection. To assess the diagnostic and prognostic values of serum
miR-1246 in ESCC, the ROC curve assay of
miR-1246 was conducted. Results showed that the sensitivity and specificity of
miR-1246 were 71.3% and 73.9%, respectively, in distinguishing ESCC patients from healthy controls. The abundant exosomal circulating
miR-1246 in serum significantly correlates with the ESCC tumor-node-metastasis stage, and it is the most potent independent risk factor for poor survival [
33]. Tanaka
et al. [
18] analyzed the exosomes in serum from patients who suffered from ESCC and confirmed that exosomal
miR-21 expression is significantly higher in the serum of ESCC patients than in that of benign controls. Exosomal
miR-21 expression is related to clinicopathological stage, tumor classification, positive lymph node status and metastatic status; therefore,
miR-21 has been suggested to be a biomarker for detecting ESCC progression. In the present study, we developed a workflow for analyzing next-generation RNA sequencing of data that focus on miRNAs in human esophageal cancer cells and their corresponding exosomes. Using this technology, we detected 9,595,761 (cells) and 7,193,132 (exosomes) unique small RNAs. This result indicates the presence of a significant number of small RNA in exosomes, among which, miRNAs predominate. This workflow has revealed several trends in known and novel miRNA expression profiles for esophageal cancer-derived exosomes. A total of 80 miRNAs, including 48 known and 32 novel miRNAs, were firstly reported in esophageal cancer cell-derived exosomes. In addition, comparison with the miRBase 18.0 revealed several common miRNAs that were expressed in the cells and exosomes. The expression levels of most known miRNAs were significantly lower in the exosomes than in the cells. We also found several novel miRNAs that presented different profiles between the cells and their corresponding exosomes. Several novel miRNAs were expressed at higher levels in the exosomes than in the cells, implying that these novel miRNAs may be uniquely packaged into exosomes with specialized, but unknown functions. This phenomenon may be also caused by the fact that most studies only focused on miRNAs with high read counts in cells. The studies based on miRNAs are also generally from high to low read counts to establish a miRNA database. Some novel miRNAs are highly expressed in exosomes, but lowly expressed in cells. Taylor
et al. [
34] studied the miRNA expression in ovarian tumor cells and their corresponding exosomes. Among the 218 mature miRNAs that were positive in both cells and exosomes, 12 and 31 were elevated in cells and exosomes, respectively. Bellingham
et al. [
35] proposed a selective mechanism for the incorporation and release of miRNAs in exosomes that contain limited or no
18S and
28S cellular ribosomal species. Not all mRNAs and miRNAs contained within cells can be directly targeted and packaged in exosomes. Therefore, the differential expression profiles of miRNAs in the cells and exosomes probably have different biological functions.
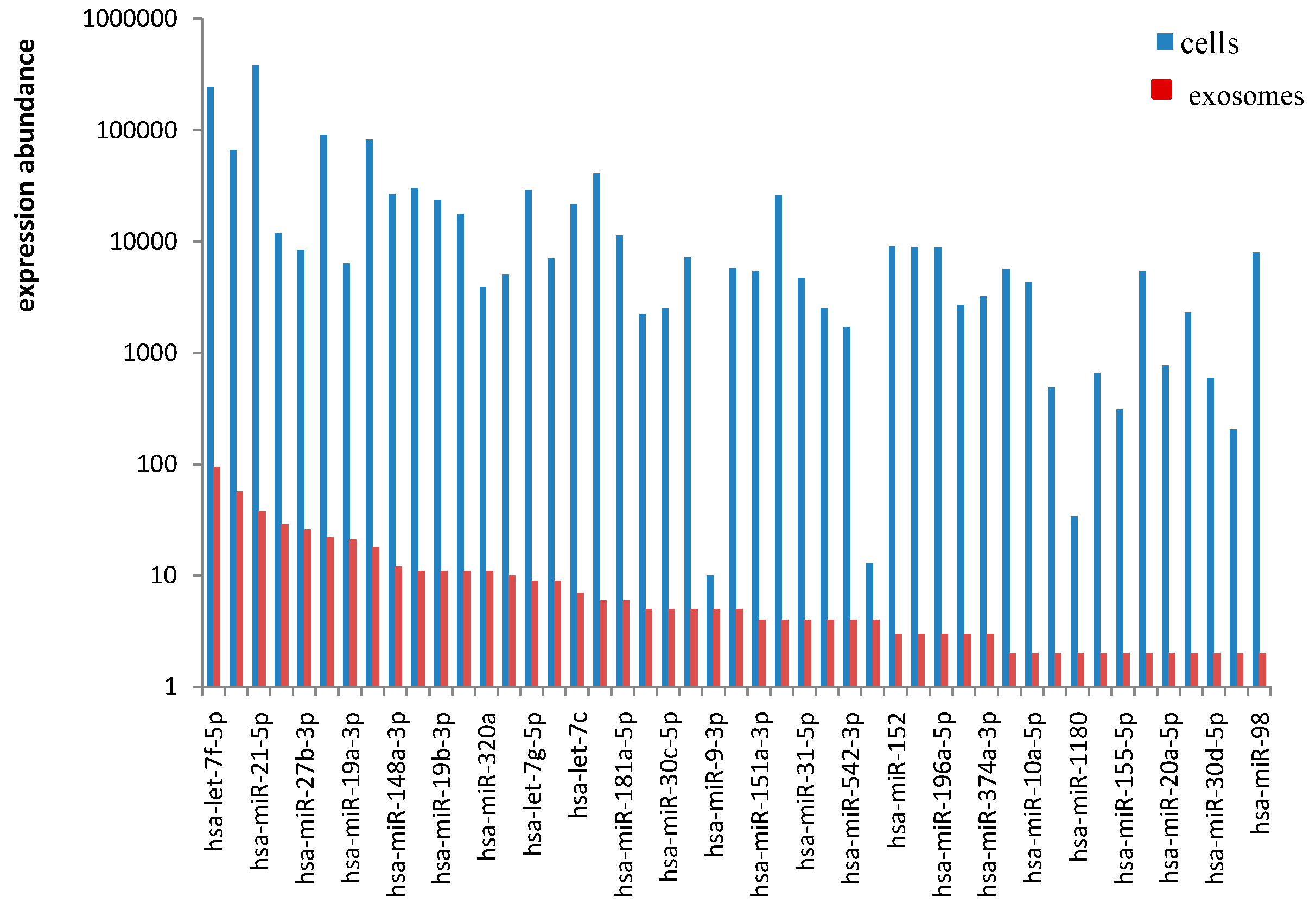
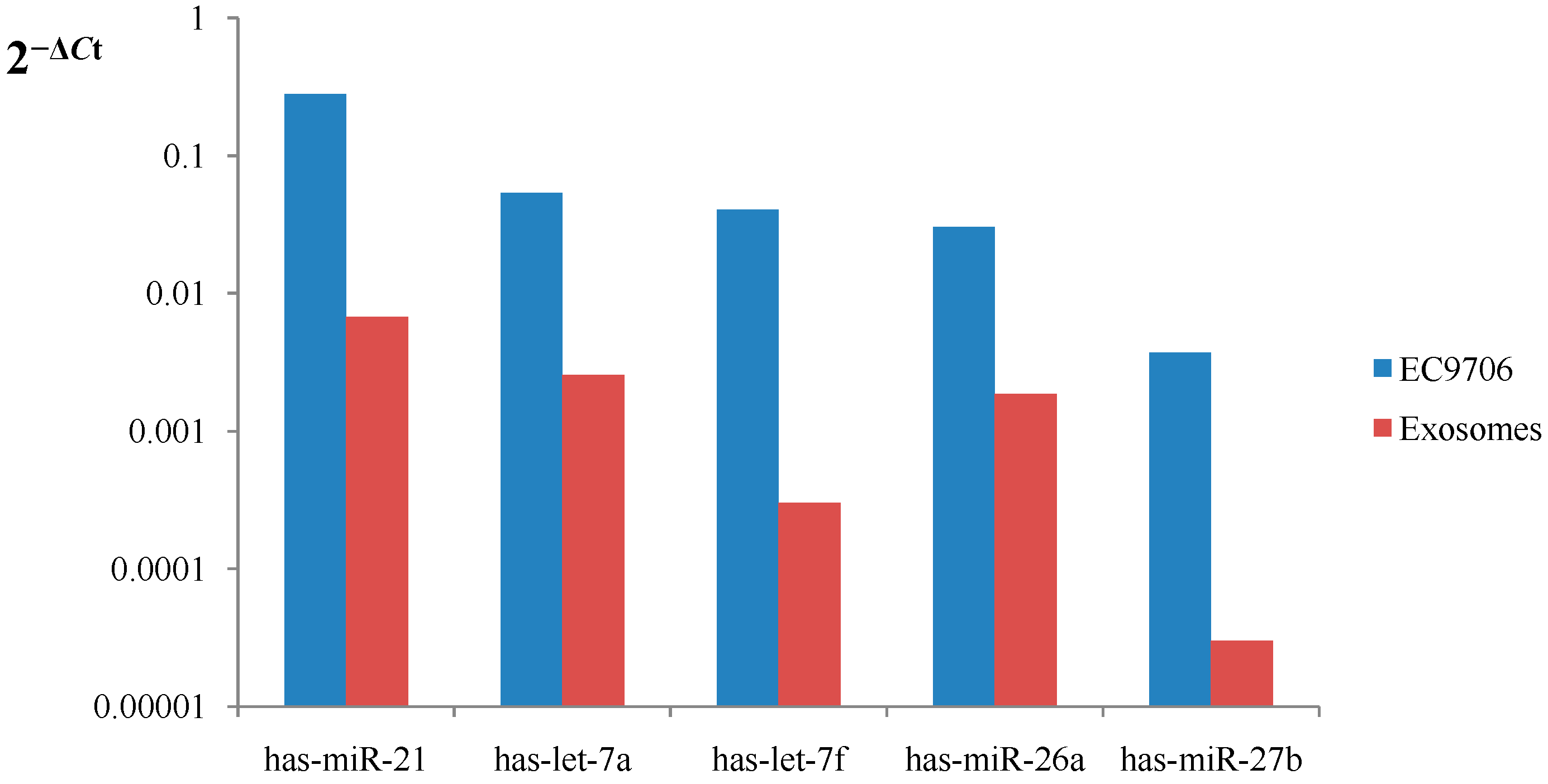
In the present study,
has-miR-21,
has-let-7 family,
miR-26a and
miR-27b were abundant in EC9706 cells and their corresponding exosomes. Known miRNA species have been previously sorted into exosomes in other cell lines. Chiba
et al. [
36] revealed that exosomes derived from the colorectal cancer cell lines contain miRNAs, such as
miR-21, and can be delivered into recipient cells through exosomes. Moreover, studies on
miR-21 and esophageal cancer revealed that
miR-21 is post-transcriptionally regulated by phosphatase and tensin (
PTEN) via binding to the 3'-UTR of
PTEN mRNA. Consequently,
PTEN inhibits tumor cell growth and invasion by blocking the PI3K/AKT pathway [
37,
38]. Yan
et al. [
39] found, through locked nucleic acid silencing combined with microarray technology, that
miR-21 knockdown can inhibit the growth and migration of breast cells
in vitro and tumor growth in nude mice. A previous study found that the
let-7 miRNA family is abundant in both the intracellular and extracellular fractions of a metastatic gastric cancer cell line (AZ-P7a); this study suggested that AZ-P7a cells release
let-7 miRNAs via exosomes into the extracellular environment to maintain their oncogenesis [
16]. Kobayashi
et al. [
40] determined that the miRNA transcripts of the
let-7 family exist in both ovarian cancer cell lines and their exosomes; they established that the release and miRNA content of exosomes significantly differ between ovarian cancer cell lines and correlate with their invasive potential. Colamaio
et al. [
41] demonstrated that the
let-7a overexpression in the follicular thyroid carcinoma cells increases cell adhesion and reduces cell migration; however,
let-7a silencing in normal rat thyroid cells induces the opposite effects through stable transfections.
Has-miR-26a and
has-miR-27b were both found in ovarian tumor cells and their corresponding exosomes; however, their expression levels significantly differ between the two samples [
34]. Several studies on breast cancer have shown that
miR-26a can inhibit cell proliferation, colony formation and migration, as well as promote apoptosis by regulating several carcinogenesis-related processes, including several mechanisms that involve the targeting of
MCL-1,
MTDH and
EZH2 [
42,
43]. The altered expression levels of different miRNAs involved in multiple signaling transduction pathways can determine cancer development and/or progression by interacting with one another. Namwat
et al. [
44] showed that the oncogenic factor
miR-21 is up-regulated, while the tumor suppressor
let-7a is down-regulated in human cholangiocarcinoma tissues. The high level of IL-6 in such a chronic inflammation-related cancer possibly stimulates
miR-21 expression, and
RAS activation is partly correlated with
let-7a down-regulation. Kida
et al. [
45] found that the
PPARα protein level in the human hepatocellular carcinoma cell line HuH7 significantly decreases when the levels of
miR-21 or
miR-27b are over-expressed and inhibited, respectively. This result suggests that the two miRNAs may serve as important regulators for the fatty acid catabolism of liver cells. Another study on miRNAs in oral squamous cell carcinoma (OSCC) reported that increasing
miR-125b and
miR-100 levels reduces cell proliferation; however, co-transfecting the two miRNAs significantly influences proliferation than transfecting OSCC cells individually. Researchers proposed that the reduced proliferation observed in cells co-transfected with
miR-125b and
miR-100 is caused by the additive effect of the two miRNAs on gene expression rather than a synergistic change [
46]. miRNA expression profiles were previously confined to tissues and cells; today, circulating miRNAs reportedly exist in the peripheral blood with exosomes as carriers. The effective delivery of miRNAs via exosomes in the bloodstream has provided a new mechanism for cell-to-cell communication. Investigating the mechanisms by which these candidate miRNAs carried by exosomes function in esophageal cancer development is important.
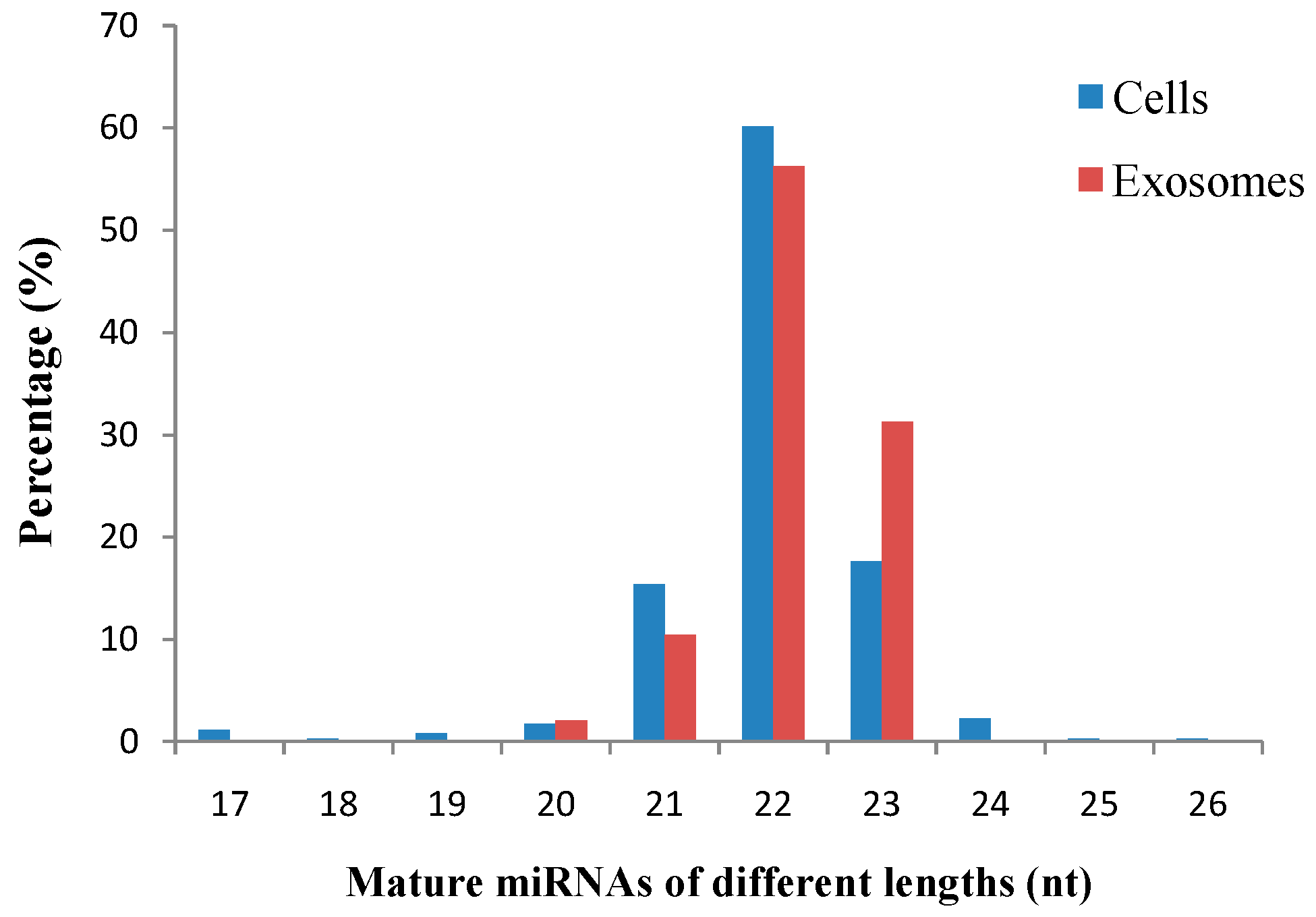
Our study revealed a significant number of isomiRs derived from almost all detected miRNAs. In total, 17,111 known and 5212 novel isomiRs were detected in our two libraries. Most isomiRs showed variability at their 5' and/or 3' ends, likely resulting from the variability in either Dicer or Drosha cleavage positions within the pre-miRNA hairpin; an example of which is given in
Figure 8, which shows sequenced isomiRs for
miR-21 with high read counts in cells and exosomes. This result indicates that the majority of esophageal cancer-derived miRNA nucleotide variants resulted from post-transcriptional modifications. IsomiRs have several biological functions, such as the amelioration of the signal-to-noise ratio in miRNA–mRNA communication by targeting miRNA-controlled genetic networks. A close correlation exists between the expression characteristics of isomiRs and canonical miRNAs. These isomiRs are functional, and their biological roles likely involve the improvement of the signal-to-noise ratio in miRNA–mRNA communication by targeting miRNA-controlled genetic networks [
47]. This finding suggests that isomiRs may have various functions. However, further studies need to be conducted for verification.
Recent studies have suggested that tumor-derived exosomes present a novel mechanism for intercellular communication [
48,
49]. The miRNA profiles of circulating exosomes from the body fluids of patients with tumors have also been proposed to be used as signatures for disease diagnosis. Furthermore, small non-coding RNA signatures exist in microvesicles isolated from glioblastoma multiforme (GBM) patient serum. A study has shown that exosomal miRNA expression levels are significantly higher in 25 patients than in age- and sex-matched healthy controls [
50]. A study on pancreatic adenocarcinoma (PC) revealed that serum exosomal
miR-17-5p and
miR-21 levels are significantly higher in PC patients than in healthy participants and non-primary carcinoma patients; this result suggests that serum exosomal miRNAs may serve as potential biomarkers of PC [
51]. Furthermore, a study on exosomal microRNA in lung cancer indicated that a significant difference in total exosome and exosomal miRNA levels exists between lung cancer patients and controls; this study also noted that exosomal miRNA profiling has the potential to be used as a screening tool for cancer detection because of its minimal invasiveness [
52].
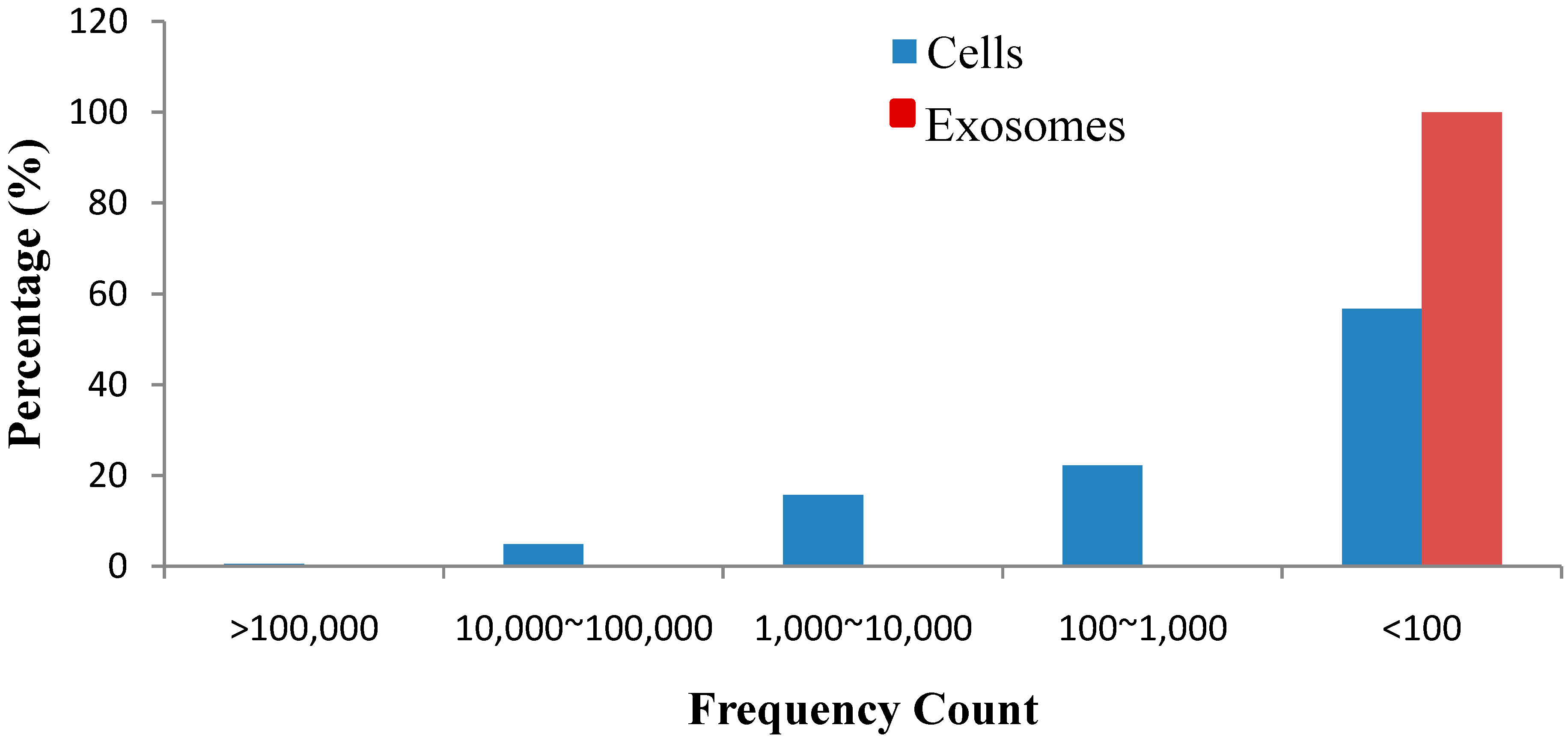
This study is the first to report that exosomes released from esophageal cancer cells contain a series of miRNAs and that the profiles of several exosomal miRNAs resemble those of their parent cells. However, several novel miRNAs in exosomes still need to be investigated. In conclusion, the discovery of these known and novel miRNAs using Solexa high-throughput sequencing provided new insights into the expression profiles and distribution characteristics of miRNAs in esophageal cancer-derived exosomes and afforded significant values to further explore miRNAs in intercellular communication mediated by exosomes in esophageal cancer.
4. Experimental Section
4.1. Cell Culture
The human esophageal cancer cell line EC9706 was purchased from Shanghai Tiancheng Science and Technology Co., Ltd. EC9706 cells were cultured in RPMI-1640 medium (Hyclone, Logan, UT, USA) supplemented with 10% heat-inactivated fetal bovine serum (Hyclone), 100 U/mL penicillin and 100 U/mL streptomycin under a humidified 5% CO2 atmosphere at 37 °C. The cells were centrifuged at 10,000× g for 30 min to remove exosomes in fetal bovine serum and then ultracentrifuged (Beckman Coulter, Brea, CA, USA) at 200,000× g for 6 h to remove bovine-derived exosomes.
4.2. Exosome Isolation
EC9706 cells (40 mL) at a density of 1 × 108 cells per 175-cm2 flask were cultured in complete RPMI-1640 medium at 37 °C and 5% CO2. After 48 h, exosomes were isolated and purified from the culture medium of EC9706 using a sequential centrifugation protocol. Briefly, the culture medium was collected and centrifuged at 300× g for 10 min, 800× g for 10 min, 1200× g for 20 min and 10,000× g for 30 min to remove lifted cells and cellular debris. The supernatant was then ultracentrifuged at 100,000× g for 3 h using a 70Ti rotor (Beckman Coulter) to pelletize the exosomes. The exosome pellets were washed with filtered phosphate-buffered saline (PBS) and then recentrifuged at 100,000× g for 2 h. The supernatant was removed, and the final exosomal pellet was resuspended in 100 μL of PBS. All centrifugation steps were conducted at 4 °C.
4.3. Transmission Electron Microscopy
A 20-μL aliquot of the suspension was loaded onto a carbon-coated grid for 2 min at room temperature. The grid was positioned with the coating side facing the drop containing exosomes. The samples were fixed by covering the grid with 10 μL of 1% aqueous phosphotungstic acid for 1 min and then observed under a transmission electron microscope (Hitachi, Shiga, Japan).
4.4. Western Blot
Samples of cells and exosomes were washed and resuspended in RIPA lysis buffer (Bi Yuntian, Hefei, China) with protease inhibitor mixture. After centrifuging at 14,000× g and for 5 min at 4 °C, the supernatant was collected, and the protein concentration was determined using a BCA Protein Assay Kit (Generay, Shanghai, China). Up to 67 μg of proteins were denatured by boiling in sodium dodecyl sulfate (SDS) loading buffer, loaded onto SDS-polyacrylamide gels and then transferred onto polyvinylidene difluoride membranes (0.45 μm pore size, Millipore, Billerica, MA, USA). The blots were blocked overnight with 5% non-fat dry milk in Tris-buffered saline containing Tween 20. After blocking, the membranes were incubated with primary mouse monoclonal antibody anti-CD63 (1:1000) (sc-51662, Santa Cruz, CA, USA) or mouse anti-β-actin (1:400, internal standard) (BM0627, Boster, Wuhan, China) in TBST for 1.5 h at room temperature. The membranes were washed six times for 5 min each before incubating with the secondary antibody. The secondary antibody conjugated with horseradish peroxidase was incubated at a dilution ratio of 1:5000 for 1 h at room temperature. Immunoreactive bands were visualized using enhanced chemiluminescence (Thermo, Pittsburgh, PA, USA) and Image Scanner III (EPSON, Tokyo, Japan).
4.5. RNA Isolation and Analysis
Total RNAs from exosomes and cultured cells were isolated using the mirVana miRNA Isolation Kit (Ambion, Austin, TX, USA) according to the manufacturer’s total RNA isolation procedure. The RNA quality and concentration were assessed with a 260/280 ratio using a NanoDrop spectrophotometer (NanoDrop ND-1000 Technologies, Inc., Wilmington, DE, USA).
4.6. Small RNA Libraries Construction and Solexa Sequencing
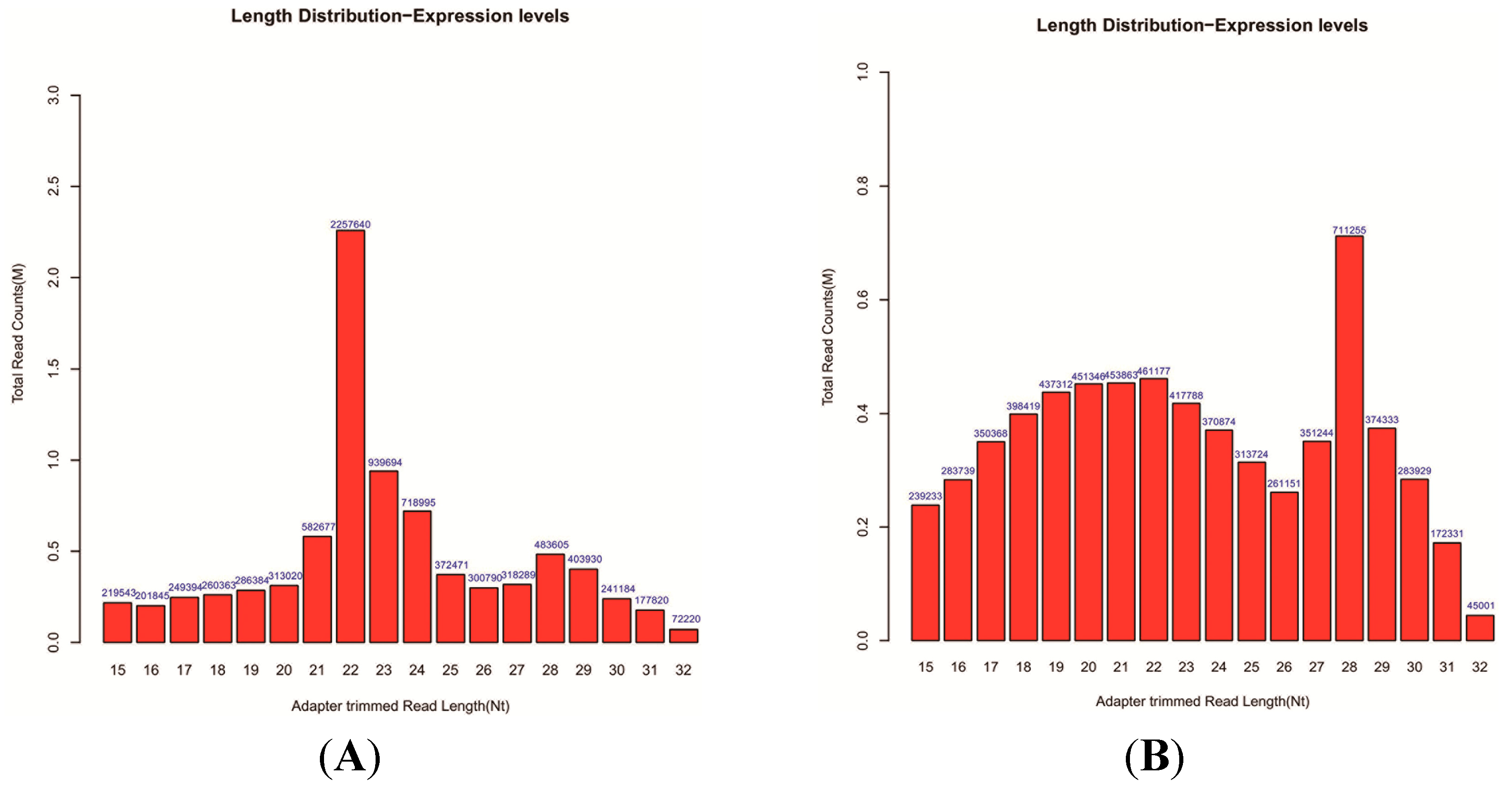
Approximately 1 μg of total RNA from each sample was used to prepare the miRNA sequencing library. The 3'-adapter was ligated to the isolated small RNAs using T4 RNA ligase (Epicentre, San Diego, CA, USA). The 5'-adapter was ligated to the isolated small RNAs using T4 RNA ligase. The adapter-ligated sRNAs were used as templates for RT-PCR to amplify single-stranded cDNA templates to double-stranded cDNAs. Then, ~110–130-bp PCR-amplified fragments (correspond to ~15–32 nt small RNAs) from the 15% denaturation PAGE gels were extracted and then purified. After the completed libraries were quantified using Agilent 2100 Bioanalyzer (Agilent Technologies Inc., Santa Clara, CA, USA), the DNA fragments in the libraries were denatured with 0.1 M NaOH to generate single-stranded DNA molecules, captured using Illumina flow cells, amplified in situ and then finally sequenced for 36 cycles using a Genome Analyzer IIx (Illumina, San Diego, CA, USA) according to the manufacturer’s instructions.
4.8. Expression Patterns of Known and Novel miRNAs
The clean reads were analyzed against the Rfam database using BLAST. The count of clean reads that originated from each miRNA can indicate the expression level of the corresponding miRNA. To understand the differential expression of conserved and novel miRNAs between cells and exosomes, the frequency of each miRNA was normalized to the same order of magnitude using the formula: normalized expression = actual miRNA count/total count of clean reads × 1,000,000 [
20]. If the normalized expression level of an miRNA had read counts <2, this miRNA was removed from future differential expression analyses. Only the perfectly matched sequences were considered to be conserved miRNAs. Furthermore, the prediction of novel miRNAs of cells and exosomes was conducted using miRDeep 2 software.
4.9. miRNA Variants
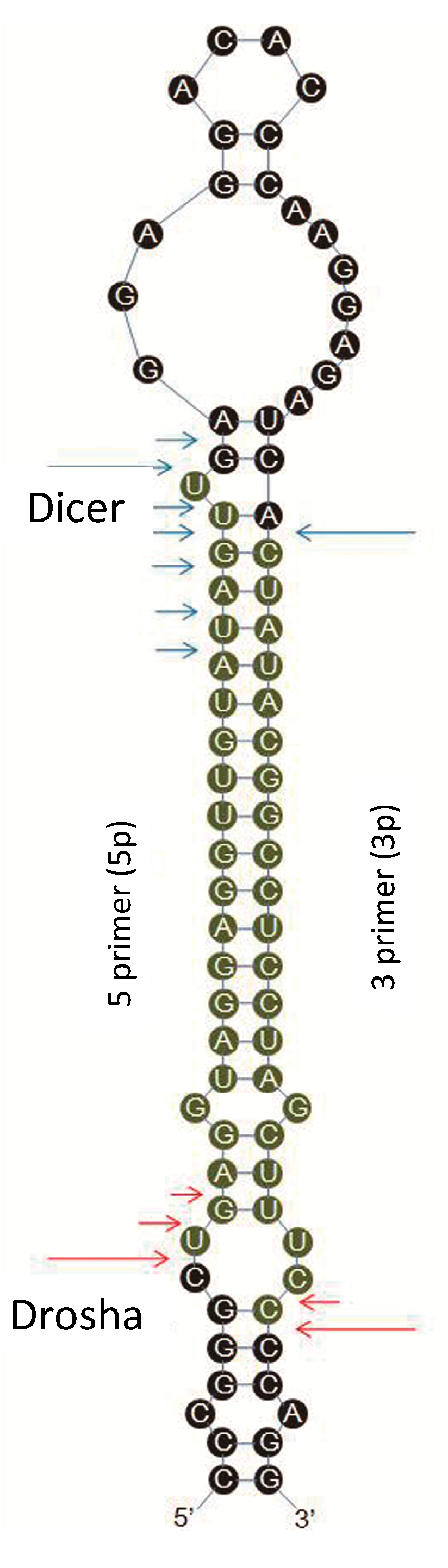
IsomiRs are mature variants of miRNAs from their miRBase reference sequences. The Solexa sequencing results in the present study revealed that most of the identified miRNAs had length and sequence heterogeneity. The considerable degree of isomiR variability can be attributed to the variability in either Dicer or Drosha cleavage positions within the pre-miRNA hairpin (
Figure 9). To characterize the isomiR variability, sequences that matched the miRNA precursors in the mature miRNA region ±4 nt (not more than one mismatch) were considered as mature miRNA isomiRs and were grouped according to the 5' (5p) or 3' (3p) arm of the precursor hairpin.
4.10. Measurement of miRNA Expression Level by qRT-PCR
Total RNA was isolated from exosomes and cultured cells using a mirVana miRNA Isolation Kit (Ambion, Austin, TX, USA). qRT-PCR was performed using SYBR assay according to the manufacturer’s instructions. Each reaction mixture comprised 1 μL of RT-primer, 2.5 μL of 5× reverse-transcription buffer, 1 μL of 2.5 mM dNTPs, 0.25 μL of RNase-Inhibitor, 0.25 μL of MMLV reverse transcriptase (Promega, Madison, WI, USA) and 5 ng of RNA (exosomes) or 500 ng of RNA (cells). ddH
2O was added to a mixed volume of 12.5 μL. The RT reactions were conducted under the following conditions: 42 °C for 60 min, 70 °C for 10 min and then held at 4 °C. For the exosomes, the total RNA content was extremely low. The RT product was pre-amplified prior to the RT-PCR step to enhance sensitivity [
53]. The reaction product was then setup for RT-PCR with the ABI PRISM 7300 real-time PCR system (Applied Biosystems, Carlsbad, CA, USA). Briefly, each reaction was performed with a final volume of 10 μL containing 1 μL of DNA, 3.75 μL of SYBR Green, 0.75 μL of Plus, 1 μL of 5 µM forward primer, 1 μL of 5 μM reverse primer and 2.5 μL of ddH
2O. The reaction mixtures were incubated at 95 °C for 5 min, followed by 40 cycles of 95 °C for 15 s, 60 °C for 30 s and 72 °C for 30 s. Fluorescence measurements were then conducted. To calculate the relative fold change values, the
Ct value data were normalized to U6 and cel-miR-39 as the endogenous controls for cells and exosomes, respectively. The relative quantification (fold change) for the miRNA expression of the host cells and exosomes was determined using 2
−ΔCt (where Δ
Ct = (
Ct of miRNA of interest) − (
Ct of endogenous control gene (U6 or cel-miR-39)). Three technological replicates were used to ensure the reliability of the quantitative analysis. The melting curves in each experiment were analyzed to examine the sensitivity and specificity of the primers.
Figure 9.
The predicted structure of pre-miRNA is presented as a graph. The experimentally inferred Drosha and Dicer cleavage positions are indicated with red and blue arrows, respectively. Long arrows represent cleavage sites for the most abundant isomiR, and short arrows indicate other isomiRs.
Figure 9.
The predicted structure of pre-miRNA is presented as a graph. The experimentally inferred Drosha and Dicer cleavage positions are indicated with red and blue arrows, respectively. Long arrows represent cleavage sites for the most abundant isomiR, and short arrows indicate other isomiRs.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
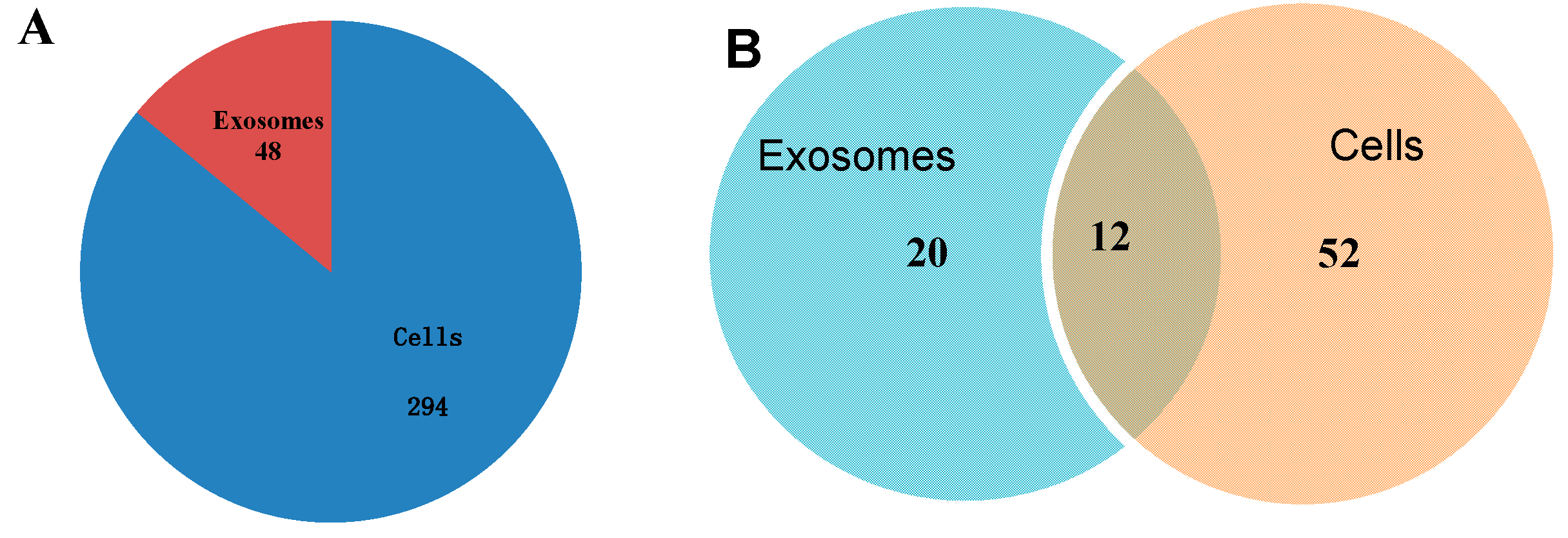{kind=link}
{kind=link}
{kind=link}
{kind=link}







