1. Introduction
Nonalcoholic fatty liver disease (NAFLD) is characterized by excessive lipid accumulation in the liver without chronic alcohol consumption. It encompasses diverse hepatic disorders ranging from hepatic steatosis to nonalcoholic steatohepatitis (NASH), fibrosis, and cirrhosis. The prevalence of NAFLD has been increasing worldwide and coincides with the frequency of metabolic disorders including obesity, type 2 diabetes, and hyperlipidemia. Hepatic steatosis refers to over-accumulation of triglyceride (TG) in hepatocytes. TG accumulation results from increased lipid delivery to the liver, increased
de novo lipogenesis, and reduced fatty acid (FA) oxidation, reduced lipolysis, and decreased lipid secretion [
1,
2]. According to “two-hit” hypothesis, the progression of NAFLD occurs in two steps. The first step is the accumulation of TG in the liver, and the second step involves additional pathophysiological insults, including oxidative stress, inflammation, and cell death [
1,
2].
Alisma orientale Juzepzuk is widely cultivated in China, Japan, and Korea. It has been used in clinical practice for the treatment of edema, dysuria, and diarrhea, and has also been prescribed as an important component of herbal combinational therapy for the treatment of hypertension, hyperlipidemia, and hyperglycemia. Furthermore, many studies have reported that it has therapeutic potential for inflammation [
3], allergy [
4], and oxidative stress [
5]. A recent study has shown that
Alisma orientale has a protective effect against nonalcoholic fatty liver induced by high fat diet (HFD) [
6]. However, the mechanism involved in this protective effect has not been characterized. The chemical constituents of
Alisma orientale include sesquiterpenes, protostane-type triterpenes, and guaiane-type and kaurane-type diterpenes [
7].
The endoplasmic recticulum (ER) is an intracellular organelle that regulates lipid production, protein synthesis for most cellular organelles, and Ca
2+ storage [
8,
9]. Different stimuli that disrupt ER homeostasis increases the accumulation of unfolded proteins in the ER, which leads to ER stress. To solve ER stress, unfolded protein response (UPR) is activated. The UPR attenuates protein translation, degrades unfolded proteins, and increases protein folding capacity of the ER [
10]. Chronic or increased ER stress leads to the pathogenesis of multiple diseases including diabetes [
11]. Recently, it was reported that ER stress is associated with the development of hepatic steatosis [
12]. ER stress disturbs hepatic lipid metabolism by regulating lipogenic gene expression and apolipoprotein secretion, and by promoting insulin resistance. Furthermore, ER stress activates Nrf2, JNK, and NFκ-B pathways, which play important roles in inflammatory process [
13].
Although
Alisma orientale was found to protect against HFD-induced hepatic steatosis in rat [
6], the underlying mechanism was not characterized. Furthermore, it remains unclear whether
Alisma orientale extract can attenuate ER stress, a major contributor of hepatic steatosis. Therefore, this study was designed to investigate the protective effect of
Alisma orientale against ER stress and hepatic steatosis
in vitro and
in vivo. To this end, the experiments were conducted in Human hepatocellular carcinoma (HepG2) cells and mice treated with tunicamycin (pharmacologic ER stress inducer), palmitate-treated HepG2 cells, and HFD obese animals. Furthermore, the molecular mechanisms of protective effects against ER stress-induced hepatic steatosis were characterized.
3. Discussion
Alisma orientale is a well-known Chinese traditional medicine, which exhibits anti-inflammatory and anti-allergic properties [
3,
14]. Chronic ER stress has been reported to induce metabolic diseases including type 2 diabetes, hyperlipidemia, and obesity [
11,
12,
13]. Recently, it was reported that ER stress causes the development of non-alcoholic fatty liver and alcoholic fatty liver by regulating lipid metabolism [
12]. Therefore, a substance that could attenuate ER stress would be a therapeutic candidate for fatty liver disease. Reportedly, tauroursodeoxycholic acid and 4-phenylbutyric acid attenuate ER stress by increasing protein folding and trafficking, and decrease hepatic lipid accumulation in
ob/
ob mice [
21,
22,
23]. Here, we demonstrated that MEAO prominently attenuated ER stress and prevented the development of the hepatic steatosis induced by ER stress.
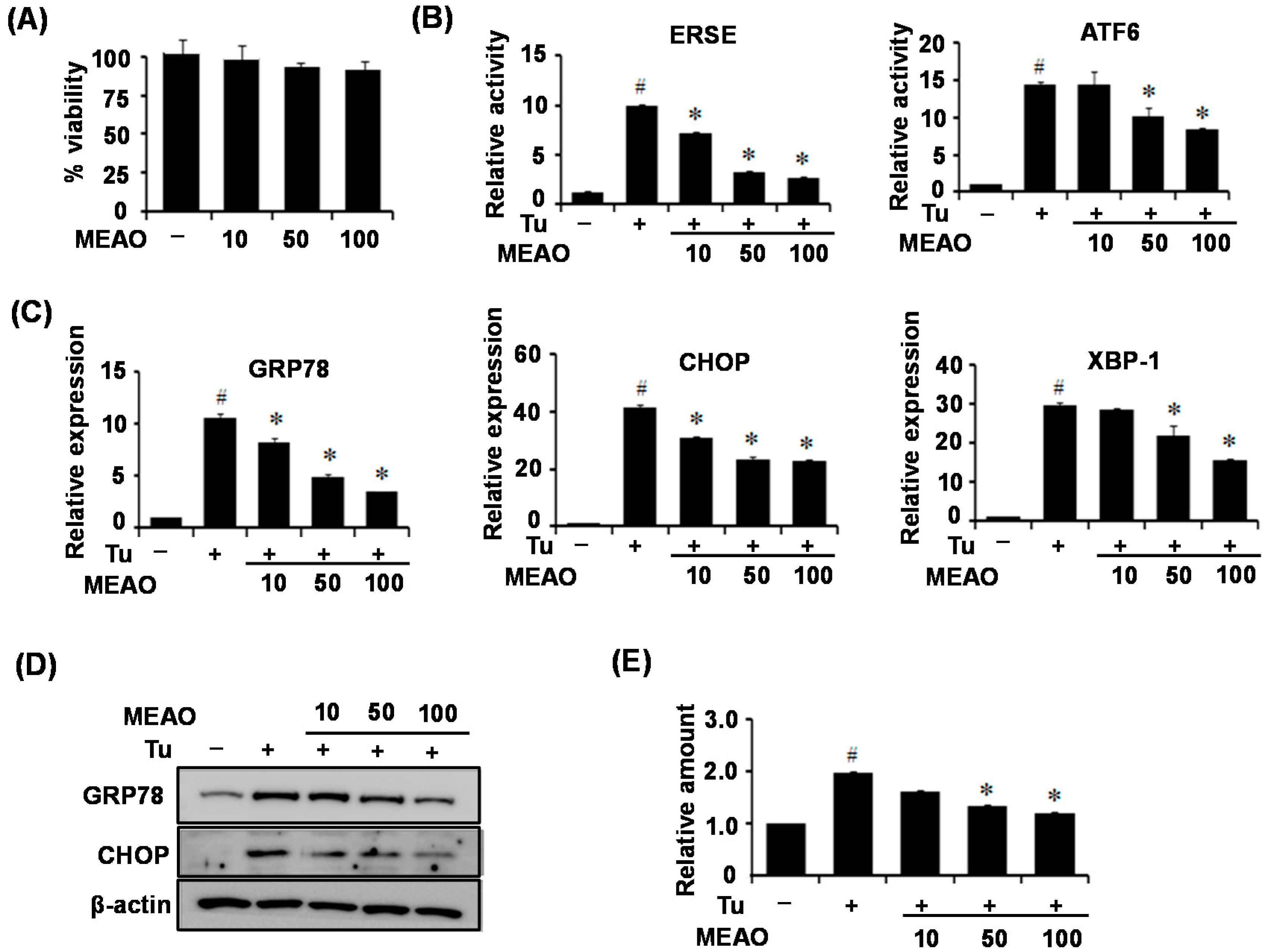
Alisma orientale has been proved to display positive pharmacological effects against several diseases. However, to our knowledge no study has evaluated its protective effects against ER stress, which is a major pathogenic factor for several diseases including hepatic steatosis. Therefore, in this study, we investigated the protective effects of MEAO against ER stress, and determined whether MEAO could ameliorate ER stress-induced hepatic steatosis. We first determined the inhibitory activity of MEAO on ER stress reporters including ER stress response element or ATF6 response element. MEAO efficiently inhibited the tunicamycin-induced increase in luciferase activity of the reporters. Then, the protective effects against ER stress and ER stress-induced hepatic steatosis were examined in vitro and in vivo. For the in vitro study, HepG2 cells were treated with tunicamycin in the presence of MEAO, and ER stress markers and cellular triglyceride levels were measured in the cell extracts. MEAO greatly attenuated the tunicamycin-induced increases in both ER stress marker and cellular triglyceride levels. Similarly, for the in vivo study, C57BL6J mice were administered MEAO for 2 days, and injected with tunicamycin for 24 h. Tunicamycin treatment increased the levels of ER stress marker proteins and triglyceride accumulation in the livers, both of which decreased in MEAO-administered mice. Taken together, the in vitro and in vivo study results indicate that MEAO attenuated ER stress and improved ER stress-induced hepatic steatosis.
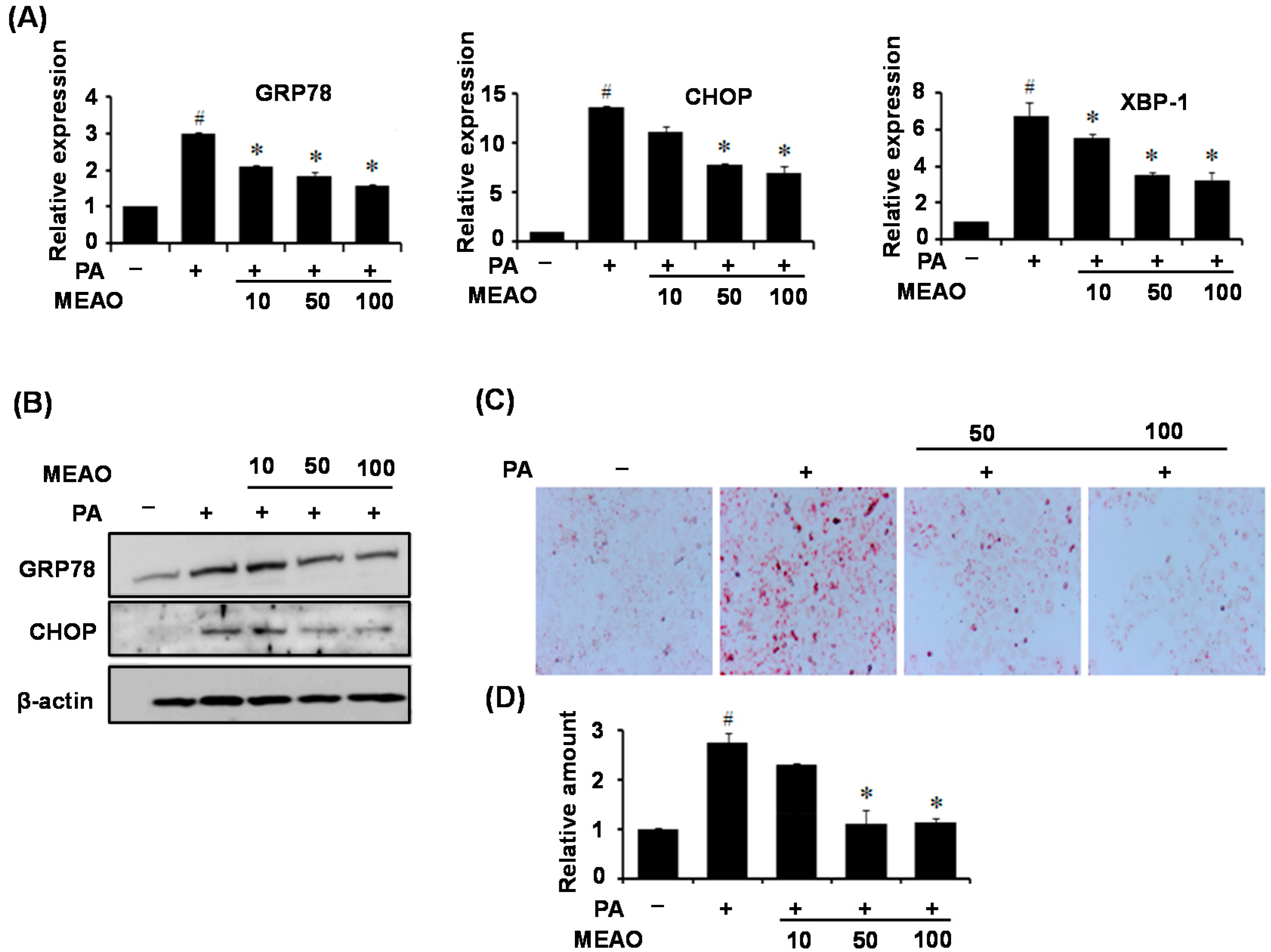
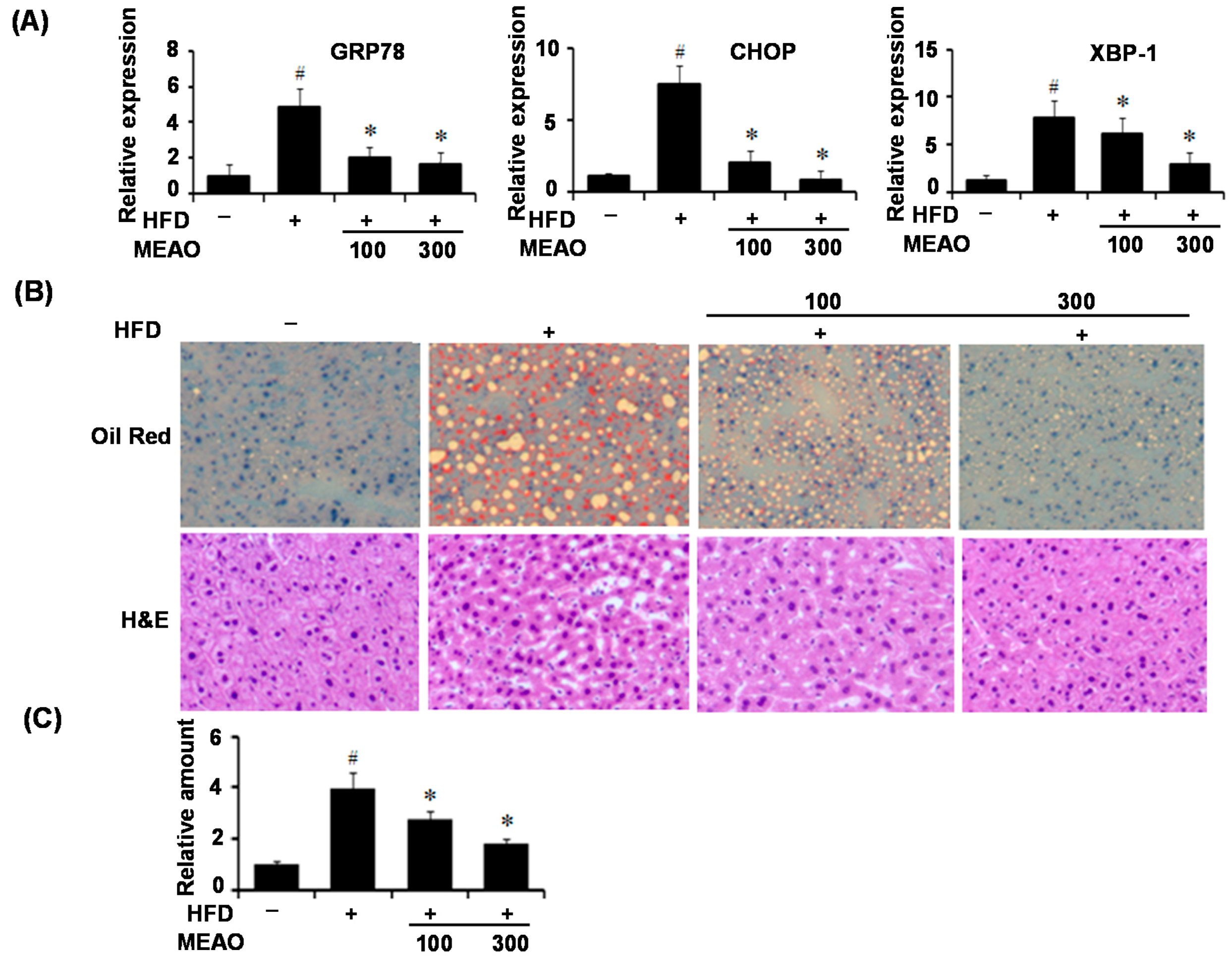
We then confirmed the protective effects of MEAO in fatty acid-treated HepG2 cells and HFD obese mice. PA treatment induced ER stress in HepG2 cells as revealed by increase in ER stress markers, and increased cellular triglyceride levels. HFD also induced hepatic ER stress and increased hepatic lipid accumulation; however, MEAO treatment efficiently blocked the increase in ER stress markers and lipid accumulation. These results demonstrate that MEAO ameliorated hepatic steatosis through the inhibition of ER stress.
The ER elicits an elaborative adaptive response under pathological and/or stressful conditions, collectively known as the UPR [
8,
9]. The three branches of the UPR regulate the transcription of various genes involved in the expansion of ER, decrease of protein synthesis, increase of protein folding capability and degradation of terminally unfolded and misfolded proteins within the ER. Previous study reported that IRE1α contributed to the development of hepatic steatosis through upregulation of gene expression involved in fatty acid and triglyceride synthesis by XBP-1 [
14]. PERK also increased the expression of the lipogenic enzymes such as FAS, ATP citrate lyase, and stearoyl-CoA desaturase-1 in the mouse mammary gland [
15]. Furthermore, CHOP deficiency in mice inhibited cholestasis induced liver fibrosis [
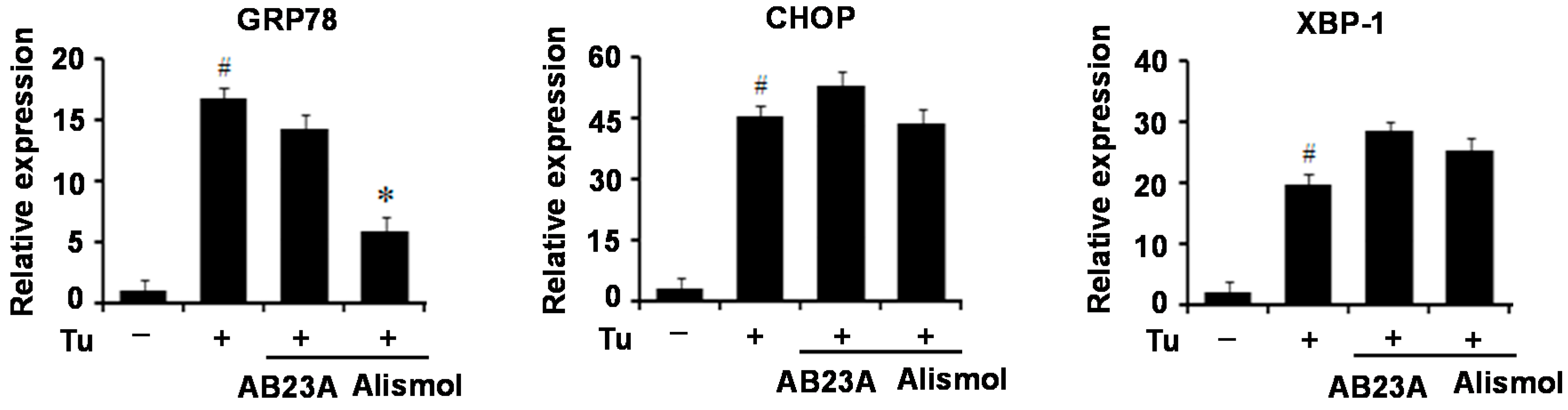
24]. According to our current results, MEAO blocked the increase in GRP78, CHOP and XBP-1x splicing respectively, suggesting that MEAO potentially acts on hepatic steatosis via inhibition of the three branches of UPR.
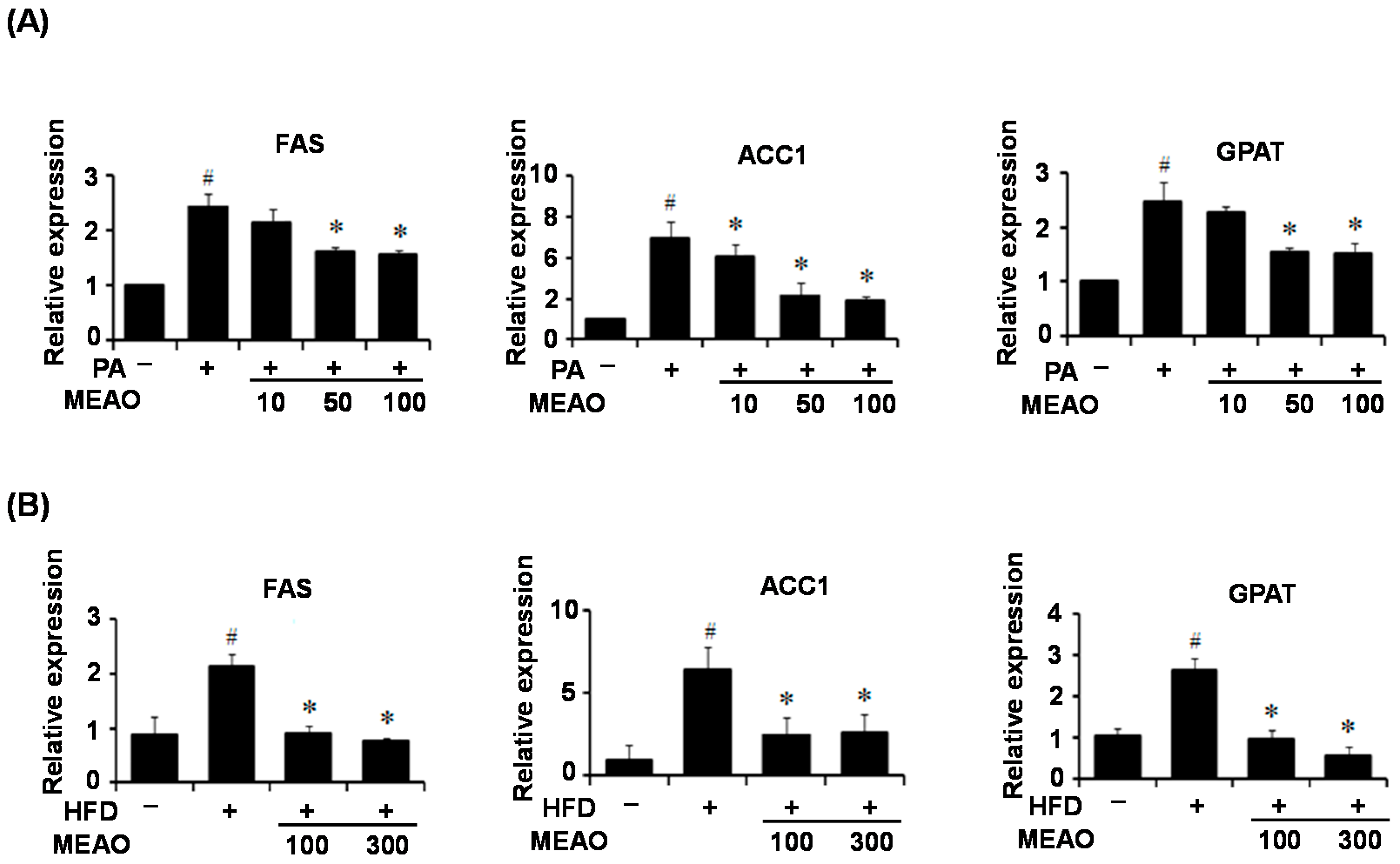
To investigate the mechanism involved in ER stress-mediated inhibition of lipid accumulation, we first examined the expression of lipogenic genes in the livers of tunicamycin-injected mice because ER stress regulates lipid metabolism-genes. Consistent with previous reports [
16,
17], tunicamycin treatment decreased the expression of lipogenic genes including FAS, ACC1, and GAPT despite the presence of hepatic steatosis. However, this was not significantly affected by MEAO. These results suggest that the rescue of tunicamycin-induced hepatic steatosis by MEAO was not due to the inhibition of lipogenesis. However, the increased expression of lipogenic genes in PA-treated HepG2 cells and in the liver of HFD obese mice was inhibited by MEAO, suggesting that MEAO potentially ameliorates fatty acid-induced hepatic steatosis through inhibition of lipogenesis.
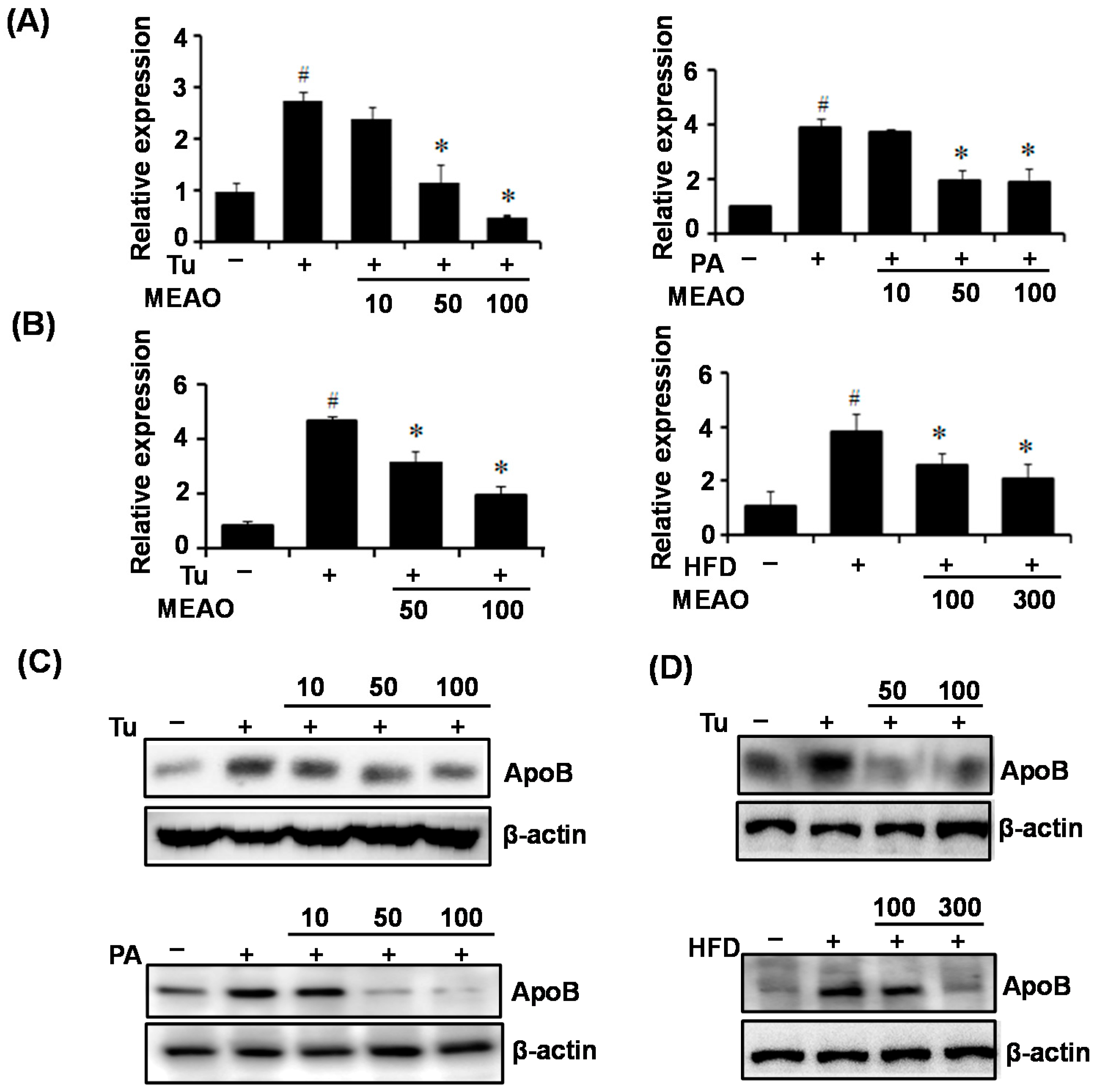
Furthermore, it was known that VLDLR mediates APOE-containg VLDL uptake into hepatic cells, and increased intracellular triglyceride accumulation. ApoB is a secretary protein involved in the transfer of lipids out of hepatic cells. ER stress has been shown to increase VLDLR expression and inhibit ApoB secretion, which might be a primary mechanism of tunicamycin-induced hepatic steatosis under conditions of suppressed lipogenesis [
17,
18]. Therefore, we investigated VLDLR expression and ApoB secretion in tunicamycin-treated HepG2 cells and mice. Tunicamycin treatment increased VLDL expression and intracellular ApoB protein levels in the livers of mice as well as in HepG2 cells. However, the increases in VLDL expression and intracellular ApoB levels were inhibited by MEAO. The suppressive effects of MEAO on VLDL expression and intracellular ApoB levels were also observed in PA-incubated HepG2 cells and in the livers of HFD obese mice, indicating that MEAO suppresses VLDLR expression, decreases uptake of APOE-containing VLDL, and also inhibits ApoB secretion, which might cause the attenuation of triglyceride accumulation. Together, these results suggest that MEAO attenuated ER stress and ameliorated hepatic steatosis via the downregulation of VLDLR expression, and the improvement of ApoB secretion under conditions of suppressed lipogenesis.
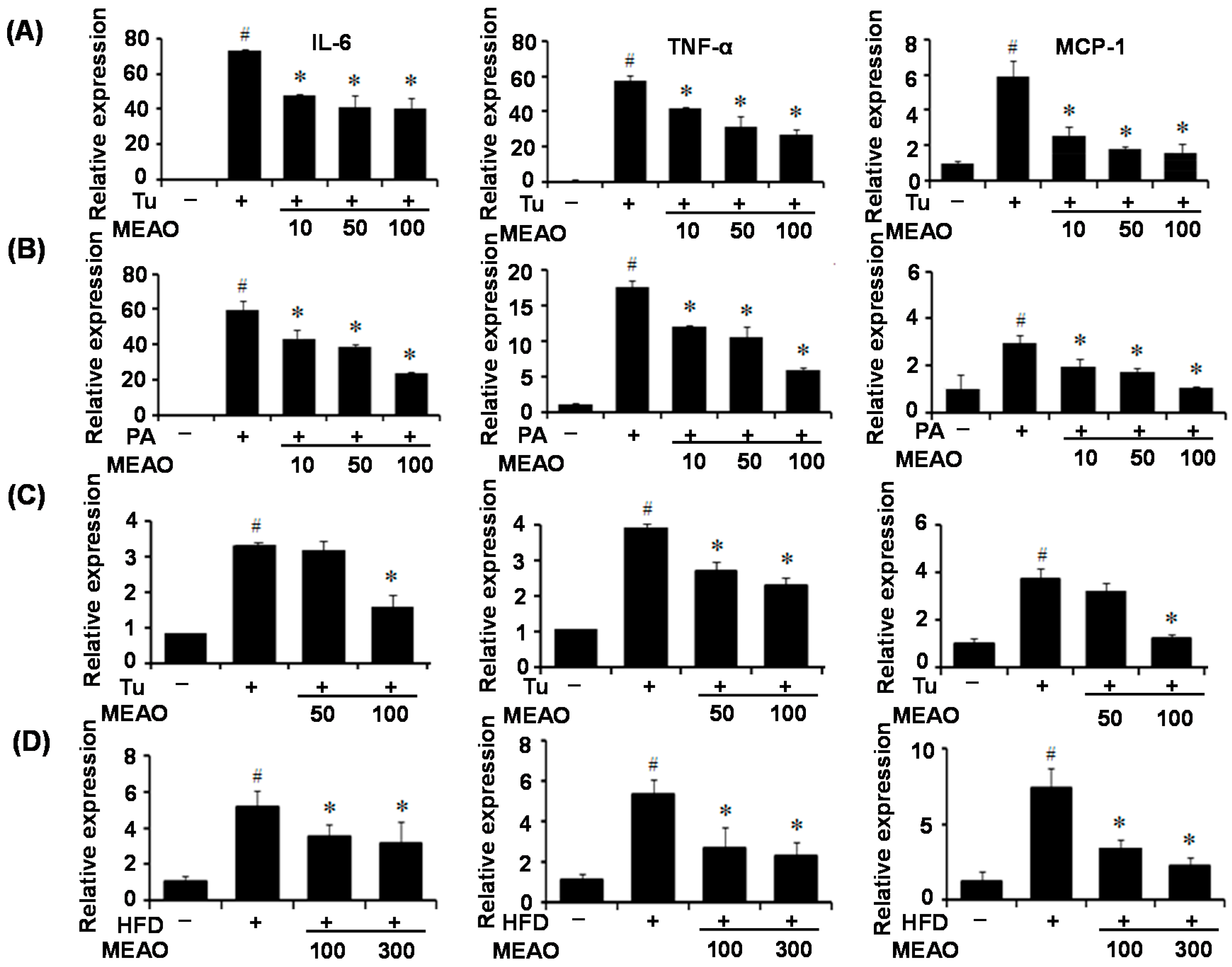
Inflammation has been demonstrated to be an important mechanism of hepatic steatosis and to exacerbate liver injury and NASH [
1,
2]. In addition, it has been reported that ER stress causes inflammation [
13]. Therefore, we investigated whether MEAO alleviates hepatic ER stress-associated inflammation
in vitro and
in vivo. Pro-inflammatory cytokines, such as IL-6, TNF-α, and MCP1 were induced by treatment with tunicamycin and PA. However, MEAO prevented the increase of inflammatory cytokine expression. These results indicate that MEAO ameliorated the ER stress induced-inflammation associated with hepatic steatosis.
Terpenoid is a main chemical component of
Alisma orientale along with protostane-type triterpenes, guaiane-type sesquiterpenes, and kauranetype diterpenes [
20]. Many terpenes has been isolated from
Alisma orientale [
25,
26] which were known to have various functions including anti-type I–IV allergic response activity, anti-inflammatory activity, anti-hepatitis B virus activity, anti-tumor activity, and anti-complementarity activity [
3,
4,
5]. Among these components, alismol is a guaiane-type sesquiterpenes that has been shown to inhibit nitric oxide synthesis in RAW 264.7 cells stimulated with interferon-gamma plus lipopolysaccharide and to suppress mainly Ca
2+ influx through a voltage-dependent Ca
2+ channel [
27,
28]. In this study, alismol significantly blocked the increase in GRP78 expression mediated by tunicamycin, whereas it did not affect the levels of CHOP and XBP-1c, suggesting that alismol might represent one of the active components of MEAO responsible for anti-ER stress and hepatic steatosis which might exert a protective effect against ER stress through the inhibition of ATF6-GRP78 pathway among the three branches of UPR.
In conclusion, MEAO efficiently attenuated ER stress and prevented the development of hepatic steatosis through inhibition of the expression of hepatic lipogenic genes and of VLDLR, and enhancement of ApoB secretion. Our experimental evidence supports the use of MEAO as a potential therapeutic agent for the treatment of ER stress-induced diseases.
4. Experimental Section
4.1. Reagents
Tunicamycin and palmitate were purchased from Sigma-Aldrich (St. Louis, MO, USA). Alismol and alisol B23-acetate were purchased from Core Science (Seoul, Korea). Antibodies against GRP78 and CHOP were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibody against ApoB 100 was purchased from Cell Signaling Technology (Beverly, MA, USA).
4.2. Cell Lines
Human hepatocellular carcinoma cells (HepG2) were purchased from the American Type Culture Collection (Manassas, VA, USA). HepG2 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing glucose (Invitrogen, Carlsbad, CA, USA) supplemented with 10% (v/v) fetal bovine serum (Invitrogen).
4.3. Preparation of Alisma Orientale Extract
Dried tubers of Alisma orientale Juzepzuk used in this study was purchased from Naemomea Dah Corporation (Ulsan, Korea, in July 2014), and was identified by Professor Ki-Tae Ha, School of Korean Medicine, Pusan National University (Korea). The whole plant voucher specimen (number: pnuhc001) was registered and deposited at the School of Korean Medicine, Pusan National University. Dried specimen (400 g) was ground into power, and extracted three times with methanol (1 L × 3) for 3 h at room temperature, and filtered through a filter paper. The filtrate was evaporated in vacuo at 40 °C to produce a methanol extract of Alisma orientale (MEAO, 58.76 g). Appropriate amount of the extract was dissolved in dimethyl sulfoxide (DMSO) prior to experiment.
4.4. Cell Viability Assay
Cell viability was measured by a quantitative colorimetric assay with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoli-um bromide (MTT). HepG2 cells were plated in 96-well plates at a density of 5000 cells/well and then cultured for 24 h. After incubation the cells with various concentrations of MEAO (10, 50, and 100 µg/mL) for 16 h, MTT solution (2.0 mg/mL) was added to each well for 4 h. DMAO was added to each well after removing the media, and the optical density (OD) was measured at 540 nm with a microplate reader.
4.5. Reporter Assay
To determine the protective activity against ER stress, HepG2 cells (2 × 104 cells/well in a 96-well plate) were transfected with an ERSE-Luc reporter or ATF6-Luc reporter using a Cignal™ Reporter Assay kit (Qiagen, Valencia, CA, USA). The ERSE-Luc and ATF6-Luc were purchased from Addgene (Cambridge, MA, USA). The consensus ERSE is 5′-CCAAT(N9)CCACG and ATF6 sequence is 5′-TCGAGACAGGTGCTGACGTGGCGATTCC. The cells were then incubated with tunicamycin (2 μg/mL) in the presence of the MEAO and then the luciferase activities were determined with a Dual-Glo Luciferase assay system kit (Promega, Madison, WI, USA).
4.6. Animal Experiments
C57BL/6 mice were purchased from Central Lab. Animal Inc. (Seoul, Korea). The animals were maintained on a 12 h light/dark cycle under 21–23 °C of temperature and 40%–60% of humidity. For the tunicamycin-induced ER stress animal model, mice at 8 weeks of age were randomly divided into four groups (n = 7) with equal mean body weights. The mice were administrated MEAO (50 mg/kg body weight and 100 mg/kg body weight) for 2 days and then injected intraperitoneally with DMSO or tunicamycin at a dose of 1 mg/kg body weight for 24 h. Then, the mice were administrated with MEAO for additional 2 days and sacrificed. For high fat diet (HFD) obese mice, the mice at 6 weeks of age were randomly divided into four groups (n = 7) with equal mean body weights: lean control (distilled water-treated) group, high-fat diet (distilled water-treated) group, high-fat diet and MEAO (100 mg/kg/day of body weight) group, high-fat diet and MEAO (300 mg/kg/day of body weight) group. The experimental diets were the AIN93G-based on the High-fat diet containing 60% kcal fat (37.1% of saturated fat) and the control diet containing 10% kcal fat. MEAO was administered orally daily for 16 weeks. All animal experiments were approved by the Pusan National University Animal Experiment Ethics Committee and were conducted according to the institutional guidelines for the care and use of laboratory animals.
4.7. Histological Analysis
Liver specimens were fixed in 10% formalin and embedded in paraffin, and cutted into 3 µm sections. The liver slides were washed with 60% isopropanol for 5 min and stained with Oil-red O working solution (1.5 mg/mL Oil-red O/60% isopropanol) for 15 min at room temperature. The slides were washed with distilled water and photographed under a light microscope. For hematoxylin and eosin (H&E) staining, the liver slides were stained with hematoxylin and eosin, and the histopathology was observed under a microscope.
4.8. Measurement of Triglyceride Level
Liver tissues and HepG2 cells were homogenized in chloroform-methanol solution (2:1, v/v) and choloroform-methanol-H2O solution (8:4:3, v/v), respectively. The homogenates were incubated at room temperature for 1 h and centrifuged at 800× g for 10 min. The obtained bottom layer (organic phage) was dried overnight. After dissolving the dried bottom layer in ethanol, triglyceride concentrations were determined using an enzyme reaction kit (Asan pharmaceutical, Seoul, Korea) and normalized to the protein concentration.
4.9. Quantitative Polymerase Chain Reaction (q-PCR)
RNA was isolated from the livers of the mice and HepG2 cells with Trizol™ (Invitrogen). The cDNA was generated from 1 μg of total RNA using GoScript™ Reverse Transcription System (Promega, Madison, WI, USA) following the manufacturer’s protocol. The primers used in this study are listed (
Table S1).
4.10. Western Blot
Equal amounts of protein (40 μg/lane) from liver homogenates and HepG2 cell lysates were resolved by 8% SDS polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluroide membranes (Millipore, Massachusetts, MA, USA). The levels of GRP78, CHOP, and actin were detected using an enhanced chemiluminescence western blot detection kit (Amersham, Uppsala, Sweden).
4.11. Statistical Analysis
All data are presented as mean ± SEM. Statistical comparisons were analyzed using Student’ t-test and one-way analysis of variance (ANOVA), followed by Tukey’s test. Values were considered statistically significant at p < 0.05.



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







