
Resveratrol Protects against Helicobacter pylori-Associated Gastritis by Combating Oxidative Stress

Abstract
:1. Introduction
2. Results and Discussion
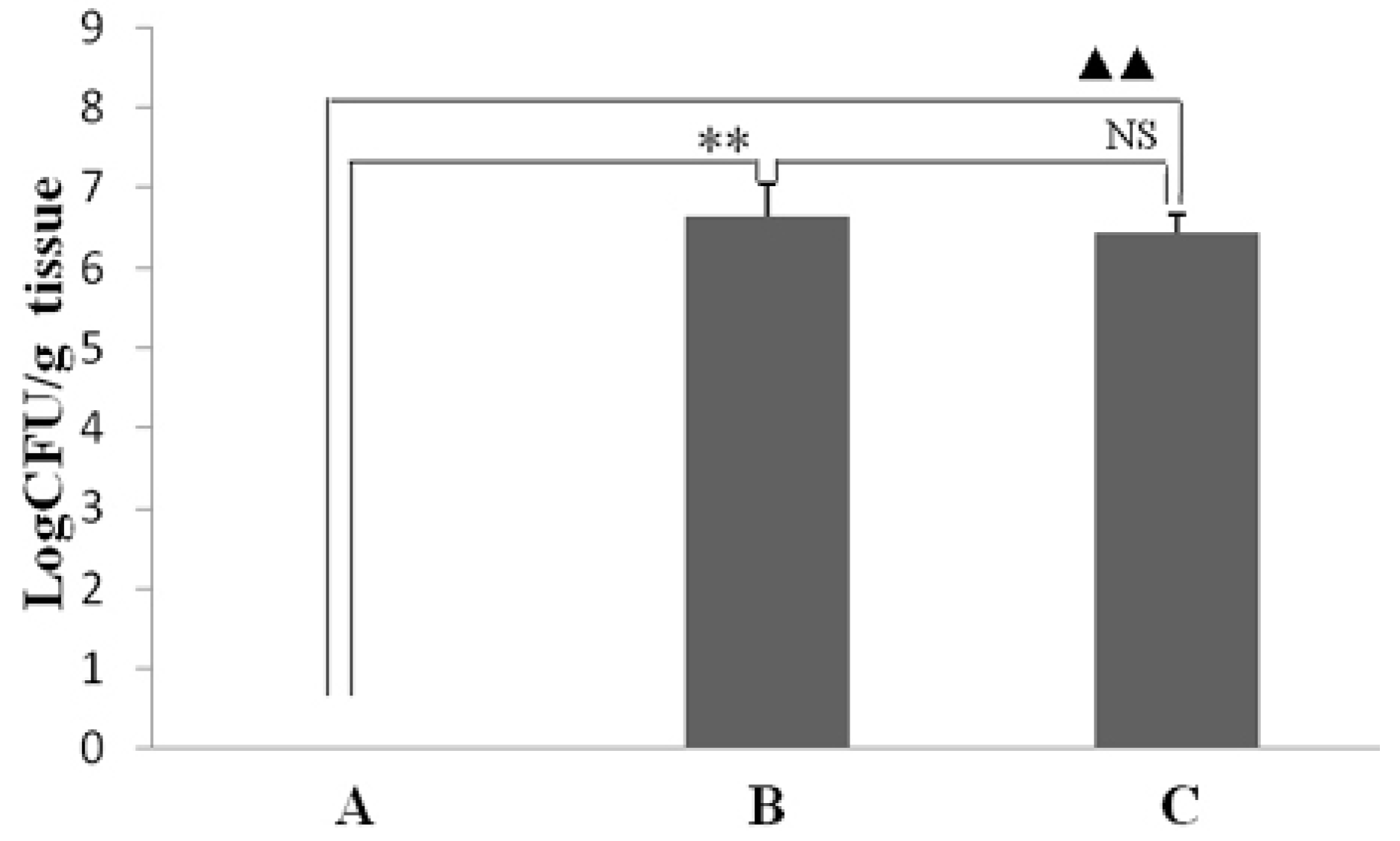
2.1. Effect of Resveratrol on the Colonization Number of H. pylori in the Stomach

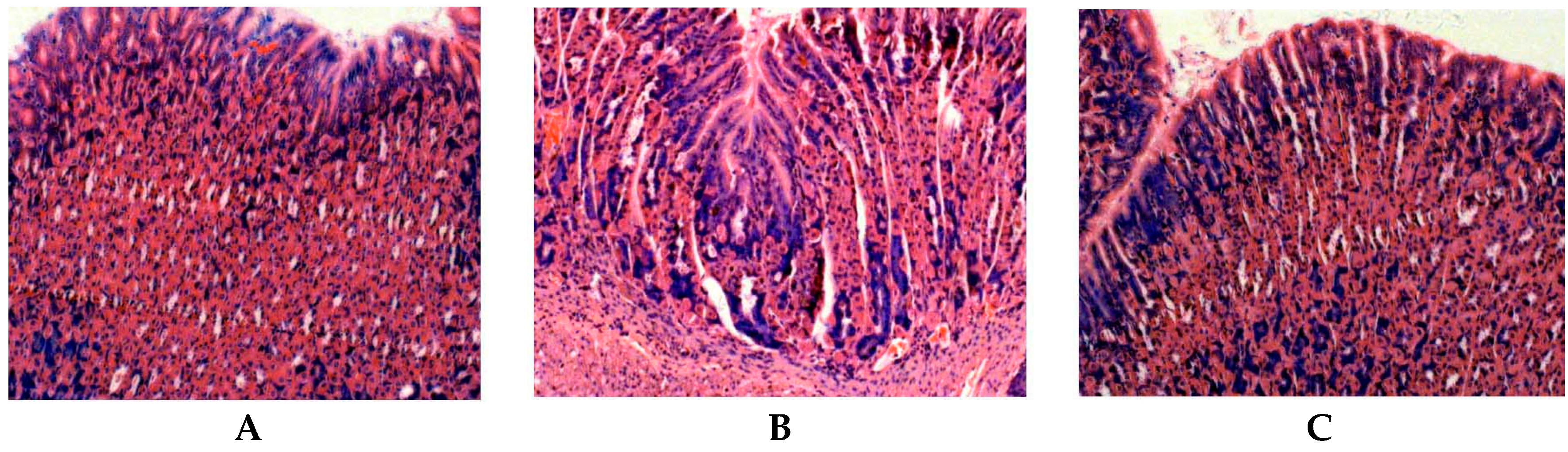
2.2. Effect of Resveratrol on H. pylori-Caused Gastric Tissue Inflammation
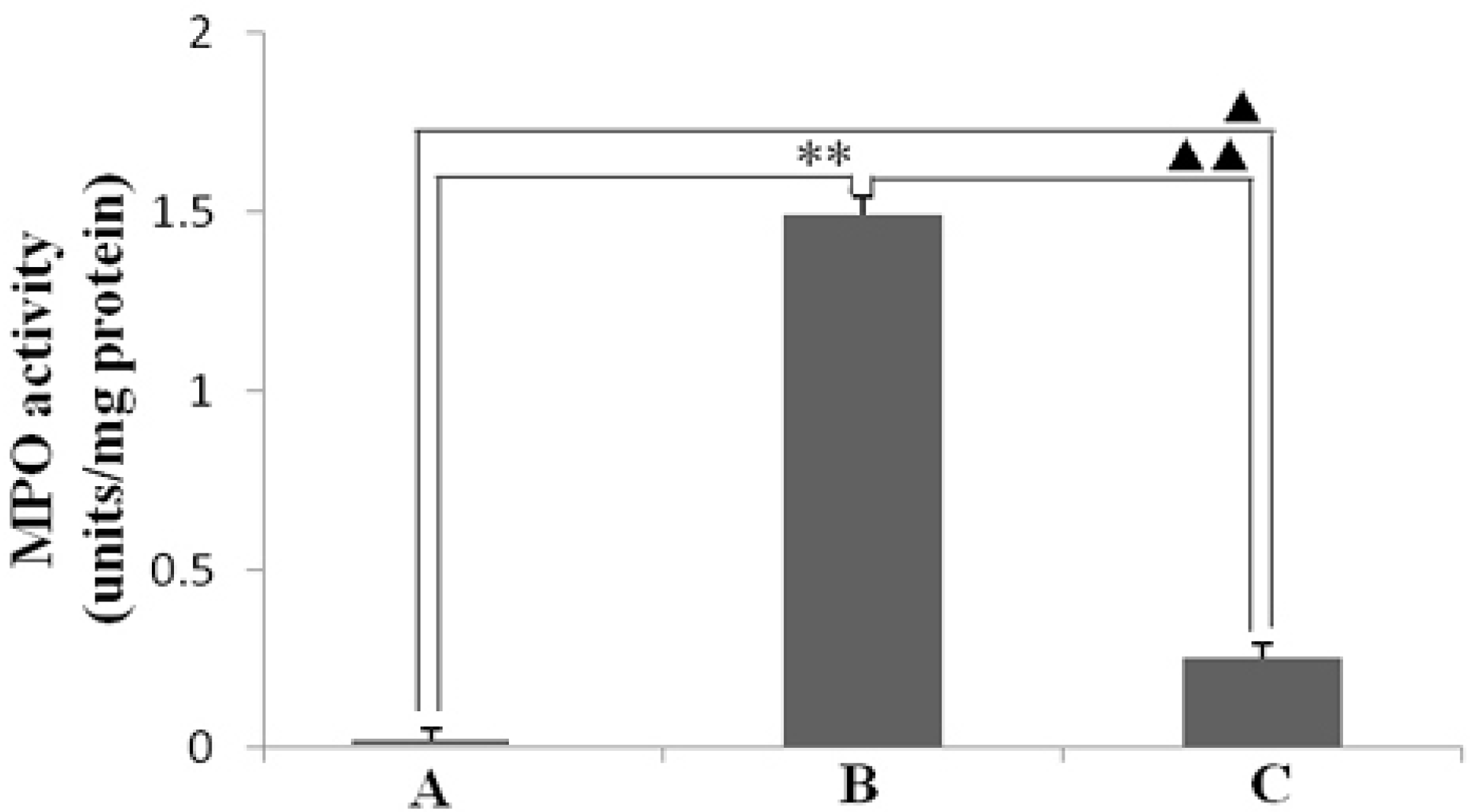
2.3. Effect of Resveratrol on H. pylori-Caused MPO Activity and LPO Level



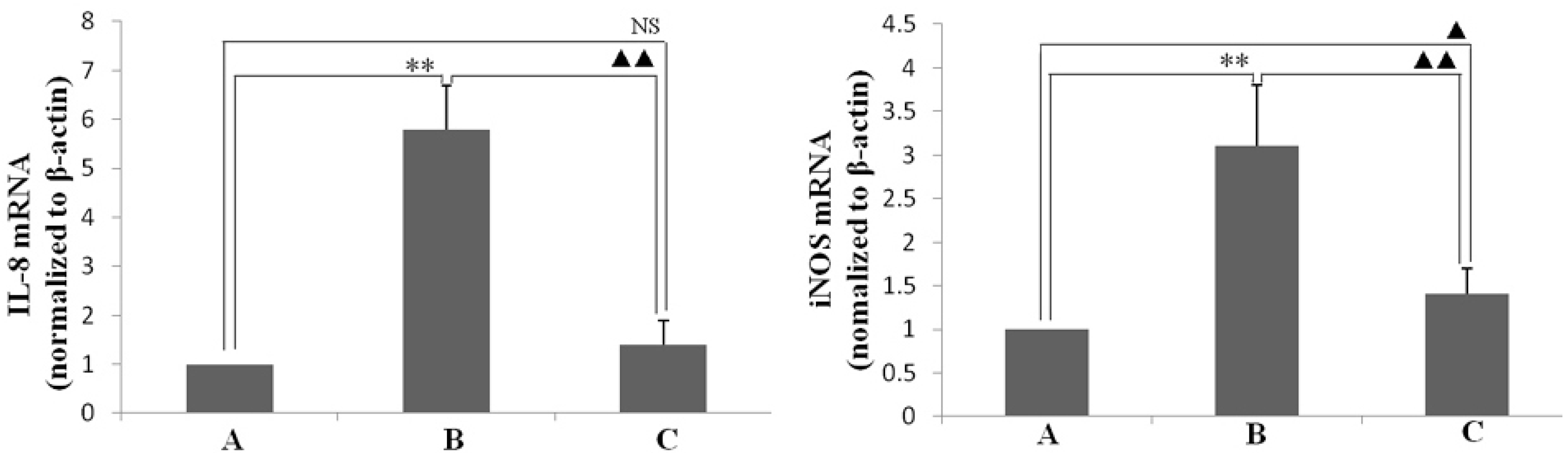
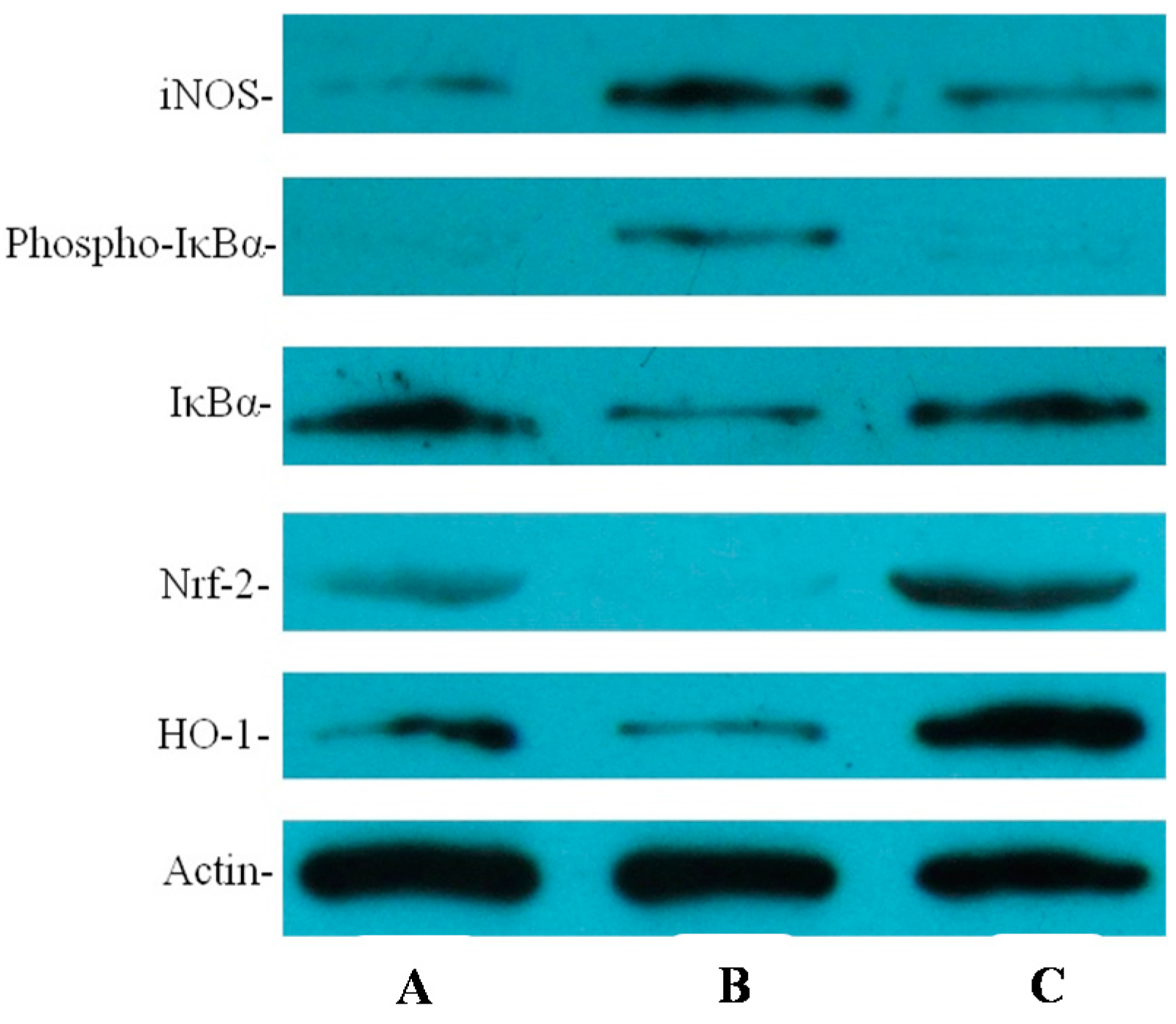
2.4. Effect of Resveratrol on H. pylori-Caused Expression of IL-8 and iNOS, HO-1 and Nrf2, Level of Phosphorylation of IκBa



3. Experimental Section
3.1. Animals and Resveratrol
3.2. Growth of H. pylori and Inoculation of Mice
3.3. Experimental Design
3.4. Determination of H. pylori Colonization in Mouse Stomach
3.5. Histological Observation and Grade
3.6. Determination of LPO Level and MPO Activity
3.7. Quantitative RT-PCR to Assess the Expression Level of iNOS and IL-8 mRNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Base Pair (bp) |
|---|---|---|
| iNOS | Forward-5′-ACAATACAAGATGACCCTA-3′ | 357 |
| Reverse-5′-CAGATGTTCCTCTATTTTT-3′ | ||
| IL-8 | Forward-5′-ATGGCTGCTCAAGGCTGGTCC-3′ | 389 |
| Reverse-5′-ATTCTCTTGTTCTCAGGTC-3′ | ||
| β-Actin | Forward-5′-ATGGTGGGAATGGGTCAGA-3′ | 391 |
| Reverse-5′-CGTGAGGGAGAGCATAGCC-3′ |
3.8. ELISA Assay for IL-8
3.9. Western Blot Analyses for the Levels of iNOS, HO-1, Nrf2 and IκBα, the Degree of IκBα Phosphorylated
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Blaser, M.J. Helicobacter pylori and the pathogenesis of gastroduodenal inflammation. J. Infect. Dis. 1990, 161, 626–633. [Google Scholar] [CrossRef] [PubMed]
- NIH Consensus Conference. Helicobacter pylori in peptic ulcer disease. NIH consensus development panel on Helicobacter pylori in peptic ulcer disease. J. Am. Med. Assoc. 1994, 272, 65–69. [Google Scholar] [CrossRef]
- Amagase, K.; Nakamura, E.; Endo, T.; Hayashi, S.; Hasumura, M.; Uneyama, H.; Torii, K.; Takeuchi, K. New frontiers in gut nutrient sensor research: Prophylactic effect of glutamine against Helicobacter pylori-induced gastric diseases in Mongolian gerbils. J. Pharmacol. Sci. 2010, 112, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Handa, O.; Naito, Y.; Yoshikawa, T. Helicobacter pylori: A ROS-inducing bacterial species in the stomach. Inflamm. Res. 2010, 59, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Kaise, M.; Miwa, J.; Iihara, K.; Suzuki, N.; Oda, Y.; Ohta, Y. Helicobacter pylori stimulates inducible nitric oxide synthase in diverse topographical patterns in various gastroduodenal disorders. Dig. Dis. Sci. 2003, 48, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Rieder, G.; Hofmann, J.A.; Hatz, R.A.; Stolte, M.; Enders, G.A. Up-regulation of inducible nitric oxide synthase in Helicobacter pylori-associated gastritis may represent an increased risk factor to develop gastric carcinoma of the intestinal type. Int. J. Med. Microbiol. 2003, 293, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lim, J.W.; Kim, K.H. Helicobacter pylori-induced expression of interleukin-8 and cyclooxygenase-2 in AGS gastric epithelial cells: Mediation by nuclear factor-κB. Scand. J. Gastroenterol. 2001, 36, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Seo, J.Y.; Kim, K.H. Inhibition of lipid peroxidation, NF-κB activation and IL-8 production by rebamipide in Helicobacter pylori-stimulated gastric epithelial cells. Dig. Dis. Sci. 2000, 45, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.W.; Kim, H.; Kim, K.H. NF-κB, inducible nitric oxide synthase and apoptosis by Helicobacter pylori infection. Free Radic. Biol. Med. 2001, 31, 355–366. [Google Scholar] [CrossRef]
- Fu, S.; Ramanujam, K.S.; Wong, A.; Fantry, G.T.; Drachenberg, C.B.; James, S.P.; Meltzer, S.J.; Wilson, K.T. Increased expression and cellular localization of inducible nitric oxide synthase and cyclooxygenase 2 in Helicobacter pylori gastritis. Gastroenterology 1999, 116, 1319–1329. [Google Scholar] [CrossRef]
- Toyoda, T.; Tsukamoto, T.; Takasu, S.; Shi, L.; Hirano, N.; Ban, H.; Meltzer, S.J.; Wilson, K.T. Anti-inflammatory effects of caffeic acid phenethyl ester (CAPE), a nuclear factor-κB inhibitor, on Helicobacter pylori-induced gastritis in Mongolian gerbils. Int. J. Cancer 2009, 125, 1786–1795. [Google Scholar] [CrossRef] [PubMed]
- Baeuerle, P.A.; Baltimore, D. NF-κB: Ten years after. Cell 1996, 87, 13–20. [Google Scholar] [CrossRef]
- Thanos, D.; Maniatis, T. NF-κB: A lesson in family values. Cell 1995, 80, 529–532. [Google Scholar] [CrossRef]
- Karin, M.; Greten, F.R. NF-κB: Linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol. 2005, 5, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Miura, S.; Mori, M.; Kai, A.; Suzuki, H.; Fukumura, D.; Suematsu, M.; Tsuchiya, M. Rebamipide, a novel antiulcer agent, attenuates Helicobacter pylori induced gastric mucosal cell injury associated with neutrophil derived oxidants. Gut 1994, 35, 1375–1378. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, S.J. Myeloperoxidase: Friend and foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Takagi, T.; Uchiyama, K.; Yoshikawa, T. Heme oxygenase-1: A novel therapeutic target for gastrointestinal diseases. J. Clin. Biochem. Nutr. 2011, 48, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Takagi, T.; Yoshikawa, T. Heme oxygenase-1: A new therapeutic target for inflammatory bowel disease. Aliment. Pharmacol Ther. 2004, 20 (Suppl. S1), 177–184. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Kundu, J.K.; Na, H.K. Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals. Planta Med. 2008, 74, 1526–1539. [Google Scholar] [CrossRef] [PubMed]
- Alam, J.; Cook, J.L. Transcriptional regulation of the heme oxygenase-1 gene via the stress response element pathway. Curr. Pharm. Des. 2003, 9, 2499–2511. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Das, D.K. Anti-inflammatory responses of resveratrol. Inflamm. Allergy Drug Target 2007, 6, 168–173. [Google Scholar] [CrossRef]
- Rubiolo, J.A.; Mithieux, G.; Vega, F.V. Resveratrol protects primary rat hepatocytes against oxidative stress damage: Activation of the Nrf2 transcription factor and augmented activities of antioxidant enzymes. Eur. J. Pharmacol. 2008, 591, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Fauconneau, B.; Waffo-Teguo, P.; Huguet, F.; Barrier, L.; Decendit, A.; Merillon, J.M. Comparative study of radical scavenger and antioxidant properties of phenolic compounds from Vitis vinifera cell cultures using in vitro tests. Life Sci. 1997, 61, 2103–2110. [Google Scholar] [CrossRef]
- Jang, D.S.; Kang, B.S.; Ryu, S.Y.; Chang, I.M.; Min, K.R.; Kim, Y. Inhibitory effects of resveratrol analogs on unopsonized zymosan-induced oxygen radical production. Biochem. Pharmacol. 1999, 57, 705–712. [Google Scholar] [CrossRef]
- Saiko, P.; Szakmary, A.; Jaeger, W.; Szekeres, T. Resveratrol and its analogs: Defense against cancer, coronary disease and neurodegenerative maladies or just a fad? Mutat. Res. 2008, 658, 68–94. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.; Shishodia, S.; Takada, Y. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer Res. 2004, 24, 2783–2840. [Google Scholar] [PubMed]
- Daroch, F.; Hoeneisen, M.; González, C.L.; Kawaguchi, F.; Salgado, F.; Solar, H.; García, A. In vitro antibacterial activity of Chilean red wines against Helicobacter pylori. Microbios 2001, 104, 79–85. [Google Scholar] [PubMed]
- Martini, S.; Bonechi, C.; Rossi, C.; Figura, N. Increased susceptibility to resveratrol of Helicobacter pylori strains isolated from patients with gastric carcinoma. J. Nat. Prod. 2011, 74, 2257–2260. [Google Scholar] [CrossRef] [PubMed]
- Paulo, L.; Oleastro, M.; Gallardo, E.; Queiroz, J.A.; Domingues, F. Anti-Helicobacter pylori and urease inhibitory activities of resveratrol and red wine. Food. Res. Int. 2011, 44, 964–969. [Google Scholar] [CrossRef]
- Smoot, D.T.; Elliott, T.B.; Verspaget, H.W.; Jones, D.; Allen, C.R.; Vernon, K.G.; Bremner, T.; Kidd, L.C.; Kim, K.S.; Groupman, J.D.; et al. Influence of Helicobacter pylori on reactive oxygen-induced gastric epithelial cell injury. Carcinogenesis 2000, 21, 2091–2095. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.T.; Ramanujam, K.S.; Mobley, H.L.; Musselman, R.F.; James, S.P.; Meltzer, S.J. Helicobacter pylori stimulates inducible nitric oxide synthase expression and activity in a murine macrophage cell line. Gastroenterology 1996, 111, 1524–1533. [Google Scholar] [CrossRef]
- Katsurahara, M.; Kobayashi, Y.; Iwasa, M.; Ma, N.; Inoue, H.; Fujita, N.; Tanaka, K.; Horiki, N.; Gabazza, E.C.; Takei, Y. Reactive nitrogen species mediate DNA damage in Helicobacter pylori-infected gastric mucosa. Helicobacter 2009, 14, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kregel, K.C.; Zhang, H.J. An integrated view of oxidative stress in aging: Basic mechanisms, functional effects, and pathological considerations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R18–R36. [Google Scholar] [CrossRef] [PubMed]
- Drake, I.M.; Mapstone, N.P.; Schorah, C.J.; White, K.L.; Chalmers, D.M.; Dixon, M.F.; Axon, A.T. Reactive oxygen species activity and lipid peroxidation in Helicobacter pylori associated gastritis: Relation to gastric mucosal ascorbic acid concentrations and effect of H. pylori eradication. Gut 1998, 42, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Mori, M.; Seto, K.; Kai, A.; Kawaguchi, C.; Suzuki, M.; Suematsu, M.; Yoneta, T.; Miura, S.; Ishii, H. Helicobacter pylori-associated gastric pro- and antioxidant formation in Mongolian gerbils. Free Radic. Biol. Med. 1999, 26, 679–684. [Google Scholar] [CrossRef]
- Mai, U.E.H.; Perez-Perez, G.I.; Allen, J.B.; Wahl, S.M.; Blaser, M.J.; Smith, P.D. Surface proteins from Helicobacter pylori exhibit chemotactic activity for human leukocytes and are present in gastric mucosa. J. Exp. Med. 1992, 175, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Craig, P.M.; Territo, M.C.; Karnes, W.E.; Walsh, J.H. Helicobacter pylori secretes a chemotactic factor monocytes and neutrophils. Gut 1992, 33, 1020–1023. [Google Scholar] [CrossRef] [PubMed]
- Krawisz, J.E.; Sharon, P.; Stenson, W.F. Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Assessment of inflammation in rat and hamster models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [PubMed]
- Bishayee, A.; Dhir, N. Resveratrol-mediated chemoprevention of diethylnitrosamine-initiated hepatocarcinogenesis: Inhibition of cell proliferation and induction of apoptosis. Chem. Biol. Interact. 2009, 179, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Jiang, A.M.; Ma, Z.Y.; Li, X.B.; Xiong, Y.Y.; Dou, J.F.; Wang, J.F. The synthetic antimicrobial peptide pexiganan and its nanoparticles (PNPs) exhibit the anti-Helicobacter pylori activity in vitro and in vivo. Molecules 2015, 20, 3972–3985. [Google Scholar] [CrossRef] [PubMed]
- Guy, B.; Hessler, C.; Fourage, S.; Haensler, J.; Vialon-Lafay, E.; Rokbi, B. Systemic immuniza- tion with urease protects mice against Helicobacter pylori infection. Vaccine 1998, 16, 850–856. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohnishi, N.; Yagi, K. Assay for lipid peroxides for animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Wu, Y.H.; Zang, X.; Wang, D.H. Role of asymmetric dimethylarginine in acute lung injury induced by cerebral ischemia/reperfusion injury in rats. J. South. Med. Univ. 2011, 31, 1289–1293. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Jiang, A.; Qi, B.; Ma, Z.; Xiong, Y.; Dou, J.; Wang, J. Resveratrol Protects against Helicobacter pylori-Associated Gastritis by Combating Oxidative Stress. Int. J. Mol. Sci. 2015, 16, 27757-27769. https://doi.org/10.3390/ijms161126061
Zhang X, Jiang A, Qi B, Ma Z, Xiong Y, Dou J, Wang J. Resveratrol Protects against Helicobacter pylori-Associated Gastritis by Combating Oxidative Stress. International Journal of Molecular Sciences. 2015; 16(11):27757-27769. https://doi.org/10.3390/ijms161126061
Chicago/Turabian StyleZhang, Xiaolin, Anmin Jiang, Banghua Qi, Zhongyou Ma, Youyi Xiong, Jinfeng Dou, and Jianfei Wang. 2015. "Resveratrol Protects against Helicobacter pylori-Associated Gastritis by Combating Oxidative Stress" International Journal of Molecular Sciences 16, no. 11: 27757-27769. https://doi.org/10.3390/ijms161126061
APA StyleZhang, X., Jiang, A., Qi, B., Ma, Z., Xiong, Y., Dou, J., & Wang, J. (2015). Resveratrol Protects against Helicobacter pylori-Associated Gastritis by Combating Oxidative Stress. International Journal of Molecular Sciences, 16(11), 27757-27769. https://doi.org/10.3390/ijms161126061