
Antioxidant and ACE Inhibitory Bioactive Peptides Purified from Egg Yolk Proteins

Abstract
:1. Introduction
2. Results and Discussion
2.1. Protein Extraction and Quantification
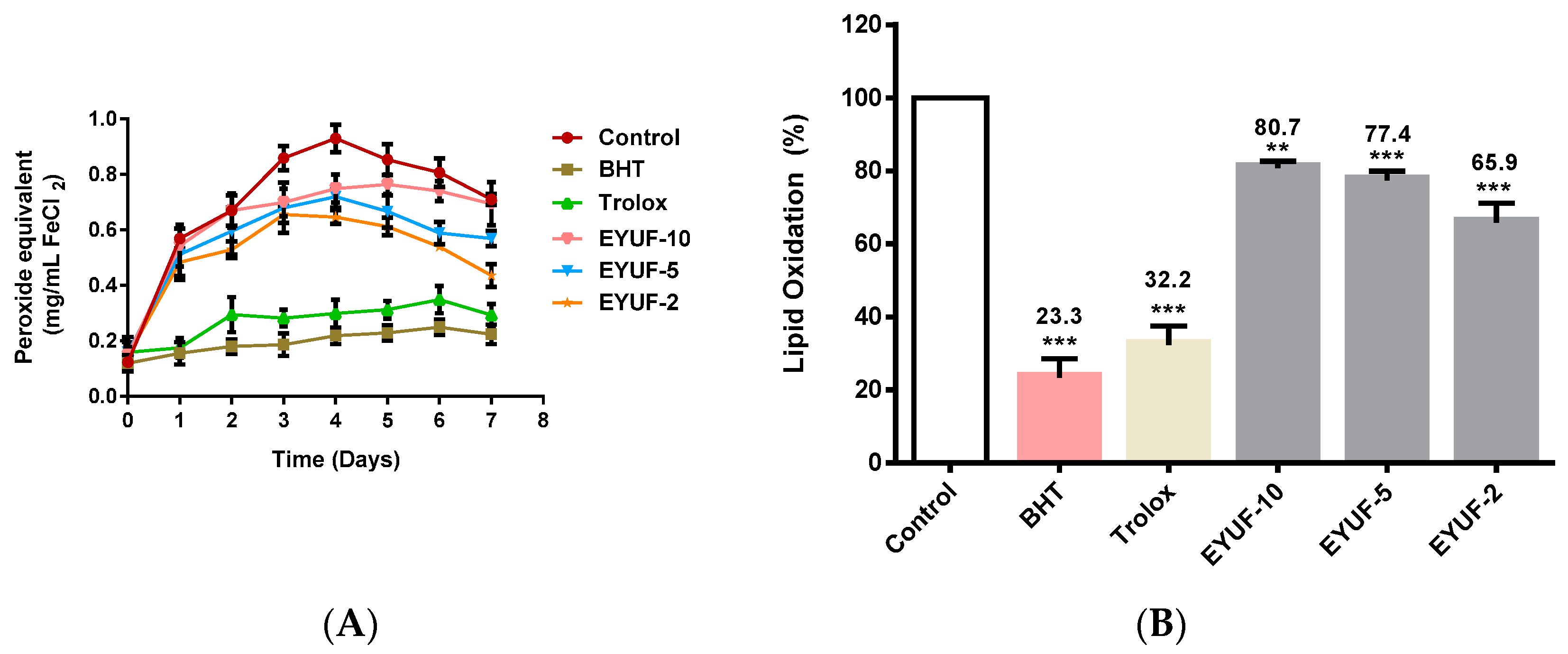
2.2. Egg Yolk Protein Hydrolysis and Ultrafiltration
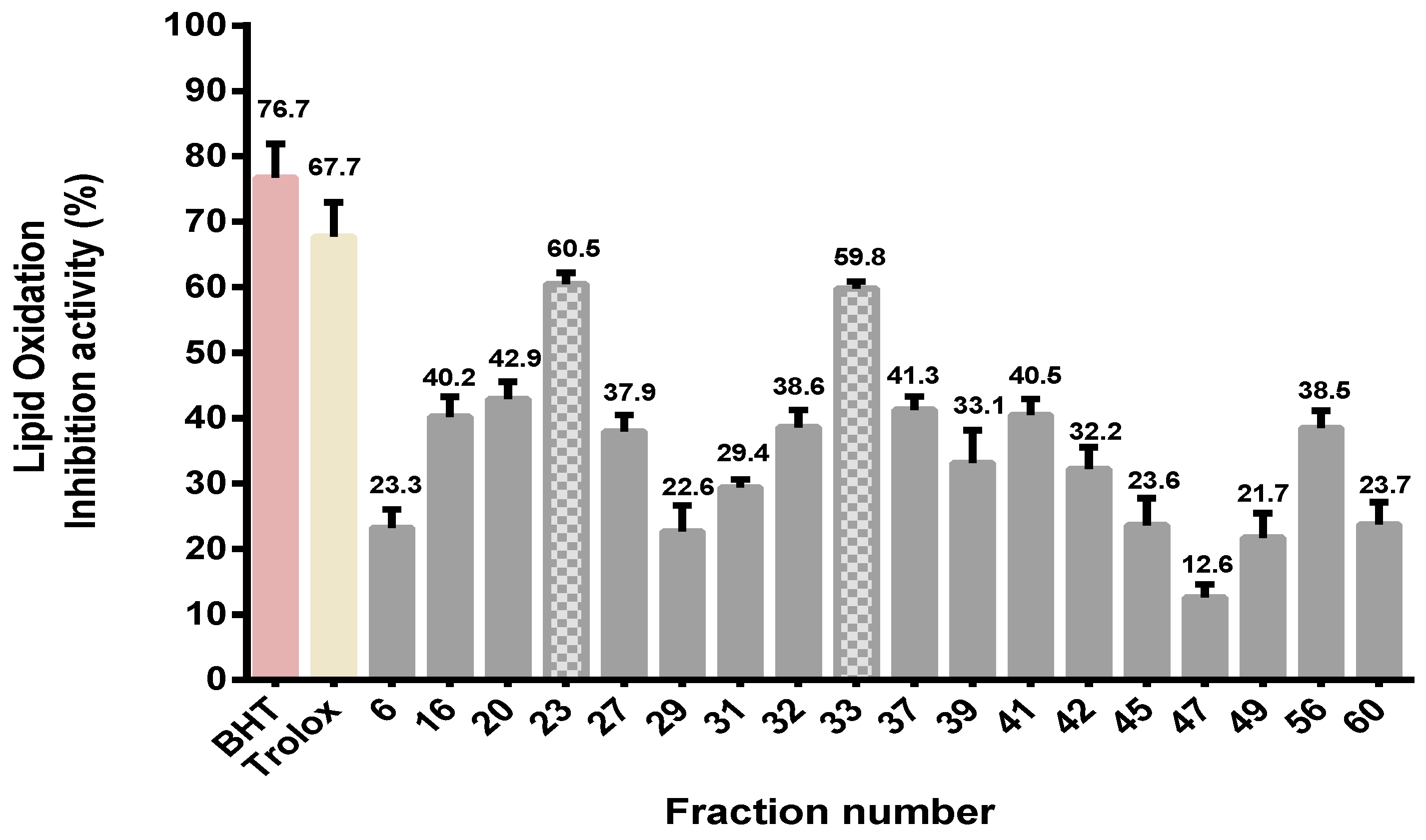
2.3. Purification of Antioxidative Peptides
2.3.1. Measurement of Lipid Oxidation Inhibition Activity of Ultrafiltration Fractions


2.3.2. Measurement of Lipid Oxidation Inhibition Activity of Purified Peptide after Gel Filtration


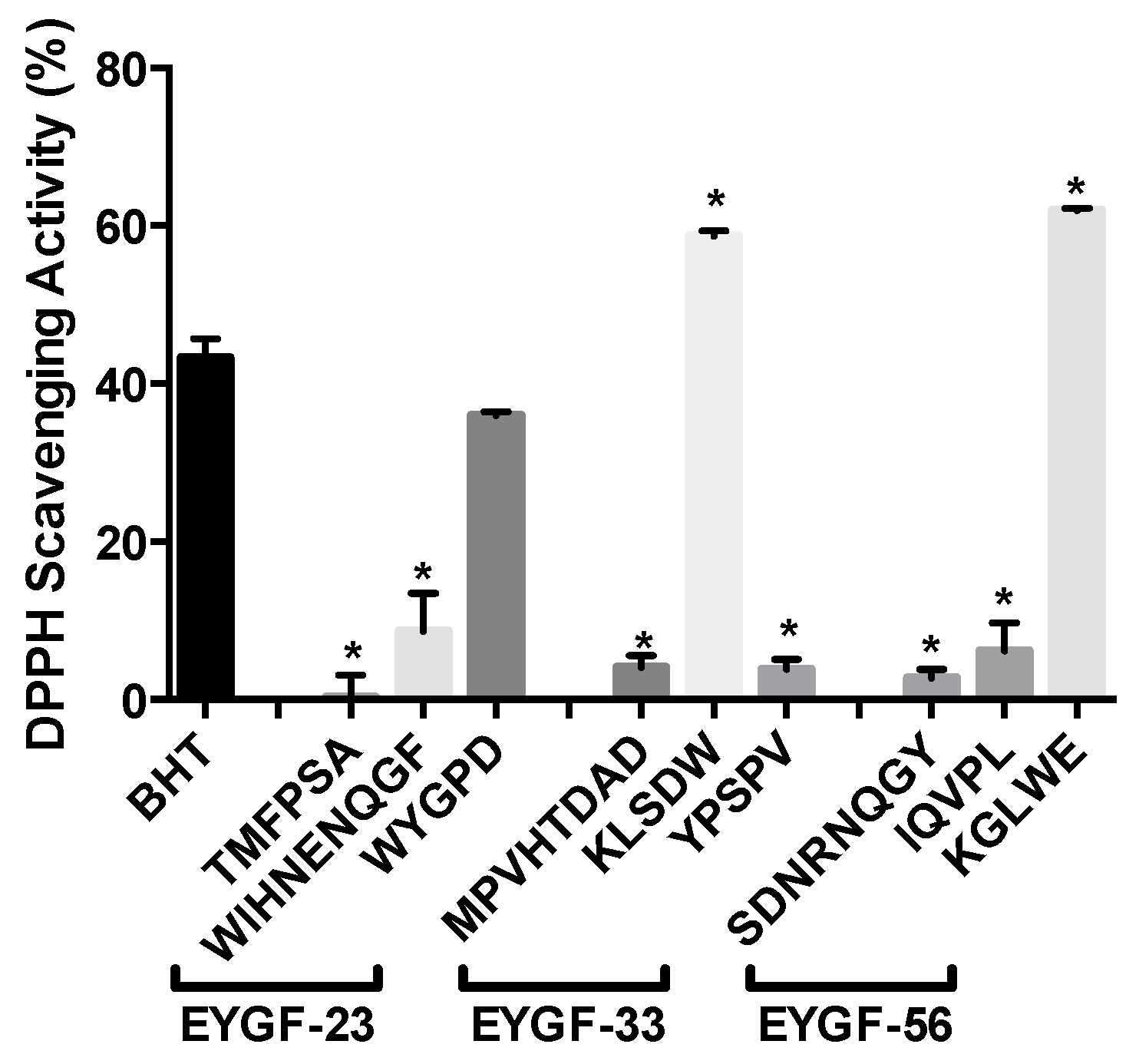
2.4. Antioxidant Activities of Egg Yolk Peptides
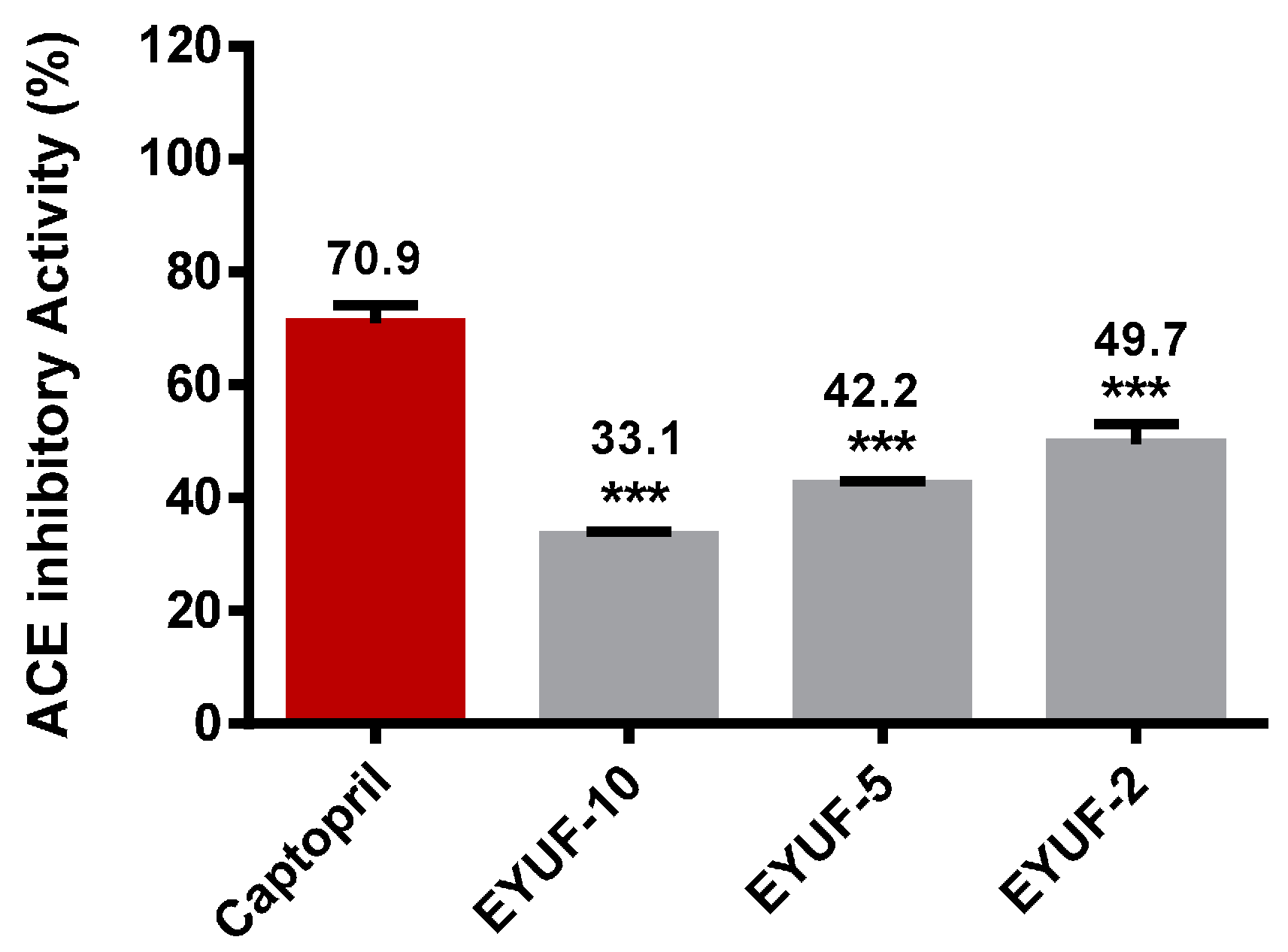
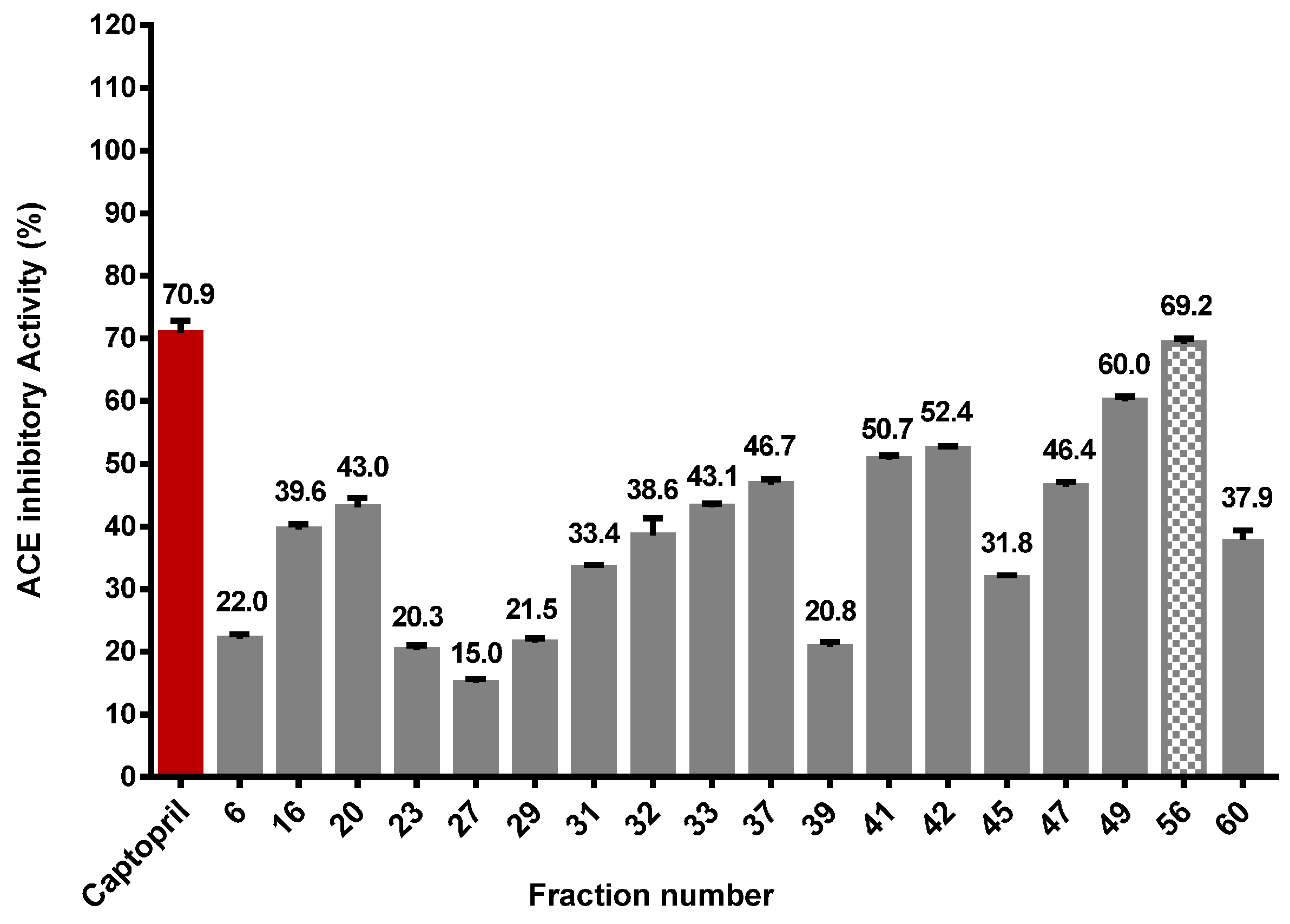
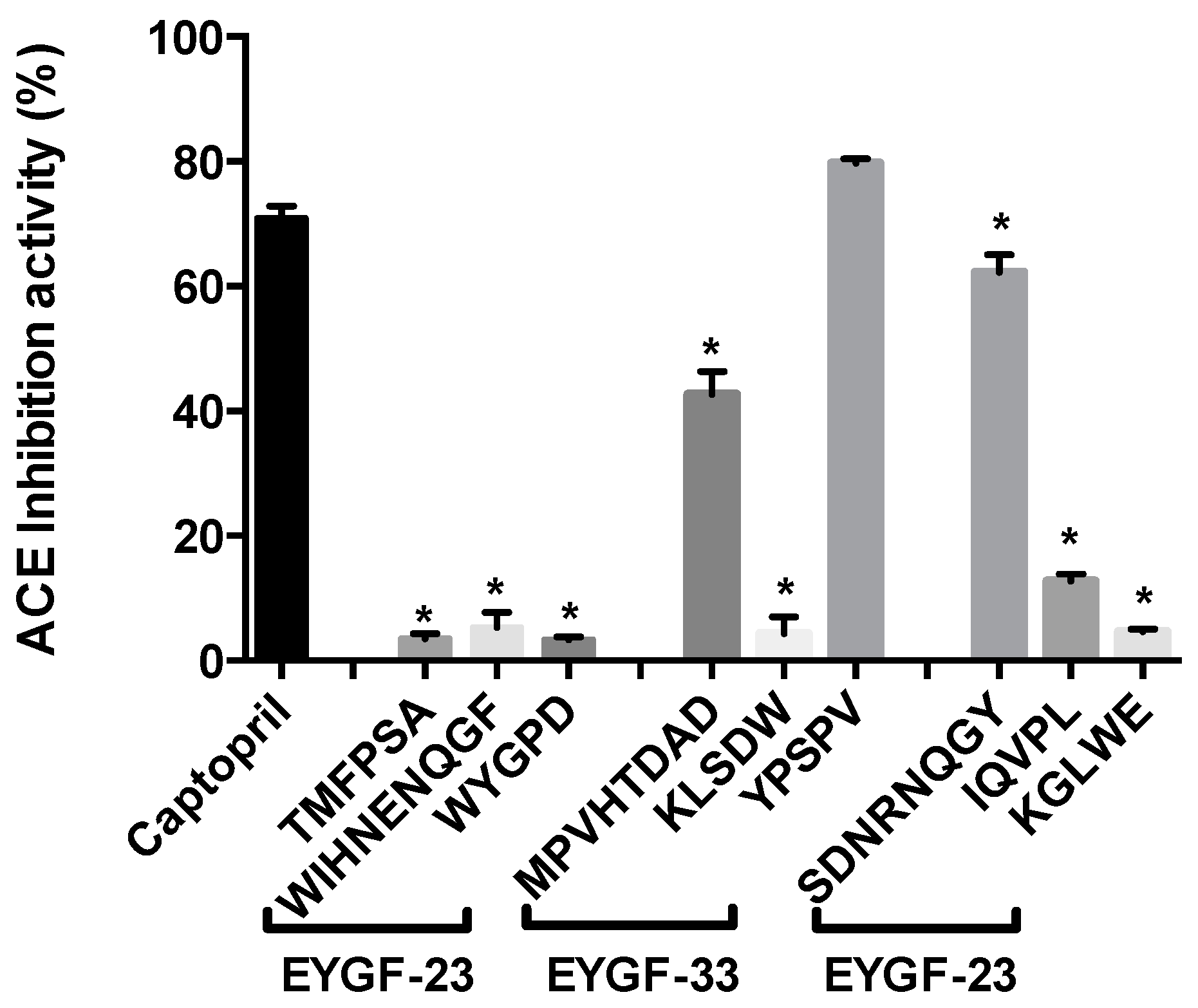
2.5. Purification of Angiotensin Converting Enzyme (ACE) Inhibitory Peptide

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | Sample Concentration (mg/mL) | Trolox a | BHT b | |||||
| 0.5 | 1 | 5 | 10 | 20 | 0.2 mg/mL | 0.2 mg/mL | ||
| DPPH Scavenging activity | EYGF-23 | 8.10% ± 1.43% a,b | 11.32% ± 1.05% a,b | 15.61% ± 2.30% a,b | 19.40% ± 1.10% a | 28.46% ± 2.13% a,b | 38.46% ± 1.43% | 23.60% ± 1.66% |
| EYGF-33 | 6.15% ± 1.15% a,b | 10.99% ± 1.90% a,b | 13.58% ± 1.16% a,b | 16.47% ± 1.49% a,b | 30.65% ± 2.86% a,b | |||
| Hydroxyl radical scavenging activity | EYGF-23 | 5.62% ± 2.33% a,b | 10.575% ± 1.36% a,b | 26.05% ± 2.86% a,b | 37.15% ± 2.15% a,b | 62.37% ± 2.21% a | 70.03% ± 3.48% | 58.97% ± 1.02% |
| EYGF-33 | 5.08% ± 1.44% a,b | 12.66% ± 1.74% a,b | 26.78% ± 1.57% a,b | 42.50% ± 2.45% a,b | 69.28% ± 3.62% b | |||
| Superoxide anion scavenging activity | EYGF-23 | 11.63% ± 2.32% a,b | 18.12% ± 0.51% a,b | 30.62% ± 1.86% a,b | 41.01% ± 1.70% a,b | 67.66% ± 2.14% | 63.48% ± 1.63% | 67.45% ± 1.67% |
| EYGF-33 | 11.59% ± 1.70% a,b | 28.13% ± 2.77% a,b | 35.43% ± 3.90% a,b | 48.85% ± 3.14% a,b | 82.88% ± 2.49% a,b | |||
| (B) | Sample Concentration (mg/mL) | EDTA a | ||||||
| 0.5 | 1 | 5 | 10 | 20 | (0.2 mg/mL) | |||
| EYGF-23 | 12.69% ± 1.15% a | 20.32% ± 1.79% a | 32.45% ± 1.84% a | 64.05% ± 1.49% a | 94.01% ± 1.40% a | 97.54% ± 1.79% | ||
| EYGF-33 | 10.24% ± 0.99% a | 24.39% ± 0.94% a | 41.37% ± 0.98% a | 61.88% ± 1.41% a | 92.22% ± 1.66% a | |||


2.6. Amino Acid Analysis and Sequencing
| Amino Acids | EYGF-23 (%) | EYGF-33 (%) | EYGF-56 (%) |
|---|---|---|---|
| Aspartic acid | 3.02 ± 0.27 | 6.22 ± 0.02 | 3.08 ± 0.33 |
| Glutamic acid | 3.41 ± 0.26 | 6.55 ± 0.014 | 9.30 ± 0.18 |
| Serine | 3.7 ± 0.13 | 5.77 ± 0.001 | 3.70 ± 0.22 |
| Glycine | 2.60 ± 0.11 | 5.70 ± 0.015 | 3.18 ± 0.45 |
| Histidine | 1.91 ± 0.26 | 1.21 ± 0.11 | 0.87 ± 0.57 |
| Arginine | 3.82 ±0.12 | 11.80 ± 0.001 | 16.25 ±0.53 |
| Threonine | 1.101 ± 0.04 | 0.01 ± 0.2 | 0.52 ± 0.02 |
| Alanine | 2.4 ± 0.14 | 7.80 ± 0.05 | 1.75 ± 0.32 |
| Proline | 8.01 ± 0.07 | 5.12 ± 0.006 | 1.68 ± 0.15 |
| Tyrosine | 12.40 ± 0.16 | 5.01 ± 0.011 | 1.51 ± 0.26 |
| Valine | 5.30 ± 0.23 | 6.71 ± 0.003 | 2.76 ± 0.52 |
| Methionine | 2.72 ± 0.17 | 0.03 ± 0.021 | 3.15± 0.22 |
| Cysteine | 0.90± 0.17 | 0.04 ± 0.13 | 0.99 ± 0.24 |
| Isoleucine | 3.12 ± 0.05 | 0.10 ± 0.09 | 1.28 ± 0.19 |
| Leucine | 5.7 ± 0.39 | 13.82 ± 0.013 | 21.64 ± 0.25 |
| Phenylalanine | 2.04 ± 0.10 | 1.05 ± 0.17 | 2.98 ± 0.24 |
| Tryptophan | 16.40 ± 0.07 | 9.44 ± 0.016 | 14.25 ± 0.29 |
| Lysine | 11.03 ± 0.02 | 8.27 ± 0.003 | 8.05 ± 0.91 |


3. Experimental Section
3.1. Protein Extraction and Quantification
3.2. Preparation of Protein Hydrolysates by Enzymatic Digestion
3.3. Fractionation of Egg Yolk Hydrolysate by Ultrafiltration
3.4. Purification Using Gel Filtration Chromatography
3.5. Evaluation of Lipid Oxidation in Linoleic Acid Model System
3.5.1. Ferric Thiocyanate (FTC) for Peroxide Formation
3.5.2. Determination of Thiobarbituric Acid Reactive Species (TBARS)
3.6. Measurement of Antioxidant Activity
3.6.1. Scavenging Activity on DPPH (1,1-Diphenyl-2-Picrylhydrazyl) Radical
3.6.2. Hydroxyl Radical Scavenging Activity (HRSA) Assay
3.6.3. Superoxide Anion Scavenging Activity (Pyrogallol)
3.6.4. Ferrous Chelating Activity Assay
3.7. Determination of Angiotensin Converting Enzyme Inhibitory Activity
3.8. Amino Acid Composition of Isolated Egg Yolk Peptide
3.9. Peptide Sequencing by LC/ESI MS/MS
3.10. Peptide synthesis
3.11. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van Nieuwenhuyzen, W. Lecithin production and properties. J. Am. Oil Chem. Soc. 1976, 53, 425–427. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Haque, E.; Chand, R. Antihypertensive and antimicrobial bioactive peptides from milk proteins. Eur. Food Res. Technol. 2008, 227, 7–15. [Google Scholar] [CrossRef]
- Alemán, A.; Pérez-Santín, E.; Bordenave-Juchereau, S.; Arnaudin, I.; Gómez-Guillén, M.C.; Montero, P. Squid gelatin hydrolysates with antihypertensive, anticancer and antioxidant activity. Food Res. Int. 2011, 44, 1044–1051. [Google Scholar] [CrossRef]
- Théolier, J.; Hammami, R.; Labelle, P.; Fliss, I. Isolation and identification of antimicrobial peptides derived by peptic cleavage of whey protein isolate. J. Funct. Foods 2013, 5, 706–714. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.; Li-Chan, E.C. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Food 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Alghazeer, R.; Saeed, S.; Howell, N.K. Aldehyde formation in frozen mackerel (Scomber scombrus) in the presence and absence of instant green tea. Food Chem. 2008, 108, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Sakagami, H.; Fugisawa, S. Cytotoxicity and apoptosis induction by butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT). Anticancer Res. 2003, 23, 4693–4701. [Google Scholar] [PubMed]
- Vandghanooni, S.; Forouharmehr, A.; Eskandani, M.; Barzegari, A.; Karil, V.; Kashanian, S.; Ezzati Nazhand Dolatabadi, J. Cytotoxicity and DNA fragmentation properties of butylated hydroxyanisole. DNA Cell Biol. 2013, 32, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F. Nutraceuticals and functional foods: Whole versus processed foods. Trends Food Sci. Technol. 2009, 20, 376–387. [Google Scholar] [CrossRef]
- Megías, C.; Yust, M.D.D.; Pedroche, J.; Lquari, H.; Girón-Calle, J.; Alaiz, M.; Millán, F.; Vioque, J. Purification of an ACE inhibitory peptide after hydrolysis of sunflower (Helianthus annuus L.) protein isolates. J. Agric. Food Chem. 2004, 52, 1928–1932. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Gibbs, B.F.; Zougman, A.; Masse, R.; Mulligan, C. Production and characterization of bioactive peptides from soy hydrolysate and soy-fermented food. Food Res. Int. 2004, 37, 123–131. [Google Scholar] [CrossRef]
- Miguel, M.; Lopez-Fandino, R.; Ramos, M.; Aleixandre, A. Short-term effect of egg-white hydrolysate products on the arterial blood pressure of hypertensive rats. J. Nutr. 2005, 94, 731–737. [Google Scholar] [CrossRef]
- Cinq-Mars, C.D.; Hu, C.; Kitts, D.D.; Li-Chan, E.C.Y. investigations into inhibitor type and mode, simulated gastrointestinal digestion, and cell transport of the angiotensin I-converting enzyme–inhibitory peptides in Pacific hake (Merluccius productus) fillet hydrolysate. J. Agric. Food Chem. 2007, 56, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Samaraweera, H.; Zhang, W.; Lee, E.J.; Ahn, D.U. Egg yolk phosvitin and functional phosphopeptides—Review. J. Food Sci. 2011, 76, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wu, H.; Li, X.; Lai, F.; Xiao, X. Purification and characterization of high antioxidant peptides from duck egg white protein hydrolysates. Biochem. Biophys. Res. Commun. 2014, 452, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Zambrowicz, A.; Pokora, M.; Eckert, E.; Szoltysik, M.; Dabrowska, A.; Chrzanowska, J.; Trziszka, T. Angiotensin-I converting enzyme inhibitory and antioxidant activities of egg protein hydrolysates produced with gastrointestinal and nongastrointestinal enzymes. J. Funct. Food Health Dis. 2012, 2, 487–500. [Google Scholar]
- Park, P.J.; Jung, W.K.; Nam, K.S.; Shahidi, F.; Kim, S.K. Purification and characterization of antioxidative peptides from protein hydrolysate of lecithin-free egg yolk. J. Am. Oil Chem. Soc. 2001, 78, 651–656. [Google Scholar] [CrossRef]
- Sakanaka, S.; Tachibana, Y.; Ishihara, N.; Raj Juneja, L. Antioxidant activity of egg-yolk protein hydrolysates in a linoleic acid oxidation system. Food Chem. 2004, 86, 99–103. [Google Scholar] [CrossRef]
- Sakanaka, S.; Tachibana, Y. Active oxygen scavenging activity of egg-yolk protein hydrolysates and their effects on lipid oxidation in beef and tuna homogenates. Food Chem. 2006, 95, 243–249. [Google Scholar] [CrossRef]
- Young, D.; Nau, F.; Pasco, M.; Mine, Y. Identification of hen egg yolk-derived phosvitin phosphopeptides and their effects on gene expression profiling against oxidative stress-induced Caco-2 cells. J. Agric. Food Chem. 2011, 95, 9207–9218. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. The World Health Statistics. 2012. Available online: http://www.who.int/entity/mediacentre/news/releases/2012/world_health_statistics (accessed on 11 November 2013).
- Crowley, S.D.; Gurley, S.B.; Herrera, M.J.; Ruiz, P.; Griffiths, R.; Kumar, A.P.; Kim, H.S.; Smithies, O.; Le, T.H.; Coffman, T.M. Angiotensin II causes hypertension and cardiac hypertrophy through its receptors in the kidney. Proc. Natl. Acad. Sci. USA 2006, 103, 17985–17990. [Google Scholar] [CrossRef] [PubMed]
- Tschope, C.; Schultheiss, H.P.; Walther, T. Multiple interactions between the renin-angiotensin and the kallikrein-kinin systems: Role of ACE inhibition and AT1 receptor blockade. J. Cardiovasc. Pharmacol. 2002, 39, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, T.; Gandhi, T.K.; Fiskio, J.M.; Seger, A.C.; So, J.W.; Cook, E.F.; Fukui, T.; Bates, D.W. An evaluation of risk factors for adverse drug events associated with angiotensin-converting enzyme inhibitors. J. Eval. Clin. Pract. 2004, 10, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Cao, J.; Guo, H.; Zhao, B. Studies on purification and the molecular mechanism of a novel ACE inhibitory peptide from whey protein hydrolysate. Food Chem. 2012, 130, 121–126. [Google Scholar] [CrossRef]
- Zhipeng, Y.; Wenzhu, Z.; Jingbo, L.; Jing, L.; Feng, C. A novel angiotensin I-converting enzyme-inhibitory peptide from egg white protein QIGLF. J. Sci. Food Agric. 2011, 91, 921–926. [Google Scholar]
- Eckert, E.; Zambrowicz, A.; Pokora, M.; Setner, B.; Dąbrowska, A.; Szołtysik, M.; Szewczuk, Z.; Polanowski, A.; Trziszka, T.; Chrzanowska, J. Egg-yolk protein by-product as a source of ACE-inhibitory peptides obtained with using unconventional proteinase from Asian pumpkin (Cucurbita ficifolia). J. Proteom. 2014, 110, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.M.; Li-Chan, E.C. Angiotensin I converting enzyme inhibitory peptides from in vitro pepsin-pancreatin digestion of soy protein. J. Agric. Food Chem. 2005, 53, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.; Fitzgerald, R.J. Antihypertensive Peptides from Food Proteins. Bioactive Food Peptides in Health and Disease, 2nd ed.; Intech Publishers Inc.: New York, NY, USA, 2013; pp. 45–72. [Google Scholar]
- Chen, H.M.; Muramoto, K.; Yamauchi, F.; Nokihara, K. Antioxidant activity of designed peptides based on the antioxidative peptide isolated from digests of a soybean protein. J. Agric. Food Chem. 1996, 44, 2619–2623. [Google Scholar] [CrossRef]
- Vioque, J.; Sáchez-vioque, R.; Clemente, A.; Pedroche, J.; Millán, F. Partially hydrolyzed rapeseed protein isolates with improved functional properties. J. Am. Oil Chem. Soc. 2000, 77, 447–450. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Autolysis-assisted production of fish protein hydrolysates with antioxidant properties from Pacific hake (Merluccius productus). Food Chem. 2008, 107, 768–776. [Google Scholar] [CrossRef]
- Jamdar, S.N.; Rajalakshmi, V.; Pednekar, M.D.; Juan, F.; Yardi, V.; Sharma, A. Influence of degree of hydrolysis on functional properties, antioxidant activity and ACE inhibitory activity of peanut protein hydrolysate. Food Chem. 2010, 121, 178–184. [Google Scholar] [CrossRef]
- Chen, H.M.; Muramoto, K.; Yamauchi, F. Structural analysis of antioxidative peptides from soybean β-conglycinin. J. Agric. Food Chem. 1995, 43, 574–578. [Google Scholar]
- Howell, N.K.; Kasase, C. Bioactive peptides and proteins from fish muscle and collagen. In Bioactive Proteins and Peptides as Functional Foods and Nutraceuticals; Blackwell Publishing Ltd. and Institute of Food Technologists Press: Ames, Iowa, IA, USA, 2010; pp. 203–224. [Google Scholar]
- Pihlanto, A.; Mäkinen, S. Antihypertensive Properties of Plant Protein Derived Peptides. J. Agric. Food Chem. 2013, 57, 5113–5120. [Google Scholar]
- Dai, C.; Ma, H.; Luo, L.; Yin, X. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. Eur. Food Res. Technol. 2013, 236, 681–689. [Google Scholar] [CrossRef]
- Wang, J.S.; Zhao, M.M.; Zhao, Q.Z.; Jiang, Y.M. Antioxidant properties of papain hydrolysates of wheat gluten in different oxidation systems. Food Chem. 2007, 101, 1658–1663. [Google Scholar] [CrossRef]
- Luo, H.Y.; Wang, B.; Li, Z.; Chi, C.F.; Zhang, Q.H.; He, G.Y. Preparation and evaluation of antioxidant peptide from papain hydrolysate of Sphyrna lewini muscle protein. LWT Food Sci. Technol. 2013, 51, 281–288. [Google Scholar] [CrossRef]
- Saeed, S.; Gillies, D.; Wagner, G.; Howell, N.K. ESR and NMR spectroscopy studies on protein oxidation and formation of dityrosine in emulsions containing oxidised methyl linoleate. Food Chem. Toxicol. 2006, 44, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Xiong, Y.L.; Zhai, J.; Zhu, H.; Dziubla, T. Fractionation and evaluation of radical-scavenging peptides from in vitro digests of buckwheat protein. Food Chem. 2010, 118, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Le, G.W.; Shi, Y.H.; Shrestha, S. Angiotensin I–converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Murray, B.A.; Fitzgerald, R.J. Angiotensin converting enzyme inhibitory peptides derived from food proteins: Biochemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 773–791. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists International. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists International: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Osawa, T.; Namiki, M. Natural antioxidants isolated from Eucalyptus leaf waxes. J. Agric. Food Chem. 1985, 33, 777–780. [Google Scholar] [CrossRef]
- Bersuder, P.; Hole, M.; Smith, G. Antioxidants from a heated histidine-glucose model system. I: Investigation of the antioxidant role of histidine and isolation of antioxidants by high-performance liquid chromatography. J. Am. Oil Chem. Soc. 1998, 75, 181–187. [Google Scholar]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimzadeh, M.; Nabavi, S.; Nabavi, S. Antioxidant activities of methanol extract of Sambucus ebulus L. flower. Pak. J. Biol. Sci. 2009, 12, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Aluko, R.E.; Muir, A.D. Improved method for direct high-performance liquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions. J. Chromatogr. A 2002, 950, 125–130. [Google Scholar] [PubMed]
- Badii, F.; Howell, N. Fish gelatin: Structure, gelling properties and interaction with egg albumen proteins. Food Hydrocoll. 2004, 20, 630–640. [Google Scholar] [CrossRef]
- Yi, E.C.; Lee, H.; Aebersold, R.; Goodlett, D.R. A microcapillary trap cartridge-microcapillary high-performance liquid chromatography electrospray ionization emitter device capable of peptide tandem mass spectrometry at the attomole level on an ion trap mass spectrometer with automated routine operation. Rapid Commun. Mass Spectrom. 2003, 17, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousr, M.; Howell, N. Antioxidant and ACE Inhibitory Bioactive Peptides Purified from Egg Yolk Proteins. Int. J. Mol. Sci. 2015, 16, 29161-29178. https://doi.org/10.3390/ijms161226155
Yousr M, Howell N. Antioxidant and ACE Inhibitory Bioactive Peptides Purified from Egg Yolk Proteins. International Journal of Molecular Sciences. 2015; 16(12):29161-29178. https://doi.org/10.3390/ijms161226155
Chicago/Turabian StyleYousr, Marwa, and Nazlin Howell. 2015. "Antioxidant and ACE Inhibitory Bioactive Peptides Purified from Egg Yolk Proteins" International Journal of Molecular Sciences 16, no. 12: 29161-29178. https://doi.org/10.3390/ijms161226155
APA StyleYousr, M., & Howell, N. (2015). Antioxidant and ACE Inhibitory Bioactive Peptides Purified from Egg Yolk Proteins. International Journal of Molecular Sciences, 16(12), 29161-29178. https://doi.org/10.3390/ijms161226155