
Molecular Characterization of Vitellogenin and Its Receptor Genes from Citrus Red Mite, Panonychus citri (McGregor)


Abstract
:1. Introduction
2. Results
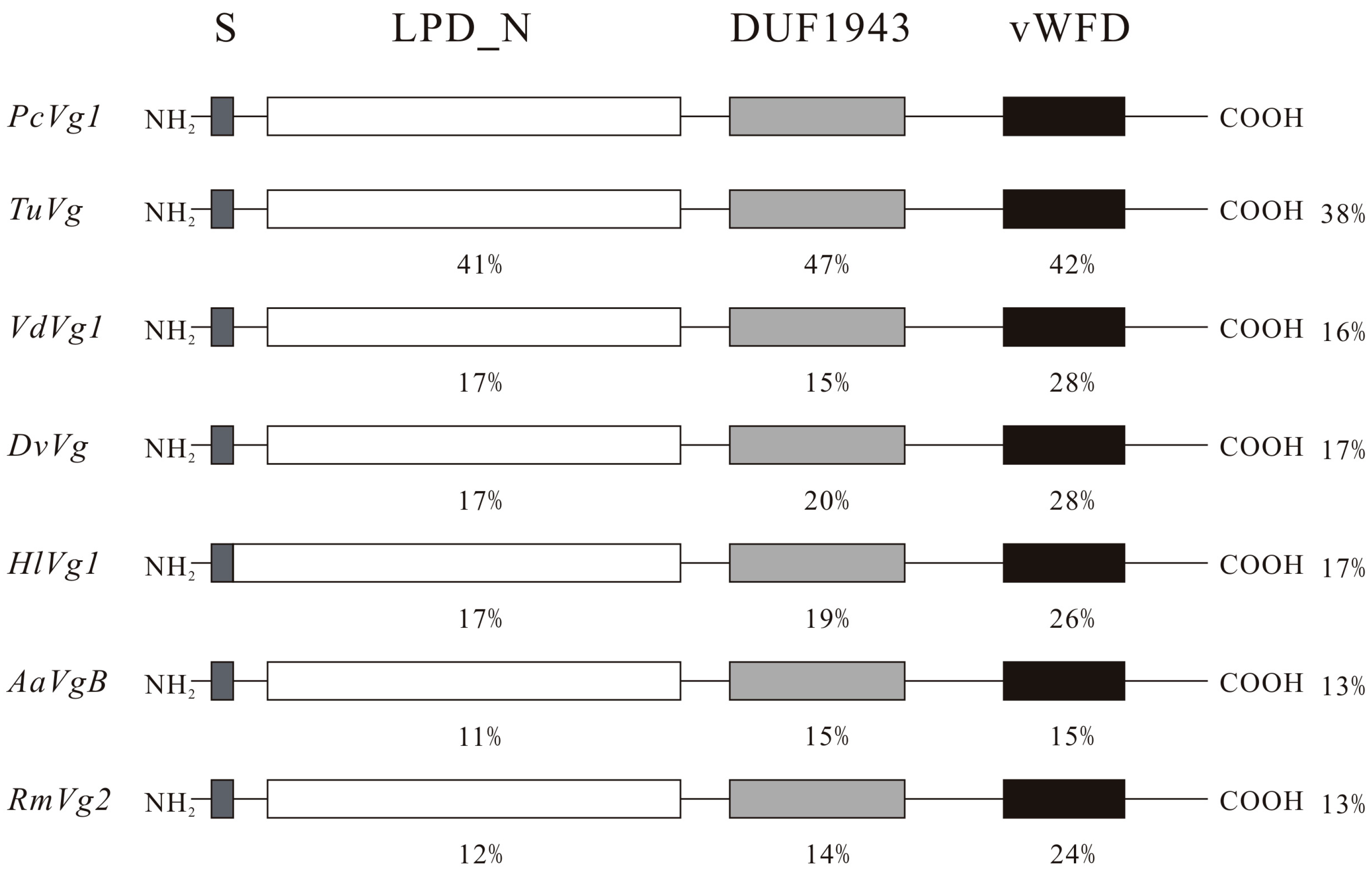
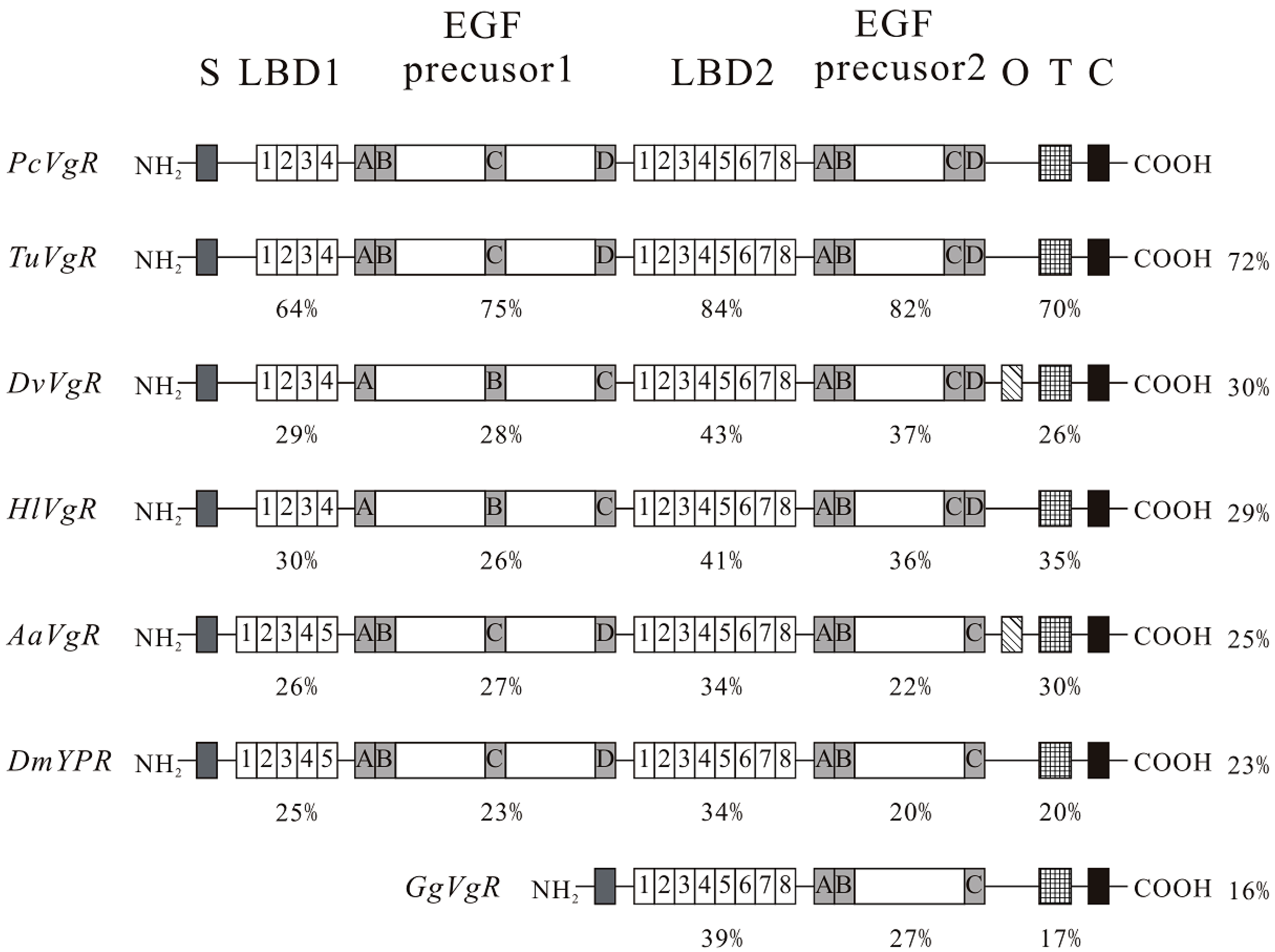
2.1. Nucleotide (nt) and Deduced Amino Acids (aa) Sequences
2.2. Phylogenetic Analyses of PcVg1 and PcVgR



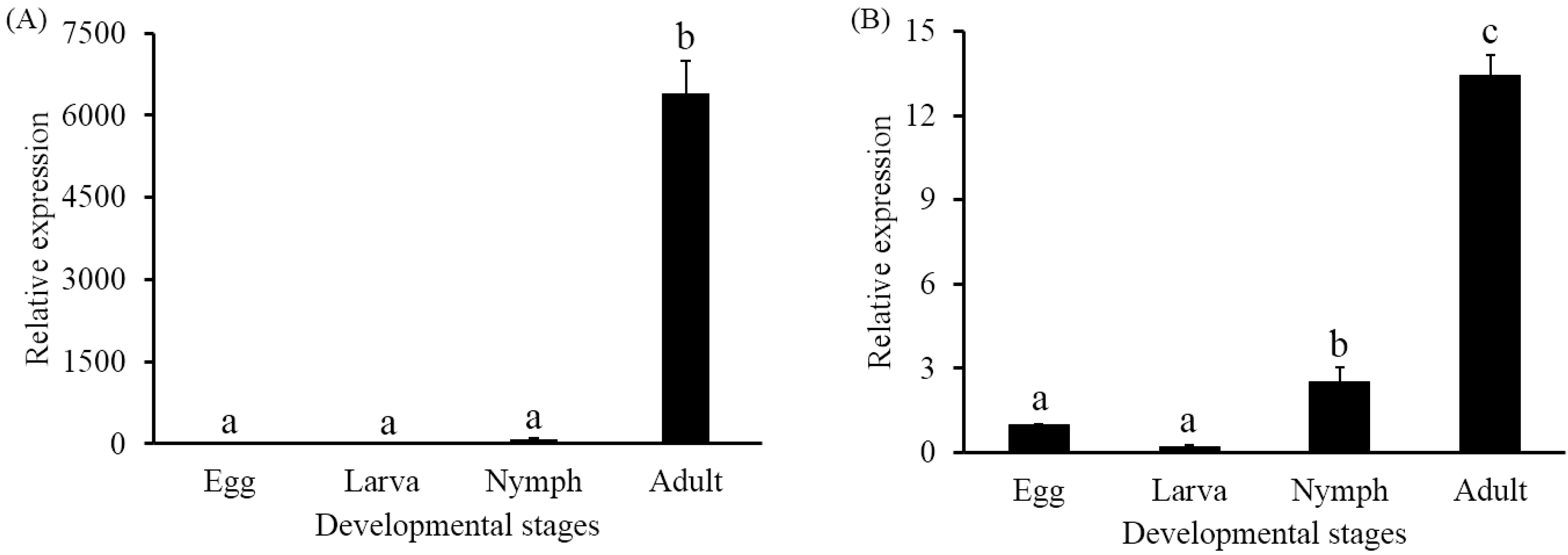
2.3. Developmental Expression Profiles

3. Discussion
4. Experimental Section
4.1. Mites
4.2. RNA Extraction and Transcription
4.3. Cloning Full-Length PcVg1 and PcVgR cDNAs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiments | Primer Names and Sequences (5' to 3') | Product Length (bp) |
|---|---|---|
| 3'-RACE | Vg1-S1: GGTCATTCCGAACTCTTTCC | 1939 |
| Vg1-S2: AGACGGTACCCACTACAACG | 1941 | |
| VgR-S1: ACATTTAGAGCCATTCGG | 454 | |
| VgR-S2: CCAGTGAGTTTGGACGAT | 303 | |
| 5'-RACE | Vg1-A1: TTCAGCGAGAACCATTTGGA | 1552 |
| Vg1-A2: TGAGTGTTCGGTGTTGGTGA | 1468 | |
| VgR-A1: ATCGAGGTGATTCATCGTCA | 1089 | |
| VgR-A2: TGAGCCATCGAAACAATCCT | 938 | |
| Splicing primer | VgR-S-S1: ACCCTGAGAAAGGTCTTATG | |
| VgR-S-A1: CAATGAAGGAGCACAATGT | 1481 | |
| VgR-S-S2: GTGCAATTACCTGTCCAC | ||
| VgR-S-A2: CAACCCAATAAATCATTTTC | 1197 | |
| Oligo (dT) primer | UPM:CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | |
| NUP: AAGCAGTGGTATCAACGCAGAGT | ||
| Full-length confirmation | Vg1-F-S1: TCGAACATGAAGATCGCTC | |
| Vg1-F-A1: GTGAGTTCCTTAAGAGCCAAGT | 1952 | |
| Vg1-F-S2: TCAGACTCACAAATCGATTACC | ||
| Vg1-F-A2: TCTTCGTGGGCAAGAGTT | 2267 | |
| Vg1-F-S3: AATACATCGCTAACCTTACCTG | ||
| Vg1-F-A3: GAGATGATTTAAATGCCTCG | 1682 | |
| VgR-F-S1: CTCAAAATGTGGCCTAAACTAGTC | ||
| VgR-F-A1: TCGGGGATAAAACTGGATG | 1909 | |
| VgR-F-S2: TCTCGTCGTGGTCGTTCA | ||
| VgR-F-A2: CCTCGTCCTCCTGGTTAACAC | 1873 | |
| VgR-F-S3: GTTGCTCTGATGGTCATTGT | ||
| VgR-F-A3: TTAAACTTTTATAAAAACACGTTGG | 2096 |
4.4. Sequence and Phylogenetic Analyses
4.5. qPCR
| Genes | GenBank No. | Primer Names and Sequences (5' to 3') | Amplicon Sizes (bp) | Amplification Efficiency (%) |
|---|---|---|---|---|
| PcVg1 | KC978893 | Vg1-RTS: GCCTCAAACGAAGCTCAATC | 183 | 97.7 |
| Vg1-RTA: AGCCAAAGCGTCGAGTAAAA | ||||
| PcVgR | KC978894 | VgR-RTS: TTGTTTCGATGGCTCAGATG | 150 | 108.9 |
| VgR-RTA: TCACCGTGTGGACAATCAGT | ||||
| GAPDH | HM582445 | GAP-RTS: CTTTGGCCAAGGTCATCAAT | 159 | 108.1 |
| GAP-RTA: CGGTAGCGGCAGGTATAATG |
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Takafuji, A.; Fujimoto, H. Winter survival of the non-diapausing population of the citrus red mite, Panonychus citri (McGregor) (Acarina: Tetranychidae) on pear and citrus. Appl. Entomol. Zool. 1986, 21, 467–473. [Google Scholar]
- Vassiliou, V.A.; Papadoulis, G. First record of the citrus red mite Panonychus citri in Cyprus. Phytoparasitica 2009, 37, 99–100. [Google Scholar] [CrossRef]
- Ding, T.B.; Niu, J.Z.; Yang, L.H.; Zhang, K.; Dou, W.; Wang, J.J. Transcription profiling of two cytochrome P450 genes potentially involved in acaricide metabolism in citrus red mite Panonychus citri. Pestic. Biochem. Physiol. 2013, 106, 28–37. [Google Scholar] [CrossRef]
- Li, Z.J.; Zhang, S.C.; Liu, Q.H. Vitellogenin functions as a multivalent pattern recognition receptor with an opsonic activity. PLoS One 2008, 3, e1940. [Google Scholar] [CrossRef] [PubMed]
- Reading, B.J.; Hiramatsu, N.; Sawaguchi, S.; Matsubara, T.; Hara, A. Conserved and variant molecular and functional features of multiple egg yolk precursor proteins (vitellogenins) in white perch (Morone americana) and other teleosts. Mar. Biotechnol. 2009, 11, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Ogihara, K.; Horigane, M.; Nakajima, Y.; Moribayashi, A.; Taylor, D. Ecdysteroid hormone titer and its relationship to vitellogenesis in the soft tick, Ornithodoros moubata (Acari: Argasidae). Gen. Comp. Endocriol. 2007, 150, 371–380. [Google Scholar] [CrossRef]
- Sappington, T.W.; Raikhel, A.S. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect Biochem. Mol. Biol. 1998, 28, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Raikhel, A.S.; Takeda, M. Biosynthesis and processing of insect vitellogenins. In Progress in Vitellogenesis: Reproductive Biology of Invertebrates; Raikhel, A.S., Sappington, T.W., Eds.; Science Publishers, Inc.: Plymouth, UK, 2005; Volume XII, pp. 1–32. [Google Scholar]
- Khalil, S.M.S.; Donohue, K.V.; Thompson, D.M.; Jeffers, L.A.; Ananthapadmanaban, U.; Sonenshine, D.E.; Mitchell, R.D.; Roe, R.M. Full-length sequence, regulation and developmental studies of a second vitellogenin gene from the American dog tick, Dermacentor variabilis. J. Insect Physiol. 2011, 57, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.; Snigirevskaya, E.S.; Raikhel, A.S. The cell biology of yolk protein precursor synthesis and secretion. In Progress in Vitellogenesis: Reproductive Biology of Invertebrates; Raikhel, A.S., Sappington, T.W., Eds.; Science Publishers, Inc.: Plymouth, UK, 2005; Volume XII, pp. 33–68. [Google Scholar]
- Piulachs, M.D.; Guidugli, K.R.; Barchuk, A.R.; Cruz, J.; Simoes, Z.L.P.; Belles, X. The vitellogenin of the honey bee, Apis mellifera: structural analysis of the cDNA and expression studies. Insect Biochem. Mol. Biol. 2003, 33, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Boldbaatar, D.; Battsetseg, B.; Matsuo, T.; Hatta, T.; Umemiya-Shirafuji, R.; Xuan, X.N.; Fujisaki, K. Tick vitellogenin receptor reveals critical role in oocyte development and transovarial transmission of Babesia parasite. Biochem. Cell Biol. 2008, 86, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, K.W.; Smolenaars, M.M.W.; van Hoof, D.; van der Horst, D.J. Sequence analysis of the non-recurring C-terminal domains shows that insect lipoprotein receptors constitute a distinct group of LDL receptor family members. Insect Biochem. Mol. Biol. 2006, 36, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.J. Vitellogenin receptors: Oocyte-specific members of the low-density lipoprotein receptor supergene family. Int. Rev. Cytol. 1996, 166, 103–137. [Google Scholar] [PubMed]
- Tufail, M.; Takeda, M. Molecular cloning, characterization and regulation of the cockroach vitellogenin receptor during oogenesis. Insect Mol. Biol. 2005, 14, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Reading, B.J.; Hiramatsu, N.; Schilling, J.; Molloy, K.T.; Glassbrook, N. Lrp13 is a novel vertebrate lipoprotein receptor that binds vitellogenins in teleost fishes. J. Lipid Res. 2014, 55, 2287–2295. [Google Scholar] [CrossRef] [PubMed]
- Davail, B.; Pakdel, F.; Bujo, H.; Perazzolo, L.M.; Waclawek, M.; Schneider, W.J.; Le Menn, F. Evolution of oogenesis: The receptor for vitellogenin from the rainbow trout. J. Lipid Res. 1998, 39, 1929–1937. [Google Scholar] [PubMed]
- Mouchel, N.; Trichet, V.; Betz, A.; LePennec, J.P.; Wolff, J. Characterization of vitellogenin from rainbow trout (Oncorhynchus mykiss). Gene 1996, 174, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Sappington, T.W.; Kokoza, V.A.; Cho, W.L.; Raikhel, A.S. Molecular characterization of the mosquito vitellogenin receptor reveals unexpected high homology to the Drosophila yolk protein receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 8934–8939. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.E.; Lewis, D.K.; Keeley, L.L.; Pietrantonio, P.V. cDNA cloning and transcriptional regulation of the vitellogenin receptor from the imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae). Insect Mol. Biol. 2004, 13, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Ciudad, L.; Piulachs, M.D.; Belles, X. Systemic RNAi of the cockroach vitellogenin receptor results in a phenotype similar to that of the Drosophila yolkless mutant. FEBS J. 2006, 273, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Takeda, M. Molecular cloning and developmental expression pattern of the vitellogenin receptor from the cockroach, Leucophaea maderae. Insect Biochem. Mol. Biol. 2007, 37, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Warrier, S.; Subramoniam, T. Receptor mediated yolk protein uptake in the crab Scylla serrata: Crustacean vitellogenin receptor recognizes related mammalian serum lipoproteins. Mol. Reprod. Dev. 2002, 61, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Grant, B.; Hirsh, D. Receptor-mediated endocytosis in the Caenorhabditis elegans oocyte. Mol. Biol. Cell 1999, 10, 4311–4326. [Google Scholar] [CrossRef] [PubMed]
- Deborah, M.T.; Khalil, S.M.S.; Jeffers, L.A.; Sonenshine, D.E.; Mitchell, R.D.; Osgood, C.J.; Roe, R.M. Sequence and the developmental and tissue-specific regulation of the first complete vitellogenin messenger RNA from ticks responsible for heme sequestration. Insect. Biochem. Mol. Biol. 2007, 37, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.D.; Ross, E.; Osgood, C.; Sonenshine, D.E.; Donohue, K.V.; Khalil, S.M.; Thompson, D.M.; Roe, R.M. Molecular characterization, tissue-specific expression and RNAi knockdown of the first vitellogenin receptor from a tick. Insect Biochem. Mol. Biol. 2007, 37, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Boldbaatar, D.; Umemiya-Shirafuji, R.; Liao, M.; Tanaka, T.; Xuan, X.N.; Fujisaki, K. Multiple vitellogenins from the Haemaphysalis longicornis tick are crucial for ovarian development. J. Insect Physiol. 2010, 56, 1587–1598. [Google Scholar] [CrossRef] [PubMed]
- Cabrera Cordon, A.R.; Shirk, P.D.; Duehl, A.J.; Evans, J.D.; Teal, P.E.A. Variable induction of vitellogenin genes in the varroa mite, Varroa destructor (Anderson & Trueman), by the honeybee, Apis mellifera L, host and its environment. Insect Mol. Biol. 2013, 22, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Online Resource for Community Annotation of Eukaryotes. Available online: http://bioinformatics.psb.ugent.be/orcae/overview/Tetur (accessed on 10 April 2014).
- Springer, T.A. An extracellular beta-propeller module predicted in lipoprotein and scavenger receptors, tyrosine kinases, epidermal growth factor precursor, and extracellular matrix components. J. Mol. Biol. 1998, 283, 837–862. [Google Scholar] [CrossRef] [PubMed]
- Horigane, M.; Shinoda, T.; Honda, H.; Taylor, D. Characterization of a vitellogenin gene reveals two phase regulation of vitellogenesis by engorgement and mating in the soft tick Ornithodoros moubata (Acari: Argasidae). Insect Mol. Biol. 2010, 19, 501–515. [Google Scholar] [PubMed]
- Kawakami, Y.; Goto, S.G.; Ito, K.; Numata, H. Suppression of ovarian development and vitellogenin gene expression in the adult diapause of the two-spotted spider mite Tetranychus urticae. J. Insect Physiol. 2009, 55, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Takeda, M. Molecular characteristics of insect vitellogenins. J. Insect Physiol. 2008, 54, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Prat, F.; Coward, K.; Sumpter, J.P.; Tyler, C.R. Molecular characterization and expression of two ovarian lipoprotein receptors in the rainbow trout, Oncorhynchus mykiss. Biol. Reprod. 1998, 58, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, N.; Chapman, R.W.; Lindzey, J.K.; Haynes, M.R.; Sullivan, C.V. Molecular characterization and expression of vitellogenin receptor from white perch (Morone americana). Biol. Reprod. 2004, 70, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Takeda, M. Insect vitellogenin/lipophorin receptors: Molecular structures, role in oogenesis, and regulatory mechanisms. J. Insect Physiol. 2009, 55, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.H.; Wang, J.W.; Lu, K.; Zhou, J.L.; Zhou, Q. The first vitellogenin receptor from a Lepidopteran insect: Molecular characterization, expression patterns and RNA interference analysis. Insect Mol. Biol. 2011, 20, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.C.; Raikhel, A.S. Organization and developmental expression of the mosquito vitellogenin receptor gene. Insect Mol. Biol. 2001, 10, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Niu, J.Z.; Ding, T.B.; Dou, W.; Wang, J.J. Molecular characterization of two carboxylesterase genes of the citrus red mite, Panonychus citri (Acari: Tetranychidae). Arch. Insect Biochem. Physiol. 2013, 82, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.Z.; Dou, W.; Ding, T.B.; Shen, G.M.; Zhang, K.; Smagghe, G.; Wang, J.J. Transcriptome analysis of the citrus red mite, Panonychus citri, and its gene expression by exposure to insecticide/acaricide. Insect Mol. Biol. 2012, 21, 422–436. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likehood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar]
- Niu, J.Z.; Dou, W.; Ding, T.B.; Yang, L.H.; Shen, G.M.; Wang, J.J. Evaluation of suitable reference genes for quantitative RT-PCR during development and abiotic stress in Panonychus citri (McGregor) (Acari: Tetranychidae). Mol. Biol. Rep. 2012, 39, 5841–5849. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative Ct method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, R.; Ding, T.-B.; Niu, J.-Z.; Xia, W.-K.; Liao, C.-Y.; Dou, W.; Wang, J.-J. Molecular Characterization of Vitellogenin and Its Receptor Genes from Citrus Red Mite, Panonychus citri (McGregor). Int. J. Mol. Sci. 2015, 16, 4759-4773. https://doi.org/10.3390/ijms16034759
Zhong R, Ding T-B, Niu J-Z, Xia W-K, Liao C-Y, Dou W, Wang J-J. Molecular Characterization of Vitellogenin and Its Receptor Genes from Citrus Red Mite, Panonychus citri (McGregor). International Journal of Molecular Sciences. 2015; 16(3):4759-4773. https://doi.org/10.3390/ijms16034759
Chicago/Turabian StyleZhong, Rui, Tian-Bo Ding, Jin-Zhi Niu, Wen-Kai Xia, Chong-Yu Liao, Wei Dou, and Jin-Jun Wang. 2015. "Molecular Characterization of Vitellogenin and Its Receptor Genes from Citrus Red Mite, Panonychus citri (McGregor)" International Journal of Molecular Sciences 16, no. 3: 4759-4773. https://doi.org/10.3390/ijms16034759