
Efficiency of ITS Sequences for DNA Barcoding in Passiflora (Passifloraceae)

Abstract
:1. Introduction
2. Results
2.1. Sequence Characteristics

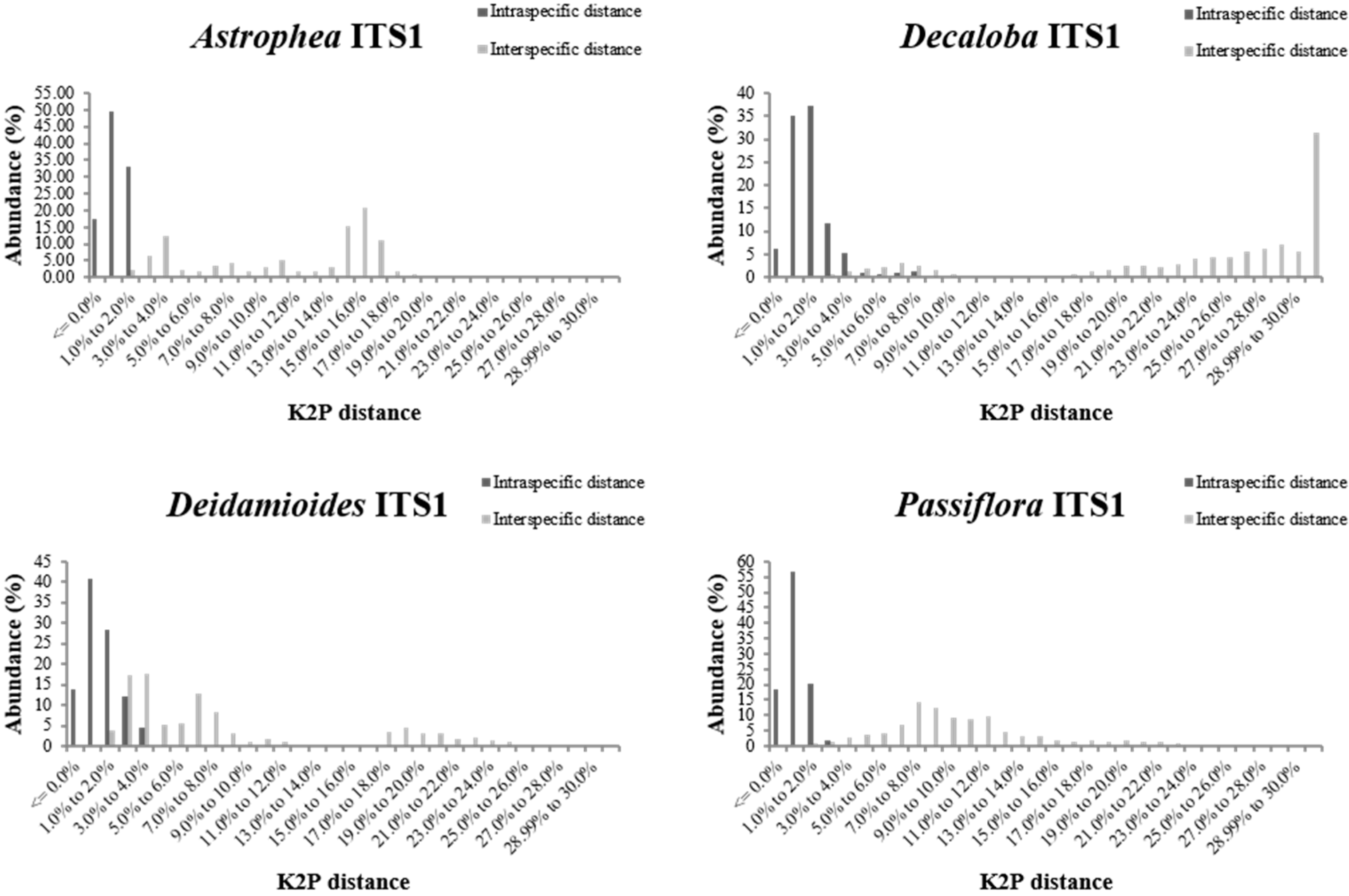{kind=link}
| Subgenus | Barcode Region | N Individuals | N Species | N Singletons | Alignment Length (bp) | Variable Characters(%) | PI Characters (%) | Overall K2P (%) |
|---|---|---|---|---|---|---|---|---|
| Astrophea | ITS1 | 53 | 16 | 12 | 291 | 32.99 | 22.68 | 8.8 |
| ITS2 | 53 | 16 | 12 | 237 | 28.27 | 16.46 | 7.2 | |
| ITS1+2 | 53 | 16 | 12 | 528 | 30.87 | 19.89 | 8.0 | |
| Decaloba | ITS1 | 314 | 134 | 85 | 359 | 76.88 | 65.46 | 24.7 |
| ITS2 | 314 | 134 | 85 | 258 | 72.87 | 56.59 | 14.0 | |
| ITS1+2 | 314 | 134 | 85 | 617 | 75.20 | 61.75 | 19.7 | |
| Deidamioides | ITS1 | 101 | 8 | 3 | 301 | 40.53 | 24.92 | 4.8 |
| ITS2 | 101 | 8 | 3 | 226 | 44.25 | 25.22 | 5.8 | |
| ITS1+2 | 101 | 8 | 3 | 527 | 42.13 | 25.05 | 5.3 | |
| Passiflora | ITS1 | 287 | 64 | 46 | 292 | 55.48 | 39.73 | 8.8 |
| ITS2 | 287 | 64 | 46 | 249 | 52.21 | 30.92 | 3.8 | |
| ITS1+2 | 287 | 64 | 46 | 541 | 53.97 | 35.67 | 6.5 |
2.2. Assessment of Barcoding Gap

2.3. “Best Match” and “Best Close Match” Analyses
| Subgenus | Barcode Region | N Individuals | BM (%) | BCM (%) | Threshold (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Correct | Ambiguous | Incorrect | Correct | Ambiguous | Incorrect | No Match | ||||
| Astrophea | ITS1 | 53 | 73.58 | 5.66 | 20.75 | 73.58 | 3.77 | 5.66 | 16.98 | 1.51 |
| ITS2 | 53 | 73.58 | 3.77 | 22.64 | 73.58 | 3.77 | 9.43 | 13.20 | 2.97 | |
| ITS1+2 | 53 | 73.58 | 3.77 | 22.64 | 73.58 | 3.77 | 5.66 | 16.98 | 1.92 | |
| Decaloba | ITS1 | 314 | 63.05 | 4.13 | 32.80 | 61.78 | 3.82 | 23.24 | 11.14 | 3.77 |
| ITS2 | 314 | 50.95 | 16.87 | 32.16 | 50.31 | 16.55 | 25.15 | 7.96 | 3.46 | |
| ITS1+2 | 314 | 64.64 | 1.27 | 34.07 | 64.01 | 1.27 | 23.88 | 10.82 | 3.43 | |
| Deidamioides | ITS1 | 101 | 96.03 | 0 | 4.96 | 95.04 | 0 | 0 | 4.95 | 2.98 |
| ITS2 | 101 | 96.03 | 0 | 4.96 | 95.04 | 0 | 0 | 4.95 | 5.41 | |
| ITS1+2 | 101 | 96.03 | 0 | 4.96 | 95.04 | 0 | 0 | 4.95 | 3.56 | |
| Passiflora | ITS1 | 287 | 81.53 | 1.74 | 16.72 | 78.39 | 1.39 | 5.57 | 14.63 | 1.83 |
| ITS2 | 287 | 54.70 | 28.57 | 16.72 | 50.87 | 27.87 | 9.40 | 11.84 | 1.19 | |
| ITS1+2 | 287 | 81.18 | 1.74 | 17.07 | 77.00 | 1.39 | 4.52 | 17.07 | 1.28 | |
2.4. Tree-Based Methods
| Subgenus | Barcode Region | N Individuals | Correct Identifications (%) | ||||
|---|---|---|---|---|---|---|---|
| NJ | ML | BI | BM | BCM | |||
| Astrophea | ITS1 | 53 | 45.28 | 22.64 | 88.68 | 73.58 | 73.58 |
| ITS2 | 53 | 35.85 | 13.21 | 50.94 | 73.58 | 73.58 | |
| ITS1+2 | 53 | 49.06 | 49.06 | 56.60 | 73.58 | 73.58 | |
| Decaloba | ITS1 | 314 | 23.25 | 22.58 | 33.76 | 63.05 | 61.78 |
| ITS2 | 314 | 21.34 | 15.29 | 29.30 | 50.95 | 50.31 | |
| ITS1+2 | 314 | 31.85 | 33.76 | 38.22 | 64.64 | 64.01 | |
| Deidamioides | ITS1 | 101 | 11.88 | 11.88 | 97.03 | 96.03 | 95.04 |
| ITS2 | 101 | 29.70 | 2.97 | 28.71 | 96.03 | 95.04 | |
| ITS1+2 | 101 | 28.71 | 27.72 | 98.02 | 96.03 | 95.04 | |
| Passiflora | ITS1 | 287 | 28.92 | 7.67 | 65.85 | 81.53 | 78.39 |
| ITS2 | 287 | 14.29 | 2.44 | 24.04 | 54.70 | 50.87 | |
| ITS1+2 | 287 | 33.80 | 33.10 | 68.64 | 81.18 | 77.00 | |
2.5. Statistical Analysis
3. Discussion
4. Experimental Section
4.1. Taxon Sampling
4.2. Data Analysis
4.2.1. Genetic Distance-Based Method
4.2.2. DNA Sequence Similarity-Based Method
4.2.3. Tree-Based Method
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identification through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, V.; Cowan, R.S.; Vogler, A.P.; Roderick, G.K.; Lane, R. Towards writing the encyclopaedia of life: An introduction to DNA barcoding. Philos. Trans. R. Soc. Lond. B 2005, 360, 1805–1811. [Google Scholar] [CrossRef]
- Hollingsworth, M.L.; Andra Clark, A.; Forrest, L.L.; Richardson, J.; Pennington, R.T.; Long, D.G.; Cowan, R.; Chase, M.W.; Gaudeul, M.; Hollingsworth, P.M. Selecting barcoding loci for plants: Evaluation of seven candidate loci with species-level sampling in three divergent groups of land plants. Mol. Ecol. Resour. 2009, 9, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haidar, N.; Savolainen, V. Land plants and DNA barcodes: Short-term and long-term goals. Philos. Trans. R. Soc. Lond. B 2005, 360, 1889–1895. [Google Scholar] [CrossRef]
- Fazekas, A.J.; Kesanakurti, P.R.; Burgess, K.S.; Percy, D.M.; Graham, S.W.; Barrett, S.C.; Newmaster, S.G.; Hajibabaei, M.; Husband, B.C. Are plant inherently harder to discriminate than animal species using DNA barcoding markers? Mol. Ecol. Resour. 2009, 9, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar]
- Kress, W.J.; Erickson, D.L. A two-locus global DNA barcode for land plants: The coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS ONE 2007, 6, e508. [Google Scholar] [CrossRef]
- Kim, H.M.; Oh, S.; Bhandari, G.S.; Kim, C.; Park, C. DNA barcoding of Orchidaceae in Korea. Mol. Ecol. Resour. 2014, 14, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Szczecinska, M.; Sawicki, J. Evaluation of 11 single-locus and seven multilocus DNA barcodes in Lamium. L. (Lamiaceae). Mol. Ecol. Resour. 2014, 14, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Vinitha, M.R.; Kumar, U.S.; Aishwarya, K.; Sabu, M.; Thomas, G. Prospects for discriminating Zingiberaceae species in India using DNA barcodes. J. Integr. Plant Biol. 2014, 56, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.G.; Hu, H.; Wang, W.; Jin, X.H. DNA barcoding of the recently evolved genus Holcoglossum (Orchidaceae: Aeridinae): A test of DNA barcode candidates. Mol. Ecol. Resour. 2011, 11, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, M.; Asif, M.; Anjum, Z.I.; Zafar, Y. Evaluating the capacity of plant DNA barcodes to discriminate species of cotton (Gossypium: Malvaceae). Mol. Ecol. Resour. 2013, 13, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Percy, D.M.; Argus, G.W.; Cronk, Q.C.; Fazekas, A.J.; Kesanakurti, P.R.; Burgess, K.S.; Husband, B.C.; Newmaster, S.G.; Barrett, S.C.; Graham, S.W. Understanding the spectacular failure of DNA barcoding in willows (Salix): Does this result from a trans-specific selective sweep? Mol. Ecol. Resour. 2014, 19, 4737–4756. [Google Scholar] [CrossRef]
- Fazekas, A.J.; Burgess, K.S.; Kesanakurti, P.R.; Graham, S.W.; Newmaster, S.G.; Husband, B.C.; Percy, D.M.; Hajibabaei, M.; Barret, S.C. Multiple multilocus DNA barcodes from the plastid genome discriminate plant species equally well. PLoS ONE 2008, 3, e2802. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.W.; Cowan, R.S.; Hollingsworth, P.M.; van den Berg, C.; Madriñan, S.; Petersen, G.; Seberg, O.; Jørgsensen, T.; Cameron, K.M.; Carine, M.; et al. A proposal for a standardized protocol to barcode all land plants. Taxon 2007, 56, 295–299. [Google Scholar]
- Sass, C.; Little, D.P.; Stevenson, D.W.; Specht, C.D. DNA barcoding in the Cycadales: Testing the potencial of proposed barcoding markers for species identification of Cycads. PLoS ONE 2007, 2, e1154. [Google Scholar] [CrossRef] [PubMed]
- China Plant BOL Group. Comparative analysis of a large dataset indicates that internal transcribed spacer (ITS) should be incorporates into the core barcode for seed plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19641–19646. [Google Scholar]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenet. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M. Refining the DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19451–19452. [Google Scholar] [CrossRef] [PubMed]
- Muellner, A.N.; Schaefer, H.; Lahaye, R. Evaluation of candidate DNA barcoding loci for economically important timber species of the mahogany family (Meliaceae). Mol. Ecol. Resour. 2011, 11, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Q.; Chen, J.Y.; Wang, S.; Xiong, S.Z. Evaluation of six candidate DNA barcoding loci in Ficus (Moraceae) of China. Mol. Ecol. Resour. 2012, 12, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.B.; Wang, Y.P.; Möller, M.; Gao, L.M.; Wu, D. Applying plant DNA barcodes to identify species of Parnassia (Parnacciaceae). Mol. Ecol. Resour. 2012, 12, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; Wang, F.Y.; Yan, H.F.; Hao, G.; Hu, C.M.; Ge, X.J. Testing DNA barcoding in closely related groups of Lysimachia. L. (Myrsinaceae). Mol. Ecol. Resour. 2012, 12, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Duan, L.; Zhou, N. Application of DNA barcoding in Roscoea (Zingiberaceae) and a primary discussion on taxonomic status of Roscoea cautleoides var. Pubescens. Biochem. Syst. Ecol. 2014, 52, 14–19. [Google Scholar] [CrossRef]
- Alves, T.L.S.; Chauveau, O.; Eggers, L.; Souza-Chies, T.T. Species discriminatory in Sisyrinchium (Iridaceae): Assessment of DNA barcodes in a taxonomically challenging genus. Mol. Ecol. Resour. 2013, 14, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef]
- Gao, T.; Yao, H.; Song, J.; Zhu, Y.; Liu, C.; Chen, S. Evaluating the feasibility of using candidate DNA barcodes in discriminating species of the large Asteraceae family. BMC Evol. Biol. 2010, 10, 324. [Google Scholar] [PubMed]
- Gao, T.; Yao, H.; Song, J.; Liu, C.; Zhu, Y.; Ma, X.; Pang, X.; Xu, H.; Chen, S. Identification of medicinal plants in the family Fabaceae using a potential DNA barcode ITS2. J. Ethnopharmacol. 2010, 130, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhu, Y.; Chen, X.; Liao, B.; Yao, H.; Song, J.; Chen, S.; Meng, F. The short ITS2 sequence serves as an efficient taxonomic sequence tag in comparison with the full-length ITS. Biomed. Res. Int. 2013, 2013, 741476. [Google Scholar] [PubMed]
- Wang, M.; Zhao, HX.; Wang, L.; Wang, T.; Yang, R.W.; Wang, X.L.; Zhou, Y.H.; Ding, C.B.; Zhan, G.L. Potential use of DNA barcoding for the identification of Salvia based on cpDNA and nrDNA sequences. Gene 2013, 528, 206–215. [Google Scholar] [CrossRef]
- Wang, X.C.; Liu, C.; Huang, L.; Bengtsson-Palme, J.; Chen, H.; Zhang, J.H.; Cai, D.; Li, J.Q. ITS1: A DNA barcode better than ITS2 in eukaryotes? Mol. Ecol. Resour. 2014. [Google Scholar] [CrossRef]
- Ulmer, T.; MacDougal, J.M. Passiflora: Passionflowers of the World, 1st ed.; Timber Press: Portland, OR, USA, 2004; pp. 7–45. [Google Scholar]
- Krosnick, S.E.; Porter-Utley, K.E.; MacDougal, J.M.; Jørgensen, P.M.; McDade, L.A. New insights into the evolution of Passiflora subgenus Decaloba. (Passifloraceae): Phylogenetic relationships and morphological synapomorphies. Syst. Bot. 2013, 38, 692–713. [Google Scholar] [CrossRef]
- Deginani, N.B. Las especies argentinas del género Passiflora (Passifloraceae). Darwiniana 2001, 39, 43–129. [Google Scholar]
- Krosnick, S.E.; Freudenstein, J.V. Monophyly and floral character homology of old world Passiflora (Subgenus Decaloba: Supersection Disemma). Syst. Bot. 2005, 30, 139–152. [Google Scholar] [CrossRef]
- Killip, E.P. The American species of Passifloraceae. Field Mus. Nat. Hist. Bot. Ser. 1938, 19, 1–613. [Google Scholar]
- Escobar, L.K. A new subgenus and five new species in Passiflora. (Passifloraceae) from South America. Ann. Mo. Bot. Gard. 2009, 76, 877–885. [Google Scholar] [CrossRef]
- Feuillet, C.; MacDougal, J.M. A new infrageneric classification of Passiflora. L. (Passifloraceae). Passiflora 2003, 13, 34–38. [Google Scholar]
- Yockteng, R.; Nadot, S. Phylogenetic relationships among Passiflora species based on the glutamine synthetase nuclear gene expressed in chloroplast (ncpGS). Mol. Phylogenet. Evol. 2004, 31, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Muschner, V.C.; Lorenz, A.P.; Cervi, A.C.; Bonatto, S.L.; Souza-Chies, T.T.; Salzano, F.M.; Freitas, L.B. A first molecular phylogenetic analysis of Passiflora (Passifloraceae). Am. J. Bot. 2003, 90, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Muschner, V.C.; Zamberlan, P.M.; Bonatto, S.L.; Freitas, L.B. Phylogeny, biogeography and divergence times in Passiflora (Passifloraceae). Genet. Mol. Biol. 2012, 35, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.K.; Gilbert, L.E.; Simpson, B.B.; Downie, S.R.; Cervi, A.C.; Jansen, R.K. Phylogenetic relationships and chromosome number evolution in Passiflora. Syst. Bot. 2006, 31, 138–150. [Google Scholar] [CrossRef]
- Hansen, A.K.; Escobar, L.K.; Gilbert, L.E.; Jansen, R.K. Paternal, maternal, and biparental inheritance of the chloroplast genome in Passiflora (Passifloraceae): Implications for phylogenetic studies. Am. J. Bot. 2007, 94, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira-Silva, C.B.M.; Jesus, O.N.; Santos, E.S.L.; Corrêa, R.X.; Souza, A.P. Genetic breeding and diversity of the genus Passiflora: Progress and perspectives in molecular and genetic studies. Int. J. Mol. Sci. 2014, 15, 14122–14152. [Google Scholar] [CrossRef] [PubMed]
- Lorenz-Lemke, A.P.; Muschner, V.C.; Bonatto, S.L.; Cervi, A.C.; Salzano, F.M.; Freitas, L.B. Phylogeographic inferences concerning evolution of Brazilian Passiflora actinia and P. elegans (Passifloraceae) based on ITS (nrDNA) variation. Ann. Bot. 2005, 95, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Koehler-Santos, P.; Lorenz-Lemke, A.P.; Muschner, V.C.; Bonatto, S.L.; Salzano, F.M.; Freitas, L.B. Molecular genetic variation in Passiflora alata (Passifloraceae), an invasive species in southern Brazil. Biol. J. Linn. Soc. Lond. 2006, 88, 611–630. [Google Scholar] [CrossRef]
- Mäder, G.; Zamberlan, P.M.; Fagundes, N.J.R.; Magnus, T.; Salzano, F.M.; Bonatto, S.L.; Freitas, L.B. The use and limits of ITS data in the analysis of intraspecific variation in Passiflora. L. (Passifloraceae). Genet. Mol. Biol. 2010, 33, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Cazé, A.L.R.; Mäder, G.; Bonatto, S.L.; Freitas, L.B. A molecular systematic analysis of Passiflora ovalis and Passiflora contracta (Passifloraceae). Phytotaxa 2013, 132, 39–46. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Gielly, L.; Miguel, C.; Valentini, A.; Vermat, T.; Corthier, G.; Brochmann, C.; Willerslev, E. Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. Nucleic Acids Res. 2007, 35, e14. [Google Scholar] [CrossRef] [PubMed]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Möller, M.; Gao, L.M.; Zhang, D.Q.; Li, D.Z. DNA barcoding for the discrimination of Eurasian yews (Taxus. L., Taxaceae) and the discovery of cryptic species. Mol. Ecol. Resour. 2011, 11, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Aubriot, X.; Lowry, P.P., II; Cruaud, C.; Couloux, A.; Haevermans, T. DNA barcoding in a biodiversity hot spot: Potential value for the identification of Malagasy Euphorbia L. listed in CITES Appendices I and II. Mol. Ecol. Resour. 2013, 13, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.C.; Zhang, J.; Han, J.P.; Song, J.Y.; Yao, H.; Zhu, Y.J.; Li, J.C.; Wang, Z.Z.; Xiao, W.; Lin, Y.L.; et al. Testing the potential of proposed DNA barcodes for species identification of Zingiberaceae. J. Syst. Evol. 2011, 49, 261–266. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTALX windows interface: flexible strategies for multiple sequences alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Theodoridis, S.; Stefanaki, A.; Tezcan, M.; Aki, C.; Kokkini, S.; Vlachonasios, K.E. DNA barcoding in native plants of the Labiatae (Lamiaceae) family from Chios Island (Greece) and the adjacent Çesme-Karaburun Peninsula (Turkey). Mol. Ecol. Resour. 2012, 12, 620–633. [Google Scholar] [CrossRef] [PubMed]
- SPSS Inc. Released; PASW Statistics for Windows, Version 18.0; SPSS Inc.: Chicago, IL, USA, 2009. [Google Scholar]
- Dong, W.; Cheng, T.; Li, C.; Xu, C.; Long, P.; Chen, C.; Zhou, S. Discriminating plants using the DNA barcode rbcLb: An appraisal based on a large dataset. Mol. Ecol. Resour. 2013, 14, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Song, J.; Zhu, Y.; Xie, C.; Chen, S. Using DNA barcoding to identify species within Euphorbiaceae. Planta Med. 2010, 76, 1784–1786. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giudicelli, G.C.; Mäder, G.; Brandão de Freitas, L. Efficiency of ITS Sequences for DNA Barcoding in Passiflora (Passifloraceae). Int. J. Mol. Sci. 2015, 16, 7289-7303. https://doi.org/10.3390/ijms16047289
Giudicelli GC, Mäder G, Brandão de Freitas L. Efficiency of ITS Sequences for DNA Barcoding in Passiflora (Passifloraceae). International Journal of Molecular Sciences. 2015; 16(4):7289-7303. https://doi.org/10.3390/ijms16047289
Chicago/Turabian StyleGiudicelli, Giovanna Câmara, Geraldo Mäder, and Loreta Brandão de Freitas. 2015. "Efficiency of ITS Sequences for DNA Barcoding in Passiflora (Passifloraceae)" International Journal of Molecular Sciences 16, no. 4: 7289-7303. https://doi.org/10.3390/ijms16047289
APA StyleGiudicelli, G. C., Mäder, G., & Brandão de Freitas, L. (2015). Efficiency of ITS Sequences for DNA Barcoding in Passiflora (Passifloraceae). International Journal of Molecular Sciences, 16(4), 7289-7303. https://doi.org/10.3390/ijms16047289